查干湖大型底棲動物群落結構及與環境因子的關系
2021-01-15 02:07:08都雪楊敬爽宋聃王慧博金星劉輝王樂霍堂斌
水產學雜志 2020年6期
都雪 ,楊敬爽,宋聃,王慧博,金星,劉輝,王樂,霍堂斌
(1.中國水產科學研究院黑龍江水產研究所,黑龍江 哈爾濱 150070;2.吉林查干湖國家級自然保護區管理局,吉林 松原 138000)
大型底棲動物是魚類等捕食者的重要食物資源,在耦合湖泊底層營養與水層營養、水體生物分解(降低有機污染)和加速物質循環等多方面維持湖泊生態系統結構和功能[1,2],在湖泊食物網中發揮重要作用。大型底棲動物生命周期長、遷移能力有限,物種類分布和生長、繁殖、群落演替和群落結構的變化受水質、水深、水溫和底質等環境因子影響[3,4]。大型底棲動物分布廣泛、形體較易辨認、對污染的逃避能力弱和對環境變化較為敏感等優勢,其群落結構是認識水域環境特點、預測環境質量的重要指標,已廣泛地應用于湖泊環境監測[5,6]。
大型底棲動物群落的分類學組成受到廣泛關注,而關于其功能組成的知識則相對有限。物種主要通過在生態系統中的特定功能來影響環境,因此,準確描述群落內功能性狀的結構和組成,可以更好地理解多樣性與功能之間的關系[7]。大型底棲動物的生態功能通常與一些特定的功能攝食類群(functional feeding groups,反映每個物種的主要攝食類型)有關。大型底棲動物的功能攝食類群通常分為撕食者(shredders)、刮食者(scrapers)、收集者(gatherers)、濾食者(filters)和捕食者(predators)[8]。當生態系統受到環境干擾時,功能性狀也會作為指示指標發生改變[9,10]。
查干湖是吉林省最大的內陸湖泊,地處嫩江與霍林河交匯的水網地區,是吉林省重要的漁業基地,也是松遼平原第一大湖。查干湖魚類種類豐富,魚類動物區系相對復雜,魚類種質資源優良,建有吉林前郭查干湖蒙古鲌國家級水產種質資源保護區和吉林前郭新廟泡特有魚類國家級水產種質資源保護區。目前,關于查干湖大型底棲動物的研究僅限于種類組成及其與水環境的關系[11],關于其功能攝食類群與環境因子間的關系研究較少。因此,開展查干湖大型底棲動物群落特征調查,探討其與主要環境生態因子的相互關系,比較基于物種組成和基于功能攝食類群組成的大型底棲動物群落特征對環境因子的響應強度,對于了解湖泊大型底棲動物的結構和功能,合理利用湖泊水資源和生物資源具有重要意義。
1 材料與方法
1.1 研究區域
查干湖(124°03'~124°34',45°09'~45°30')位于我國吉林省西北部、內蒙古自治區、黑龍江省和吉林省的金三角地區,屬霍林河末端的堰塞湖泊;是松花江、松花江南源、嫩江三江交匯處我國十大淡水湖之一、吉林省內最大的天然湖泊和最大的漁業生產基地。水域面積420 km2,平均水深2.5 m,水位在130 m 時,與辛甸泡、馬營泡構成一體,其姊妹泡是新廟泡和庫里泡,目前由引松渠將其貫通。查干湖補水主要來自天然降水、松花江引水及灌區回歸水。周邊地區為農業種植區,農業面源污染嚴重;湖周鹽堿地,沙地分布面積大,是湖區徑流匯集區;周邊旅游業及餐飲服務業快速發展,威脅查干湖水質。根據查干湖的生境特點,共設立9 個采樣點(圖1)。
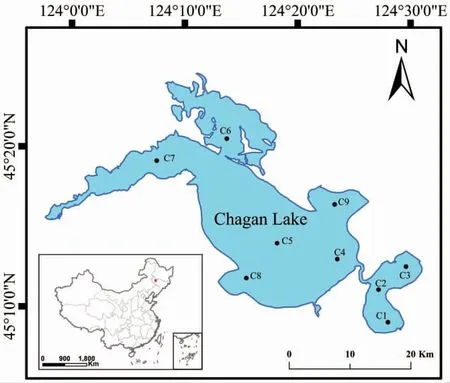
圖1 查干湖采樣點分布Fig.1 Distribution of sampling sites in Chagan Lake
1.2 大型底棲動物樣品采集與處理
2017 年春季(4 月)、夏季(7 月)和秋季(10 月)分3 個季節調查了查干湖的水質、大型底棲動物狀況。用1/16 m2的改良彼德森采泥器采集大型底棲動物定量樣品,每個調查斷面采集3 次,底泥采用40 目和60 目分樣篩進行篩選。所采集的大型底棲動物用4%甲醛溶液固定后帶回實驗室,再移入75%酒精中長期保存。在室內進行種類鑒定、個體計數。
根據攝食策略,將大型底棲動物分為5 個功能攝食類群:(1)捕食者;(2)刮食者;(3)收集者;(4)濾食者和(5)撕碎者[8]。將每個功能攝食類群中的物種密度求和,即為該功能攝食類群的總密度。
1.3 環境因子
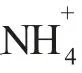
1.4 數據分析
運用主成分分析(Principal Component Analysis,簡稱PCA)簡化大型底棲動物的物種和功能攝食類群,在簡化數據集的同時評價不同季節湖泊大型底棲動物物種和功能攝食類群種類組成的特征。主成分分析前,將數據Hellinger 轉化,以減少其變異性[13]。主成分分析采用R 軟件[14]的“ade4”數據包[15]。
用冗余分析方法(Redundancy analysis,簡稱RDA)分析大型底棲動物的物種和功能攝食類群種類組成與環境因子之間的關系。數據分析前,環境變量進行[log(x)]轉換,物種數據進行Hellinger 轉化。環境因子的篩選采用向前引入法(Forward selection),環境變量的顯著性用Monte Carlo 方法檢驗(999 次無限制排列)。RDA 采用R 軟件的“vegan”數據包進行分析[16]。
2 結果與分析
2.1 調查期間的水質特征

2.2 大型底棲動物的種類組成及季節差異
調查期間共發現大型底棲動物35 種,其中水生昆蟲和軟體動物各14 種,環節動物6 種,甲殼動物1 種(表2)。優勢類群為搖蚊類和寡毛類等小型耐污種,如秋月齒斑搖蚊Stictochironomus akizukii、項圈無突搖蚊Ablabesmyia monilis、中國長足搖蚊Tanypus chinensis、半球多脈扁螺Segmentina hemisphaerula、蘇式尾鰓蚓Branchiura sowerbyi、正顫蚓Tubifex tubifex、奧特開水絲蚓Limnodrilus udekemianus 和低頭石蠶Neureclipsis sp.。
PCA 結果顯示,大型底棲動物的種類組成存在季節性差異(圖2-a)。相似性分析(ANOSIM)結果同樣顯示,不同季節大型底棲動物的種類組成存在顯著性差異(R=0.491,P=0.001),夏季種群的差異性顯著高于春季和秋季(圖2-b)。大型底棲動物功能攝食類群的季節性差異較小(R=0.087,P=0.07),且夏、秋兩季大型底棲動物功能攝食類群組成的差異大于季節間差異(圖2-d)。春季大型底棲動物的密度最高,秋季最低(圖3),三個季節中大型底棲動物的收集者所占比例均最高,刮食者的比例也從春季到秋季逐漸降低。
2.3 環境因子對大型底棲動物群落結構的影響
根據查干湖大型底棲動物物種與環境的關系,RDA 分析最終篩選出WT、WD、Chla、TP 和DO 等5個與大型底棲動物群落變化最顯著相關的環境因子。第一排序軸和第二排序軸的特征根分別為0.185 和0.063,分別解釋了26.14%和8.89%的物種數據方差變異及44.61%和15.17%的物種-環境關系變異。第一排序軸與WT 和TP 的相關性較高,第二排序軸與WD、Chla 和DO 的相關性較高(圖4-a)。
根據查干湖大型底棲動物功能攝食類群與環境的關系,RDA 分析最終篩選出DO、WD 和TSS 等3 個與大型底棲動物群落變化最顯著相關的環境因子。第一排序軸和第二排序軸的特征根分別為0.059 和0.031,分別解釋了21.61%和11.48%的物種數據方差變異及54.71%和29.05%的物種-環境關系變異。第一排序軸與DO 呈顯著正相關、與TSS呈顯著負相關,第二排序軸與WD 呈顯著正相關(圖4-b)。
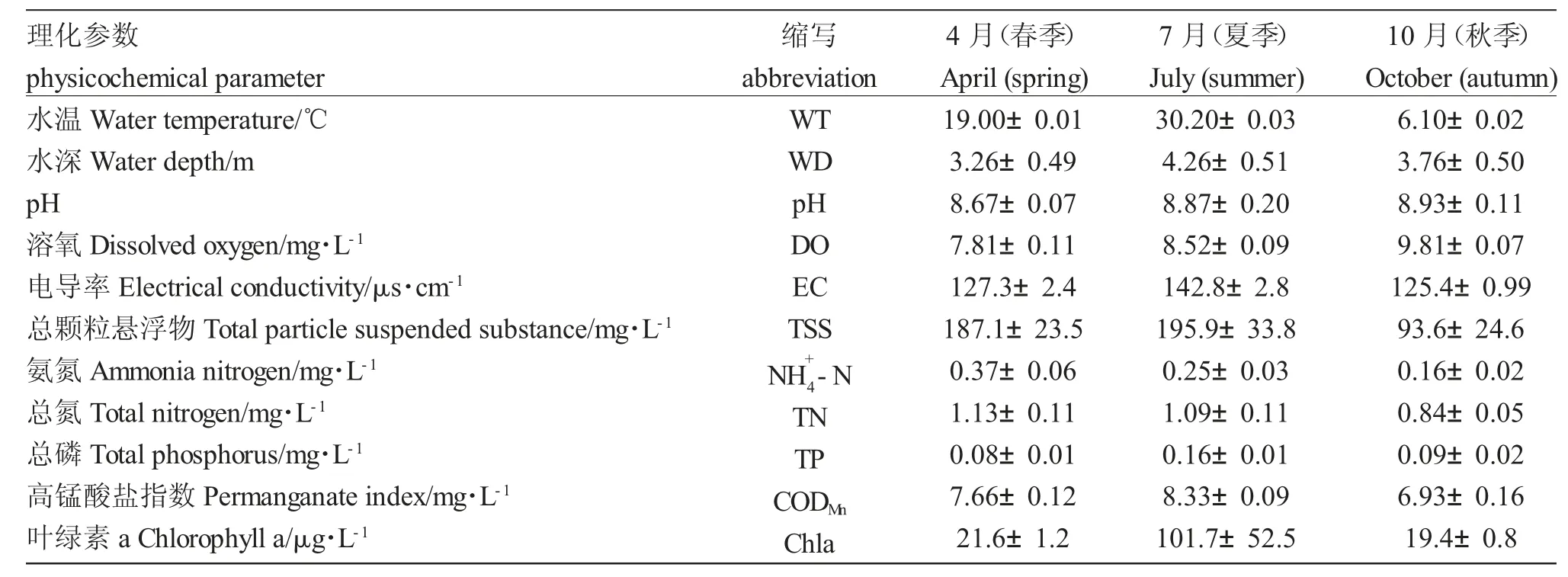
表1 查干湖理化因子(平均值±標準誤)的季節變化Tab.1 Seasonal variations in physicochemical parameters(mean ± SE)measured in Chagan Lake

表2 查干湖大型底棲動物種類組成Tab.2 Species composition of macroinvertebrate community in Chagan Lake
3 討論
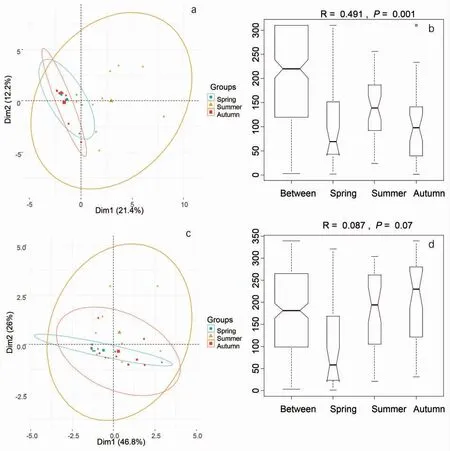
圖2 不同季節大型底棲動物群落物種(a)和攝食功能群(c)組成的主成分分析及不同季節大型底棲動物物種(b)和攝食功能群(d)的相似性分析(ANOSIM)Fig.2 The principal component analysis(PCA)of Hellinger-transformed macroinvertebrates species composition(a)and functional feeding groups(c)among seasons.Single seasons represented by circles.Analysis of similarities(ANOSIM)boxplots displaying the dissimilarities in macroinvertebrates species(b)and functional feeding group compositions between and within single season
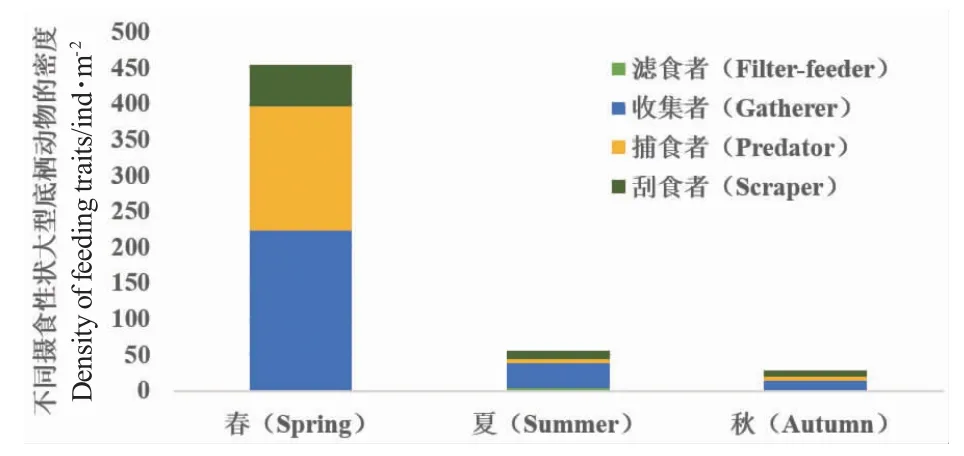
圖3 不同季節(春、夏和秋季)各攝食性狀大型底棲動物的平均密度Fig.3 Mean density of macroinvertebrates with different feeding traits in three different seasons:spring,summer,and autumn
2017 年調查期間,采集查干湖大型底棲動物35 種,其中水生昆蟲和軟體動物種類最多,均為14種,分別占總數40%。大型底棲動物種類組成、群落結構受水域的營養水平、底質類型及水生植物狀況等因素影響[3-5]。受到不同來源補給水(特別是前郭灌區的回灌水)水質的影響,以及松嫩平原區域及湖區生態系統演化的多種動態因素綜合作用[17-19],查干湖的水質有逐年惡化的趨勢,其中溶解氧濃度降低,pH 升高,氮、磷等污染物濃度有所增加。近年來,綜合評價表明,查干湖水質接近Ⅲ類標準,處于輕度富營養化狀態[20,21]。
大型底棲動物對環境敏感,其群落組成可以指示湖泊水質的變化[5,6]。大型底棲動物的攝食策略可以反映物種對環境壓力的適應[22],不同的功能攝食類群可以反映生境特征、營養循環、水質等信息[8,10]。本研究中,搖蚊類和寡毛類等小型耐污種為底棲動物群落的優勢類群。本研究結果表明,水溫、水深、葉綠素a、總磷和溶解氧等是影響大型底棲動物種類組成的主要環境因子(圖4-a),水深、溶解氧和總顆粒懸浮物是影響大型底棲動物功能攝食類群的主要因素(圖4-b)。Kitagawa 認為,湖體底部溶解氧水平是決定底棲動物分布的首要因素[23]。本研究中,大型底棲動物種類組成受溶解氧含量的影響,支持了Kitagawa 的觀點。較高的總磷含量可以增加湖泊的初級生產力,促進植食性小型底棲動物的生長[24];然而,較高的初級生產力還可能由于較高的微生物呼吸速率而導致沉積物缺氧,這會對無脊椎動物產生負面影響[25]。
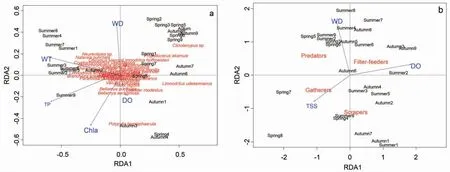
圖4 查干湖大型底棲動物物種(a)和功能攝食類群(b)與環境因子的冗余分析Fig.4 Redundancy analysis (RDA) of macroinvertebrate species composition (a) and feeding functional groups (b) with environmental variables in Chagan Lake
春季和夏季,以有機顆粒為食的收集者和以藻類為食的刮食者是大型底棲動物的優勢類群,主要是春夏季較高的水溫和光照強度促使藻類大量生長,為收集者和刮食者提供了豐富的藻類食物資源[26]。本研究中,水深與收集者和刮食者密度呈負相關。收集性狀通常與水深呈負相關[8],隨著湖泊水深的降低,風力擾動增加了有機顆粒再懸浮,間接影響了水體中有機顆粒的含量,即收集者的食物資源。春季湖泊平均水深最低,這也解釋了春季收集者密度最高,且收集者分布與水深呈負相關。較深的水域營養物質含量相對較低[10],也是收集者與水深呈負相關的因素之一。三個季節中,收集性狀均是大型底棲動物群落中最常見的攝食性狀。這一結果表明,盡管物種的分類學特征隨季節變化,但收集性狀仍然是湖泊中最優勢的性狀。本研究中的大多數昆蟲物種,以及幾乎所有的寡毛類都在一定程度上將顆粒收集作為攝食特征[27]。收集性狀的廣泛分布進一步證實了大型底棲動物在湖泊有機物循環中的作用[1]。從春季到秋季,大型底棲動物的密度逐漸下降,其原因可能是浮游植物和浮游動物的生物量逐漸降低導致沉降的有機顆粒供應減少。綜上,評估基于物種組成和功能攝食類群組成的大型底棲動物群落特征對環境因子的響應,可以更好地了解其在湖泊生態系統中的功能,為合理利用湖泊水資源和生物資源具有重要意義。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
中華詩詞(2022年6期)2022-12-31 06:41:24
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
中國生殖健康(2019年11期)2019-01-07 01:28:02
汽車觀察(2018年10期)2018-11-06 07:05:26
中國科技論壇(2017年7期)2017-07-25 08:49:53
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
