轉化生長因子-β(TGF-β)基因家族在水產養殖中的潛在應用價值
2021-01-15 02:07:10魯翠云匡友誼鄭先虎孫志鵬孫效文
水產學雜志 2020年6期
關鍵詞:生長
魯翠云,匡友誼,鄭先虎,孫志鵬,孫效文
(中國水產科學研究院黑龍江水產研究所,黑龍江 哈爾濱 150070)
轉化生長因子-β(transforming growth factorβ,TGF-β)基因是1978 年在小鼠(Mus musculus)肉瘤病毒轉化的細胞株條件培養基中被鑒定出的,后來逐步發現這是一個龐大的基因家族。該家族已發現多個基因,包括TGF-β、骨形態發生蛋白(bone morphogenetic protein,BMP)、生長分化因子(growth differentiation factor,GDF)、活化素(Activin)等,可分為兩大亞家族即TGF-β/Activin/Nodal(胚胎形態發生素)和BMP/GDF/MIS(穆勒氏抑制物)。TGF-β 基因具有調節細胞生長、參與和促進細胞免疫、腫瘤發生、心肌生長、細胞增殖、骨骼生長平衡等多種功能,是開發藥物的理想模型,甚至已成立了專門開發TGF-β 信號通路及相關生物過程有關的藥物公司。除了基礎研究、醫學和藥物學等領域涉及這個基因超家族外,近年來農業養殖動物領域也開始重視這個家族的基因參與多種經濟性狀遺傳調控的機制,希望在一些重要經濟性狀的遺傳解析中獲得新的突破。根據最新研究結果分析,水產養殖業最關心的幾個重要性狀如生長、飼料轉化率、疾病與免疫、性別分化與調控、肌間刺等都有這個家族基因參與的痕跡。本文簡要介紹了TGF-β超家族主要研究進展,分析該家族在闡明養殖魚類經濟性狀的遺傳機制方面的潛在價值。
1 TGF-β 的基本功能
TGF-β 廣泛存在于各種組織中,多種細胞表面具有TGF-β 受體,這使其具有多種功能的生物化學基礎,也是能夠影響多種經濟性狀的根本原因。
1.1 炎癥與免疫
最早發現TGF-β 具有調節炎癥發生和降低炎癥的作用。上世紀九十年代前后就已開始探索TGF-β 類藥物在消除炎癥中的作用[1]。TGF-β 屬自蛋白(self-protein),和B-細胞免疫沉默難以獲得高滴度抗體,多是用融合蛋白獲得高滴度抗體和確定其免疫功能[2]。
在多個TGF-β 成員中,TGF-β1 主要參與免疫能力的調節和炎癥的控制,調節免疫細胞的募集、保留、凋亡、分化和增殖,因此可以根據細胞情況來增強或抑制炎癥反應,TGF-β1 在感染過程中抑制了CD8+細胞毒性T 細胞的擴增,但同時也促進了促炎性CD4+Th17 細胞和調節性T 細胞的發展,抑制了T 細胞依賴性炎癥[3]。TGF-β 參與免疫與炎癥還有很多精細的調控機制,已經和繼續為人類、畜禽、魚類的疾病防治提供基礎知識。
1.2 生長與細胞增殖
參與生長控制和細胞增殖過程的調控也是TGF-β 早期被發現的主要功能之一,在2000 年之前就已發現這個作用。當時考察發現,TGF-β 超家族中的兩個亞家族都有這個功能[4]。近年更深入地認識了TGF-β 參與生長控制的細節。譬如:哺乳動物的腸上皮細胞不斷進行翻轉,隱窩中的細胞增殖而絨毛中的細胞凋亡;TGF-β 和胃泌素釋放肽(gastrin-releasing peptide,GRP)均參與腸上皮細胞的分裂、分化、粘附、遷移和死亡調節[5]。
TGF-β 超家族中對生長調節功能較大且應用較多的是GDF 亞家族中的肌肉生長抑制素(Myostatin,MSTN)及同一信號通路的其他基因。Lee 研究團隊在1997 年鑒定到GDF8(Myostatin)基因對小鼠和比利時藍牛Bos taurus 肌肉的負調控作用[6,7],后續多個研究證實,肌肉生長抑素具有進化保守性,多種動物敲除實驗證實其具有同樣的調節肌肉生長的功能,從而開啟了利用該基因創制肌肉明顯增大的動物新品種的潮流[8]。
MSTN 基因有多種機制調節肌肉生長,一種是與絲氨酸/蘇氨酸激酶受體(ActRIIB)結合直接作用于肌細胞,通過減少肌細胞數量和肌纖維數量來控制肌肉的生長;另一種是通過內分泌系統調節全身的肌肉生長[9,10]。內分泌調節是MSTN 調節垂體發育,或者通過調節肝臟產生的胰島素樣生長因子1(insulin-like growth factor 1,IGF1)來影響肌肉生長,對肌肉生長起負調控作用[11,12]。
1.3 骨細胞平衡
BMP 基因家族中除BMP1 外都屬于TGF-β 超家族成員,特征之一是兩個亞家族成員的一級序列在核心多肽上有40%以上的同源序列;之二是它們與TGF-β 家族成員在結合受體后基本都是通過SMAD 蛋白達到信號向下傳導。
BMP 功能與骨發生、再生或軟骨發生有關,是研究骨骼相關疾病的主要靶標通路。這個家族有十幾個蛋白,都有促進骨骼發生,也都在胚胎發生過程中起許多關鍵作用[13]。但每個BMP 起的作用在不同發育時期和不同組織各有特點。如BMP6 可以挽救BMP7 null 突變體中的眼缺陷,而BMP4 不能[14]。BMP 調節骨生長的同時對動物的酮體生長也有明顯的調控作用。
1.4 性別決定與性別分化
目前已鑒定多個TGF-β 基因家族中的基因參與魚類性別的決定和分化,與這個基因超家族的基本功能一致,即可以調控囊胚期細胞的增殖和減少。TGF-β 信號減少了生殖細胞的數量,促進性腺向精巢發育并決定了性別和性腺的分化[15,16];另一途徑是TGF-β 基因家族成員通過抑制芳香化酶的活性決定性別和促進性腺分化[17]。
2 TGF-β 基因家族在魚類經濟性狀遺傳機制研究中的應用與價值
水產養殖所重視的經濟性狀多是多基因調控的性狀,如生長速度、飼料轉化、脂肪代謝等,很難獲得與性狀對應的生化產物,從遺傳連鎖角度鑒定這些經濟性狀相關的遺傳位點和QTL 是一種可行的研究策略,借鑒人和模式生物相關性狀的研究結果也是一種捷徑。下面介紹的研究結果部分來自水產生物,部分來自水產生物能夠借鑒的其他生物。
2.1 水產生物TGF-β 基因家族基因的克隆與鑒定
水產生物相關基因的克隆一般根據模式生物鑒定到的新基因,推測可能具有產業應用價值的則跟進克隆。肌肉生長抑制素GDF8(Myostatin)基因可能是較早克隆出的TGF-β 家族中的基因[18]。科學家們相信這個對肌肉形成非常有調控能力的基因將掀起一次農業動物生產的革命。雖然至今這個革命仍沒有到來,但持續的研究證明,敲除這個基因能顯著改善肌肉質量和數量,對動物肉類生產的巨大推進作用將會到來。
魚類中TGF-β 超家族各亞類基因基本上與人的數量相近[19],這也符合該家族基因保守性較高的估計;軟體動物門、半索動物門各亞類的基因數量都較少,或是因為還沒有克隆或者就是進化缺失。隨著高通量測序的進行,會有更多水產生物的TGF-β 超家族各亞類基因被克隆出來。
2.2 生長
魚類中Myostatin 基因的研究報道很多。早期利用小RNA 干擾技術和蛋白拮抗技術,試圖通過降低Myostatin 的表達量達到提高受體魚的生長速度。閆學春等[20]報道,卵泡抑素(follistatin,Fst)蛋白具有拮抗TGF-β 超家族許多成員的功能,通過直接的轉目標基因可抑制肌肉生長抑制素(myostatin)的活性,提高目標鯉Cyprinus carpio 肌肉的生長。還有利用MSTN 通過Crispr/Case9 等基因編輯技術改造養殖魚類的報告。如Khalil 等[21]報道,用此基因構建的快速生長的基因敲除的溝鯰Ictalurus punctatus,但養殖魚類這方面的報道很少,尤其是國內幾乎沒有。畜禽利用肌肉生長抑素的報道特別多,主要原因可能是中國在大動物轉基因研究投入較大。雖然與養殖魚類遇到了同樣的轉基因產品難以商業化的生物安全管理問題,但有研究經費支持還是能夠促進一些研究工作持續開展,中國水產養殖業在這項高技術的研發上遇到了重大危機,主要是經費支持過少。
李勝杰等[22]利用肌內生長抑素基因上的多態標記和IGF1 等標記,篩選出快速生長的大口黑鱸Micropterus salmoides 優異種質,或可作為新品種選育的工具。
2.3 飼料轉化
飼料占水產養殖成本的65%以上(成體或亞成體養殖),提高飼料效率能夠增加養殖收益,減少水體污染。但是,飼料轉化率是一個與生長和代謝相關的綜合性狀,難以直接獲得相關的生化指標和基因,更難確定基因所在的信號通路。本課題組在鯉飼料轉化率(feed conversion ratio,FCR)的QTL 區間鑒定出18 個候選基因,其中4 個基因gdf7、smad7、tgfb2 和tgfbr2 集中于TGF-β 信號通路,而gdf7 位于兩個種質共享的QTL 區間(qFCRM12 和qF CRH24),分別解釋22.3%和33.4%的表型變異,tgfbr2 和Smad7 位于qFCRM16,可解釋表型變異率20.9%;tgfb2 位于qFCRM33,可解釋表型變異率28.4%。還有一個重要生長相關基因IGF2[23],在通路上也與TGF-β 相關聯。
有關畜禽TGF-β 參與飼料轉化已有少量報道。Jing 等[24]對豬Sus scrofa 剩余采食量(residual feed intake,RFI)高、低差異顯著的兩組樣本進行轉錄組分析,發現TGF-β 通路的基因在兩組樣品中有顯著地差異表達,認為提高TGF-β 通路的基因表達水平能夠提高豬飼料效率;Liu 等[25]在肉雞Gallus gallus RFI 高、低兩組樣本的轉錄組分析中發現,TGF-β 信號通路的基因具有顯著表達差異,可以作為飼料效率的候選基因。Jin 等[26]從TGF-β3 基因中篩選到與雞FCR 顯著相關的SNP 標記。該基因位于雞與FCR 緊密連鎖的QTL 區間附近,后續的研究從TGF-β3 及其受體基因中鑒定到nsSNP 和SNVs 標記均對雞的FCR 具有顯著貢獻[27,28]。對基因功能的分析發現,TGFBR3 基因與TGFBR2 有協同表達的特征。
2.4 脂肪沉積與代謝
TGF-β 基因超家族參與動物脂肪代謝已確定,但相關機制研究的很少。TGF-β 基因超家族參與動物脂肪代謝的途徑之一是:MSTN 抑制Pparγ和Cebpα 表達來抑制3T3-L1 脂肪細胞的分化,通過抑制關鍵的脂肪酶和促進脂解酶的表達來減少3T3-L1 細胞中脂質的積累,起到調節完全分化的3T3-L1 脂肪細胞中脂肪因子的表達和分泌的作用[29];MSTN 也可通過將白色脂肪轉化為棕色脂肪來調節脂肪的代謝[30]。BMP 亞類在成骨和破骨的骨骼平衡過程中依賴脂肪提供能量,也影響三大類脂肪組織的骨髓脂肪代謝[31]。但Pparγ 的綜述中沒有涉及肌生長抑素和BMP 對脂肪平衡的影響[32],這些轉錄因子在脂肪平衡過程的價值還需進一步探索。
我國已開展多項魚類脂肪沉積調節的遺傳機制,闡明Myostatin 通路與Pparγ 通路的協調脂肪沉積的機制將是重要的研究方向之一。
2.5 性別決定與分化
不像哺乳類動物性別決定機制比較穩定,在遺傳水平上性別分化比較清楚。魚類性別決定機制復雜,一些魚類受遺傳因素大而另一些受環境因素尤其是溫度的影響較大,遺傳決定也較哺乳類復雜得多[33]。龍娟等[17]總結,至目前已發現有10 種性別基因在不同魚類中起性別決定的作用,其中如Amhy、Amhr2、Gsdfy 和Gdf6y 等是TGF-β 基因超家族的成員。這些基因所在信號通路除了參與魚類的性別決定外,還參與魚類的性別分化等過程。王德壽團隊在羅非魚Oreochromis niloticus 性別決定與性別分化方面做了很多開創性研究并發表多篇論文[34-37]。
2.6 炎癥與免疫
這個家族調節腫瘤發生和防治的研究成果在水產養殖上可以借鑒,但水產養殖更關心的是傳染性疾病,對非傳染性疾病不是很重視,這當然與群體價值高而個體價值低有關。TGF-β 與免疫的相關作用是研究魚類疾病調控機制的重要方向,發表的文獻也多是考慮其免疫作用,目前多處于基因克隆和表達分析階段[38-40],未見對具體疾病治療上的研究報道。
2.7 肌間刺
鯉科魚類產量大,綜合效益比較高,但都有肌間刺這個讓消費者煩惱,甚至使小孩子受害的負面性狀。肌間刺作為硬骨魚類進化過程中的遺跡器官[41,42],缺失后對魚類的胚胎發育和骨骼發育都沒有明顯的負面影響[43,44]。因此,減少肌間刺會成為改善鯉科魚類品質的重要方向之一。推測,TGF-β 超家族的基因,尤其是BMP-SMAD 通路上的某個基因發生突變、或轉錄與表達過程有較大改變都可能是鯉科等幾個科的魚類具有肌間刺的原因。鑒定具體的基因突變或者轉錄表達決定機制上的變化是研究這個問題的主要目標。很多科的魚類具有肌間刺,是所有這些魚類在同一個通路上的基因產生突變,還是各自在特有的骨形成通路上的基因發生突變也是有意義的科學問題。
3 討論
3.1 TGF-β 基因超家族的分類
根據信號通路,TGF-β 基因超家族主要分為TGF-β/Activin/Nodal 和BMP/GDF 兩個亞家族(表1)。但這個家族基因數和功能不斷增多,也有人分為七個和九個亞家族。由于多肽聚合體有同源與異源之分[45],功能也有多功能和特異性功能的特點,無論從結構上分,還是從功能上分都不是很清晰,而且隨著研究的深入,這個家族的成員和功能還會增加,分類會更細。
研究表明,二聚神經生長因子(nerve growth factor,NGF)和血小板衍生生長因子(platelet derived growth factor,PDGF)的三維拓撲結構也顯示與TGF-β 相似,表明這些生長因子之間存在古老的結構保守性和相近的系統發育關系,這幾種信號通路上的蛋白家族或許可結合成為一個更大的超家族[46]。
這個家族受體的數量遠少于配體蛋白,但在生物過程的作用很大,受體主要與相關的配體結合包括與不同亞類的配體結合,使調控過程十分復雜,巨大的變異也使這個家族更多地參與經濟性狀在群體間和個體間的貢獻不同,也是經濟性狀研究的重點(表2)。
3.2 TGF-β 超家族調控是蛋白質調控性狀的典型
TGF-β 信號通路從轉錄到發揮生物學功能的生化過程極其復雜,可以作為了解蛋白調控性狀的典型案例,幫助人們深入認識“蛋白質機器”調控經濟性狀的復雜機制。這個生化過程基本上是:功能基因轉錄并翻譯出多肽類超前體,超前體由400 個左右氨基酸殘基組成(per-pro-TGF-β),殘基經過生物化學加工,如去掉殘基上的信號肽和N 端部分氨基酸殘基,形成有生物活性的前體如TGFβ1、GDF8 等,同源配體可聚合成進一步與受體結合的二聚體,即雙體如TGFβ1-TGFβ1。一些異源配體間也可聚合并同樣會與受體結合如TGFβ1-TGFβ2。一些亞家族間的前體也可形成異源二聚體如Vg1-Nodal。有些實驗表明:異源二聚體的活性更強[47];TGFβ 與受體結合后通過SMAD 蛋白達到信號向下傳導的任務即調控靶基因的轉錄與表達,獲得對目標性狀調控的結果,如免疫、骨骼生長、細胞增殖、性別決定等。
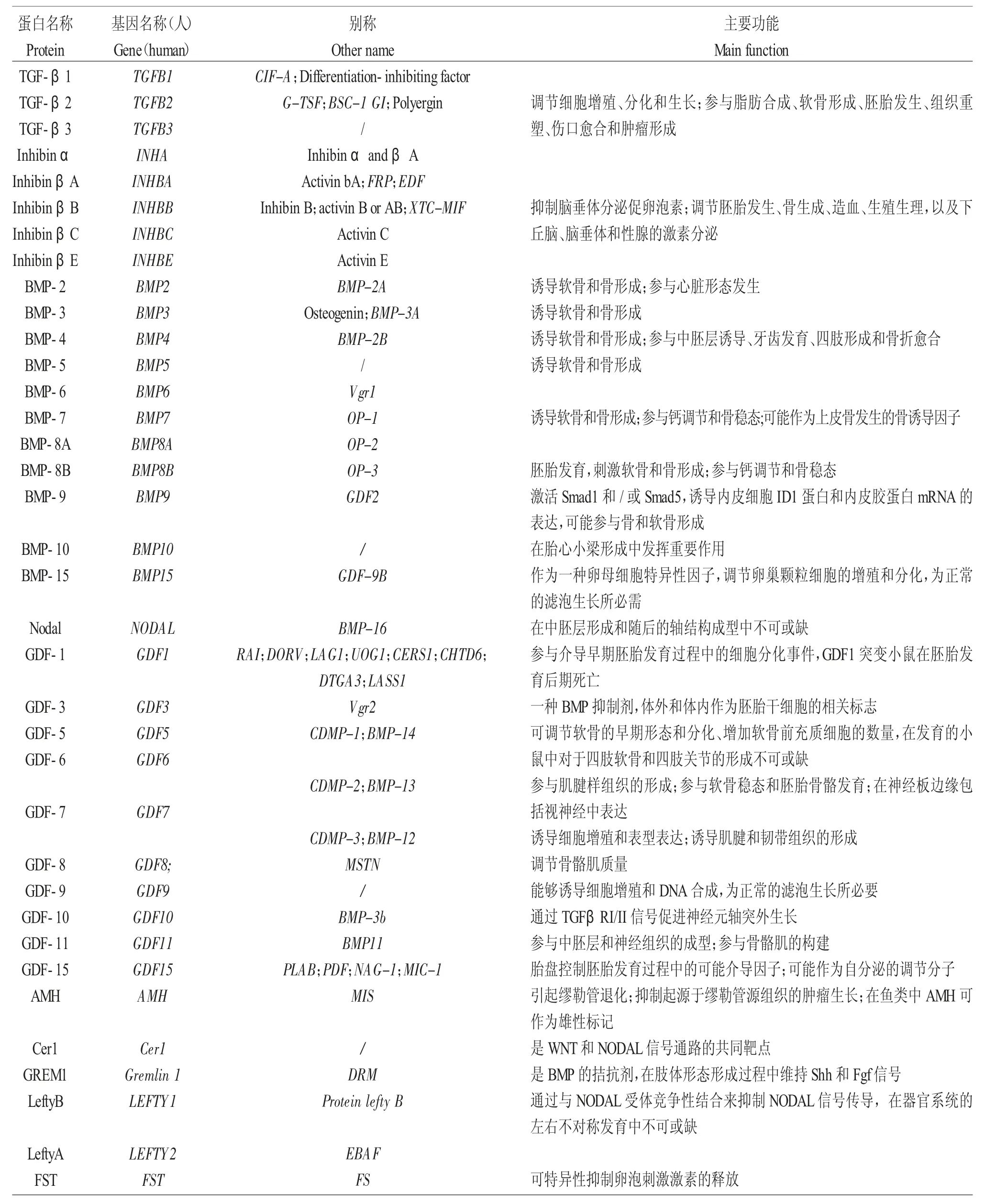
表1 TGF-β 超家族的基因及基本功能Tab.1 Genes and basic functions in the TGF-β family proteins
這個基因超家族比較復雜重要的原因之一是配體眾多,目前已發現40 多個,受體的數量遠少于配體的數量。但受體也種類繁多,如TGFβ 亞類有7 個受體,Activin/Inhibin 亞類有6 個受體,至2019年已發現總的受體為13 個[19],配體與受體間的組合十分復雜,一個二聚體可以和多個受體結合,一個受體也可以和多個配體結合[48]。

表2 TGF-β 超家族蛋白與受體的對應關系Tab.2 The relationship between TGF-β superfamily proteins and receptors
同家族多肽的結構相似性使很多信號通路間可以形成有效的串擾,通路間有相互作用也是多種蛋白信號通路的特征。TGFβ 家族配體和受體都比較多,共享同源序列比例大,更增加了家族內信號串擾的可能性[48,49]。
TGF-β 與其他信號通路相互作用的例子也有,譬如TGF-β 信號通路與GRP-R 之間相互作用。通常,TGF-β 家族中的分子識別通過與多個伙伴的相互作用網絡來實現[49,50];共受體,例如β 聚糖和內皮糖蛋白,也可以在細胞表面被切割,釋放其胞外域;釋放的果膠保持它們對配體的親和力,導致配體螯合和信號激活的抑制;協同受體,還提供了協調不同信號通路激活的機會,因此,除TGF-β 外,β 聚糖還可以結合bFGF 并協調調節FGF 和TGF-β 信號傳導[47]。這些增加了TGF-β信號通路與其他家族信號串擾的可能性。
3.3 關于TGF-β 基因超家族參與多種經濟性狀調控的思考
(一)基于TGF-β 基因超家族成員的基本功能。如肌肉生長抑素對生長的調控。這個肌肉生長的負調控因子一旦突變或者敲除,肌肉生長的內源平衡偏向于肌肉快速增加,可獲得肌肉組織增大的結果;BMP 對骨骼的調控也直接影響生長;再比如對性別的調控也是基于這個家族的基本功能即對細胞數量的調節能力,這里是調節性腺細胞的數量達到調控性別分化的結果,從而決定性別。
(二)通過調控標靶基因達到調節性狀的結果。如調控脂肪沉積的關鍵基因PPARγ 的轉錄水平調控脂肪;對飼料轉化率調控的細節還不清楚,但全基因組關聯分析鑒定到多個TGF-β 基因超家族成員參與了鯉飼料轉化效率的提高。
(三)通過調控垂體等全局性功能器官分泌的激素來調控具體的經濟性狀。如肌肉生長抑素就具有調節垂體激素釋放的功能。
TGF-β 基因超家族之所以能參與調控多種經濟性狀,關鍵點源于其基本的生物學特征即配體在多種組織中都有表達,多種細胞表面都存在受體,由此他們或是直接參與性狀的形成或者通過調節標靶蛋白配體基因表達來達到調節性狀的結果。
4 展望
綜上所述,TGF-β 代表了分泌性多肽因子的進化保守家族,可調節生理性胚胎發生和成年動物組織穩態的許多方面[51-53],據此可推斷其在動物主要經濟性狀的調控上也具有重要調節功能。雖然正在越來越多地關注這個基因超家族在畜禽和水產動物中的價值,但目前的研究還是初步和零散的,借助于人及藥物的研究成果,畜禽和水產養殖動物的研究和利用會得到快速發展。
由于TGF-β 是在多種器官和組織中表達,與特定組織調控的信號通路結合即調高或者抑制特定性狀相關基因的轉錄或表達,從而參與了該性狀的形成。醫學研究重點在于疾病形成過程的作用,藥物學研究重點是提高某種藥物或者減少某種藥物來阻止疾病的發展,農業育種工作者則重點在于首先了解這個信號通路如何在特定性狀形成過程中發揮了作用,哪些變異使其相關蛋白復合物上調,哪些變異使其下調,從而獲得可育種的標記和分子水平的其他工具如設計功能飼料等。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
