不同果實形狀番茄品種莖部內生細菌群落結構及代謝功能特征
2021-02-12 11:41:42陳思宇宋靜靜楊尚東
西南農業學報 2021年12期
孫 妍,陳思宇,肖 健,宋靜靜,楊尚東*
(1.廣西大學農學院/植物科學國家級實驗教學示范中心,廣西 南寧 530004;2.廣西北部灣海洋生物多樣性養護重點實驗室/北部灣大學海洋學院,廣西 欽州 535011)
【研究意義】番茄(SolanumlycopersicumL.)是世界范圍內廣泛栽培的重要蔬菜之一,含有豐富的有機酸、糖、礦物質、谷胱甘肽、番茄紅素和多種維生素等營養成分,有清除自由基、美白和延緩衰老等功效[1]。我國是世界上最大的番茄生產和消費國,種植番茄已成為我國部分農村的脫貧致富途徑[2]。番茄果實形狀是指示番茄品質的重要指標之一,也是番茄果實分級和育種的重要指標[3]。同時,隨著番茄采摘機器人研發的不斷深入,控制影響番茄果實形狀的因子正日益受到廣大科技人員的重視[4]。植物內生菌是能存活在植物各組織和器官內、與宿主植物關系和諧互利的一類微生物[5-6],具有促進宿主新陳代謝、防蟲防病、固氮、固磷和增強植株抗性等生物學作用[7-8],但對番茄品質尤其是果實形狀方面的影響尚不清楚。因此,解析果實形狀不同番茄品種植株莖部的內生細菌群落結構,探究內生細菌與果實形狀間的相關性,對構建新型番茄育種評價技術體系具有重要意義。【前人研究進展】Rodriguez等[9]利用Tomato analyzer可將番茄果實形狀分為扁平、矩形、橢圓形、牛心形、圓形、倒卵形、心形和長形,并發現SUN、OVATE、FAS和LC4個基因對果實形狀具有重要影響。番茄果實形狀除受基因調控外,還受外界環境、礦質營養水平、激素及病蟲害等諸多因素的綜合影響[10-13],如鉀元素不僅有助于番茄果實均勻著色,還有助于減少果實表面黃斑和畸形果[3],磷在番茄果實孕育中發揮著重要作用[14]。此外,在番茄果實發育過程中,植物激素尤其是生長素(IAA)、赤霉素(GA)、乙烯、脫落酸(ABA)和細胞分裂素(CK)也發揮著重要作用,如IAA通過刺激果皮細胞分裂觸發番茄坐果,且IAA濃度在果實中心部位最高,明顯高于其他果實部位[15-16];ABA可通過抑制乙烯的產生促進果皮細胞擴張,正向調控番茄細胞生長;GA處理可啟動果實發育功能,提高番茄果實坐果率,還可通過促進細胞分裂使果實膨大[17];ABA通過誘導氣孔關閉參與受寒冷和干旱脅迫植物的響應機制[18];CK可激活花前子房的細胞分裂并導致單性結實,調節果實生長[19]。Kumar等[16]研究還發現,SIIAA9基因表達的變化伴隨著乙烯相關基因表達的大量變化,乙烯通過與IAA進行協調作用參與果實坐果過程,因此內源激素IAA和乙烯含量水平對番茄結實的誘導至關重要;ABA合成減少、分解代謝增加或敏感性降低會導致植物氣孔關閉失敗,果實缺乏ABA會導致蒸騰作用增加,水分虧缺,抑制果實生長。已有研究證實,IAA[20]、CK[21]、ABA[22]、GA[23]和乙烯[24]等植物內源激素的產生與植物內生菌緊密相關。【本研究切入點】雖然前人對番茄果實形狀的形成開展了相關研究,但目前關于植物內生細菌對番茄果色、果形和果實甜度等果實品質影響的研究鮮見報道,特別是針對影響果實形狀形成特有內生菌屬的研究仍缺乏直接證據。【擬解決的關鍵問題】比較分析4種果實形狀番茄品種植株莖部的內生細菌群落結構及代謝功能特征,探究番茄果實形狀形成與植株內生細菌的關聯性,為構建番茄育種新型評價體系及有益微生物功能的開發利用提供理論依據和技術支撐。
1 材料與方法
1.1 試驗地概況
試驗于2020年3—6月在廣西大學農學院蔬菜基地(東經108°17′25″,北緯22°51′02″)設施大棚內進行。試驗地土壤理化性質:pH 5.68,有機質8.92 g/kg,全氮0.55 g/kg,全磷0.67 g/kg,全鉀7.51 g/kg,堿解氮15.27 mg/kg,速效磷0.67 mg/kg,速效鉀82.8 mg/kg。
1.2 試驗材料
供試橢圓形番茄品種為千禧(E)(臺灣農友種苗公司),扁平形番茄品種為桃太郎(F)(日本瀧井種苗株式會社),長型番茄品種為鳳珠(L)(臺灣農友種苗公司),圓球形番茄品種為愛麗(R)(日本瀧井種苗株式會社)。所有品種均同期育苗、定植及進行常規栽培管理。番茄進入果實成熟期時進行樣品采集。每個品種選取3株長勢一致的植株,使用經消毒的鐵鏟和剪刀隨機采集莖部樣品,送樣檢測植株莖部內生細菌。
1.3 試驗方法
1.3.1 樣品總DNA提取、PCR擴增和序列測定 利用FastDNA?Spin Kit for Soil試劑盒(MP Biomedicals,U.S)抽提總DNA,使用NanoDrop2000分光光度計(Thermo Fisher Scientific,U.S)測定DNA濃度和純度,最后凝膠電泳檢測DNA提取質量。細菌16S rRNA基因V5~V7高變區PCR擴增引物為799F:5′-AACMGGATTAGATACCCKG-3′和1193R:5′-ACGTCATCCCCACCTTCC-3′;進行PCR產物鑒定、純化及定量。Illumina MiSeq測序:以2%瓊脂糖凝膠進行PCR產物回收、純化(Extraction Kit)、Tris-HCl洗脫和電泳檢測,并用QuantusTMFluorometer (Promega,USA)對回收產物進行檢測定量。使用NEXTFLEX?Rapid DNA-Seq Kit進行建庫,利用上海美吉生物醫藥科技有限公司MiSeq PE300平臺進行測序[25]。
1.3.2 測定項目及方法 利用Uparse平臺對優化序列提取非重復序列,去除無重復的單序列,按照97%相似性對序列進行分類操作單元(OTU)聚類,得到OTU的代表序列,生成OTU表格。采用RDP classifier貝葉斯算法對序列進行分類學分析,對比Silva數據庫統計各樣本的群落物種組成。
本研究以Shannon與Simpson指數表征細菌群落多樣性,以Chao1和Ace指數指示細菌群落豐富度,以覆蓋率反映細菌群落的覆蓋度。利用R語言工具制作群落柱形圖進行分類水平上優勢物種組成和相對豐度分析;制作Venn圖統計分析樣本中所共有和特有的物種OTUs數,以深入了解番茄莖部內生細菌群落組成和結構特征;利用PICRUSt對16S rRNA基因V5~V7高變區擴增序列測序結果進行功能預測,根據KEGG數據庫的信息,獲得Pathway的2個水平信息及各功能類別的豐度表。
1.4 統計分析
試驗數據采用Excel 2019和SPSS 21.0進行統計分析。利用上海美吉生物醫藥科技有限公司的I-sanger云數據分析平臺在線數據進行番茄植株莖部內生細菌群落組成與結構及功能預測。
2 結果與分析
2.1 不同果實形狀番茄植株莖部內生細菌的OTUs聚類分析結果
由表1可知,在97%相似度水平對樣品序列進行OTU聚類,從4種果實形狀番茄的植株莖部內生細菌中共鑒定到31門84綱206目347科655屬1045種1660個OTU。其中,橢圓形番茄(E)莖部內生細菌群落由28門57綱155目269科473屬701種993個OTU組成;扁平形番茄(F)莖部內生細菌群落由24門55綱148目250科447屬660種951個OTU組成;長形番茄(L)莖部內生細菌群落由26門62綱158目267科468屬685種977個OTU組成;圓球形番茄(R)莖部內生細菌群落由25門62綱163目272科483屬723種1057個OTU組成。
2.2 不同果實形狀番茄植株莖部內生細菌的Alpha分析結果
由表2可知,4種果實形狀番茄植株莖部內生細菌的覆蓋率均在98.0%及以上,說明樣本測序數據足夠真實,可反映其內生細菌群落的多樣性;表征細菌群落多樣性的Shannon和Simpson指數間,以及表征豐富度的Chao1和Ace指數間均無顯著差異(P>0.05),說明4種果實形狀番茄植株莖部內生細菌均具有相似的多樣性和豐富度。
2.3 不同果實形狀番茄植株莖部內生細菌的群落組成分析
在門分類水平,不同果實形狀番茄植株莖部優勢內生細菌(豐度占比大于1.00%)的群落結構極相似,優勢細菌門均由放線菌門(Actinobacteriota)、變形菌門(Proteobacteria)、厚壁菌門(Firmicutes)、擬桿菌門(Bacteroidota)、酸桿菌門(Acidobacteriota)和綠彎菌門(Chloroflexi)組成(圖1)。其中,橢圓形果實番茄(E)植株莖部內生細菌優勢細菌門分類水平的豐度占比排序為放線菌門(45.22%)>變形菌門(34.28%)>厚壁菌門(14.30%)>擬桿菌門(1.72%)>酸桿菌門(1.36%)>綠彎菌門(1.31%),其他(Others)門的豐度占比為1.80%;扁平形番茄(F)植株莖部內生細菌優勢細菌門分類水平的豐度占比排序為變形菌門(46.97%)>放線菌門(32.32%)>厚壁菌門(11.79%)>擬桿菌門(3.32%)>酸桿菌門(2.04%)>綠彎菌門(1.17%),其他(Others)門的豐度占比為2.39%;長形番茄(L)植株莖部內生細菌優勢細菌門分類水平的豐度占比排序為變形菌門(48.15%)>放線菌門(34.96%)>厚壁菌門(11.28%)>酸桿菌門(1.27%)>擬桿菌門(1.19%)>綠彎菌門(1.17%),其他(Others)門的豐度占比為1.98%;圓球形番茄(R)植株莖部內生細菌優勢細菌門分類水平的豐度占比排序為放線菌門(49.28%)>變形菌門(28.53%)>厚壁菌門(15.13%)>酸桿菌門>(1.76%)>綠彎菌門(1.74%)>擬桿菌門(1.34%),其他(Others)門的豐度占比為2.23%。
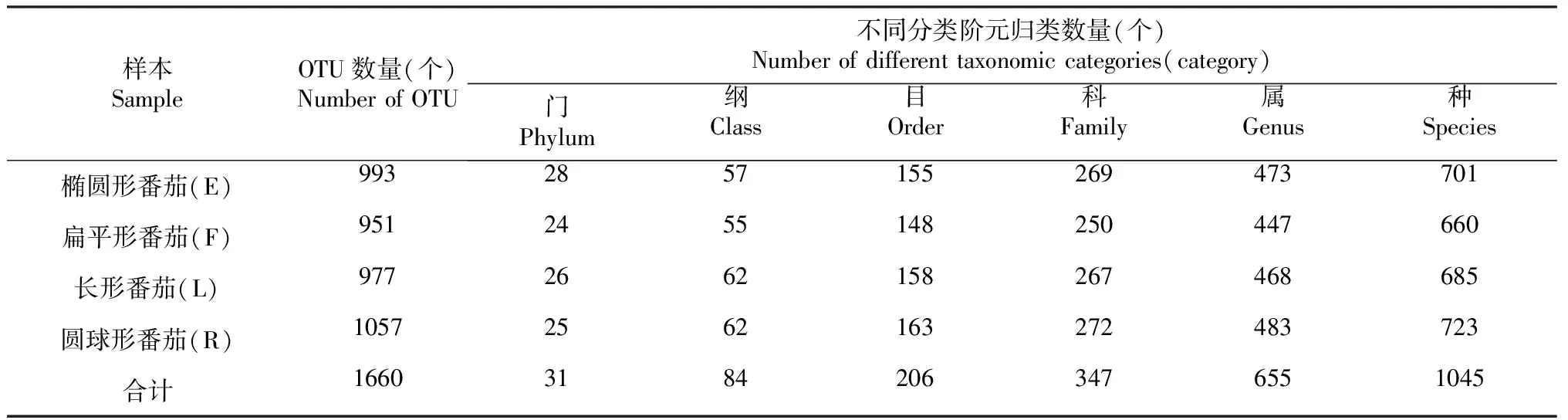
表1 不同果實形狀番茄植株莖部內生細菌在不同分類水平的數量
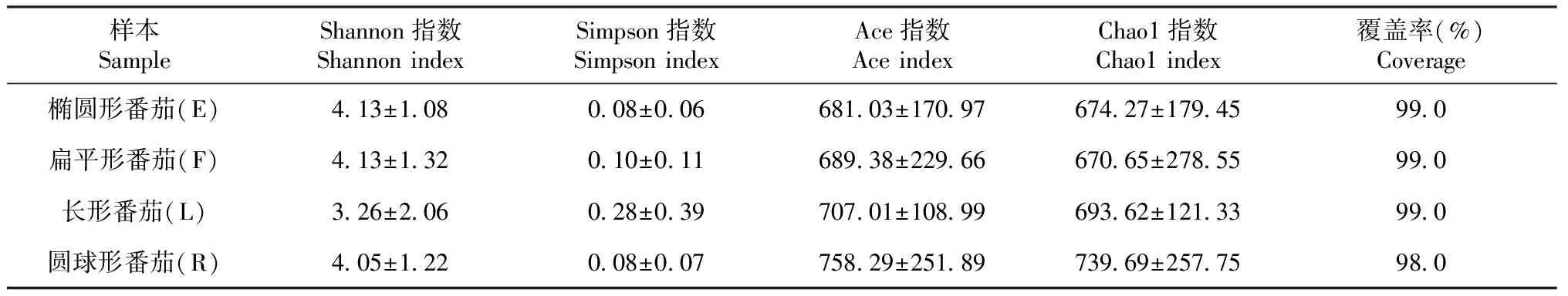
表2 不同果實形狀番茄植株莖部內生細菌的多樣性指數比較
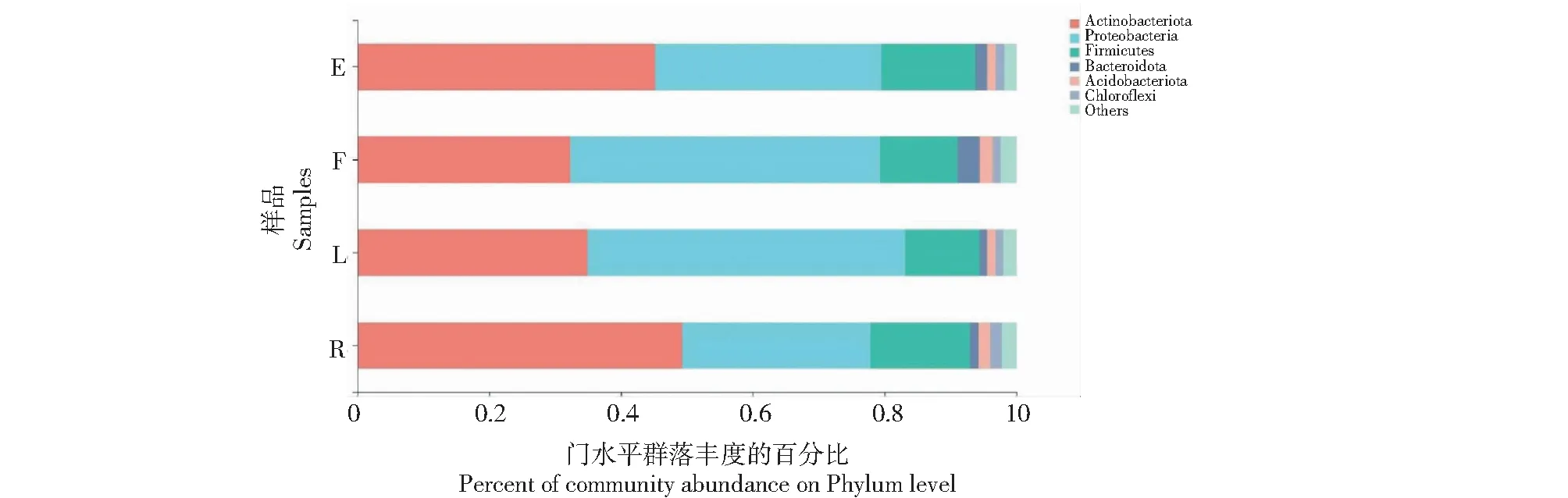
圖1 不同果實形狀番茄植株莖部優勢內生細菌門分類水平的相對豐度占比比較Fig.1 Comparison of the relative abundance of dominant endophytic bacteria in stems of tomato plant with different fruit shapes at phylum level
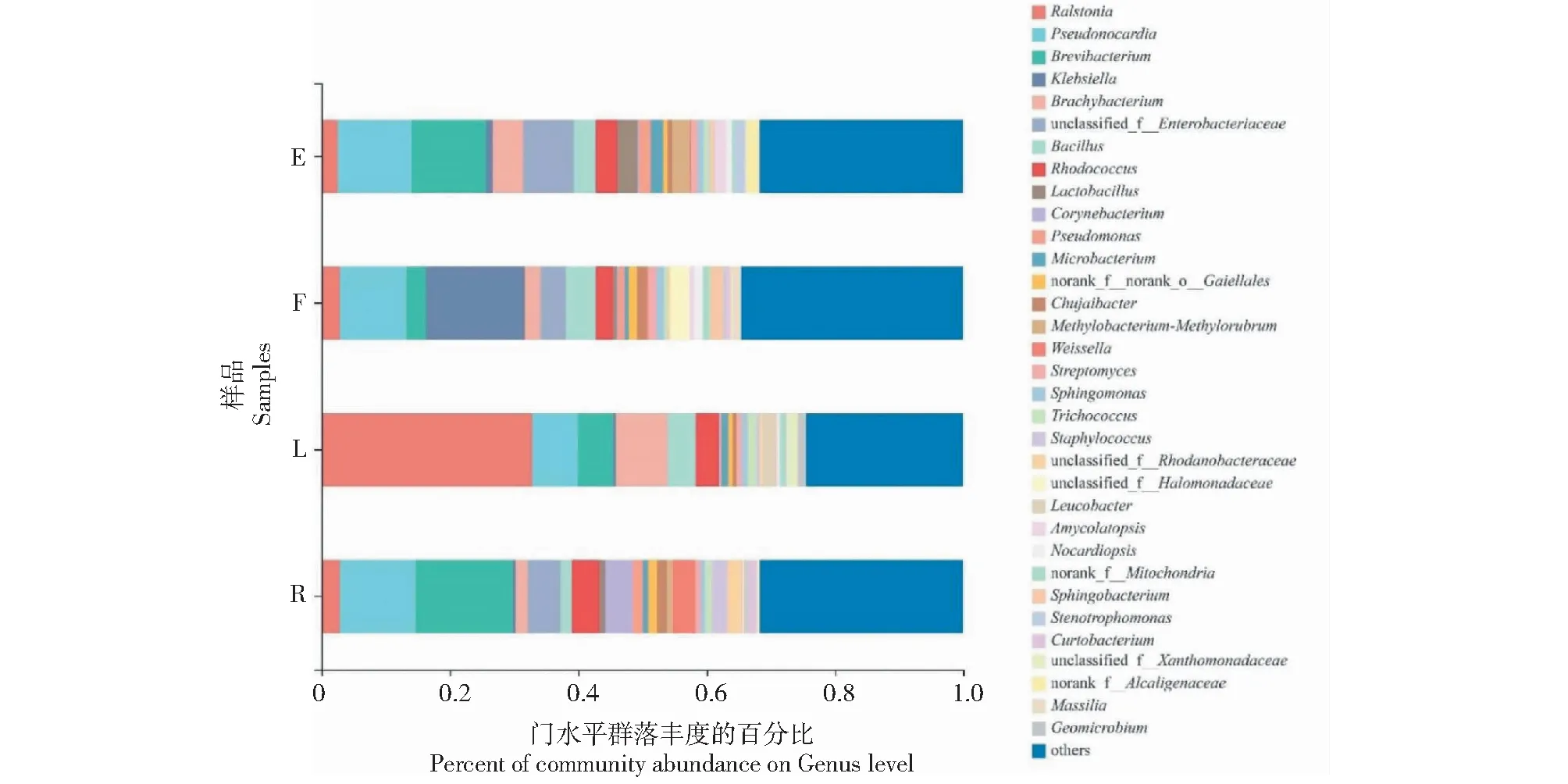
圖2 不同果實形狀番茄植株莖部優勢內生細菌屬分類水平的相對豐度比較Fig.2 Comparison of the relative abundance of dominant endophytic bacteria in stems of tomato plant with different fruit shapes at genus level
在屬分類水平,不同果實形狀番茄植株莖部共有的優勢內生細菌屬(豐度占比大于1.00%)均為青枯菌屬(Ralstonia)、假諾卡氏菌屬(Pseudonocardia)、短頸細菌屬(Brevibacterium)、短狀桿菌屬(Brachybacterium)、芽孢桿菌屬(Bacillus)和紅球菌屬(Rhodococcus)(圖2)。其中,青枯菌屬在果實長

表3 不同果實形狀番茄植株莖部優勢內生細菌屬分類水平的相對豐度占比比較
形番茄植株莖部的細菌屬中的豐度占比達32.83%,而在橢圓形番茄、圓球形番茄和扁平形番茄植株莖部的細菌屬中的豐度占比分別僅為2.55%、2.83%和2.83%。此外,果實橢圓形番茄植株莖部特有的優勢內生細菌屬為乳酸桿菌屬(Lactobacillus)、甲基桿菌—甲基羅布氏菌屬(Methylobacterium-Methylorubrum)、擬無枝酸菌屬(Amycolatopsis)、寡養單胞菌屬(Stenotrophomonas)和未分類產堿桿菌屬(norank_f__Alcaligenaceae);果實扁平形番茄植株莖部特有的優勢內生細菌屬為鞘脂單胞菌屬(Sphingomonas)、未分類鹽單胞菌屬(unclassified_f__Halomonadaceae)、擬諾卡氏菌屬(Nocardiopsis)、norank_f__Mitochondria、鞘氨醇桿菌屬(Sphingobacterium)和Massilia;果實長形番茄植株莖部特有的優勢內生細菌屬為Leucobacter、未分類黃色單胞菌屬(unclassified_f__Xanthomonadaceae)和地微桿菌屬(Geomicrobium);果實圓球形番茄植株莖部特有的優勢內生細菌屬為棒狀桿菌屬(Corynebacterium)、魏斯氏菌屬(Weissella)、葡萄球菌屬(Staphylococcus)、unclassified_f__Rhodanobacteraceae和短小桿菌屬(Curtobacterium);未分類的大腸桿菌屬(unclassified_f__Enterobacteriaceae)在果實長形番茄植株中不存在,但在果實橢圓形、圓球形和扁平形番茄植株莖部的細菌屬中的豐度占比分別達7.91%、5.12%和3.95%(表3)。
2.4 不同果實形狀番茄植株莖部細菌群落的物種Venn圖分析
由Venn圖分析結果(圖3)可知,在OTU分類水平,果實橢圓形番茄的植株莖部與其他果形番茄的植株莖部共有內生細菌OTUs為993個,特有內生細菌OTUs為154個;果實扁平形番茄的植株莖部與其他果形番茄的植株莖部共有內生細菌OTUs為951個,特有內生細菌OTUs為109個;果實長形番茄的植株莖部與其他果形番茄的植株莖部共有內生細菌OTUs為977個,特有細菌OTUs為142個;果實圓球形番茄的植株莖部與其他果形番茄的植株莖部共有內生細菌OTUs為1057個,特有細菌OTUs為184個;四者共有內生細菌OTUs為501個。
從圖4可看出,在屬分類水平,4種果實形狀番茄的植株莖部中共有內生細菌屬310個;果實橢圓形番茄的植株莖部特有內生細菌屬42個;果實扁平形番茄的植株莖部特有內生細菌屬23個;果實長形番茄的植株莖部特有內生細菌屬為38個;果實圓球形番茄的植株莖部特有內生細菌屬為49個。
2.5 PICRUSt功能預測
為了獲得不同果實形狀番茄植株莖部內生細菌的功能,本研究利用高通量測序技術和KEGG數據庫進行比對,發現4種果實形狀番茄的植株莖部內生細菌在一級功能層共有代謝(Metabolism)、環境信息處理(Environmental information processing)、遺傳信息處理(Genetic information processing)、細胞過程(Cellular processes)、人類疾病(Human diseases)和有機系統(Organismal systems)6類生物代謝通路(圖5)。其中,代謝均為4種果實形狀番茄的植株內生細菌主要功能組成,在橢圓形、扁平形、長型和圓球形番茄植株莖部的6類生物代謝通路中占比分別為78.16%、77.14%、77.95%和78.82%;環境信息處理在橢圓形、扁平形、長型和圓球形番茄植株莖部的6類生物代謝通路中占比分別為6.24%、7.07%、6.20%和5.72%;遺傳信息處理在橢圓形、扁平形、長型和圓球形番茄植株莖部的6類生物代謝通路中占比分別為5.75%、5.55%、5.65%和6.06%;細胞過程在橢圓形、扁平形、長型和圓球形番茄植株莖部的6類生物代謝通路中占比分別為4.41%、4.81%、4.51%和4.10%;人類疾病在橢圓形、扁平形、長型和圓球形番茄植株莖部的6類生物代謝通路中占比分別為3.50%、3.65%、3.76%和3.35%;有機系統在橢圓形、扁平形、長型和圓球形番茄植株莖部的6類生物代謝通路中占比分別為1.94%、1.78%、1.94%和1.94%。可見,代謝功能是4種果實形狀番茄的植株內生細菌最主要功能。此外,橢圓形和圓球形番茄植株莖部內生細菌的代謝功能預測基因拷貝數高于相應的扁平形和長型番茄;不同果實形狀番茄植株莖部內生細菌基因一級功能層預測基因種類無顯著差異,但基因拷貝數占比存在差異。

圖3 不同果實形狀番茄植株莖部內生細菌OTUs分類水平的Venn圖分析結果Fig.3 Venn diagram of endophytic bacteria at OTUs level in stem of tomato plant with different fruit shapes
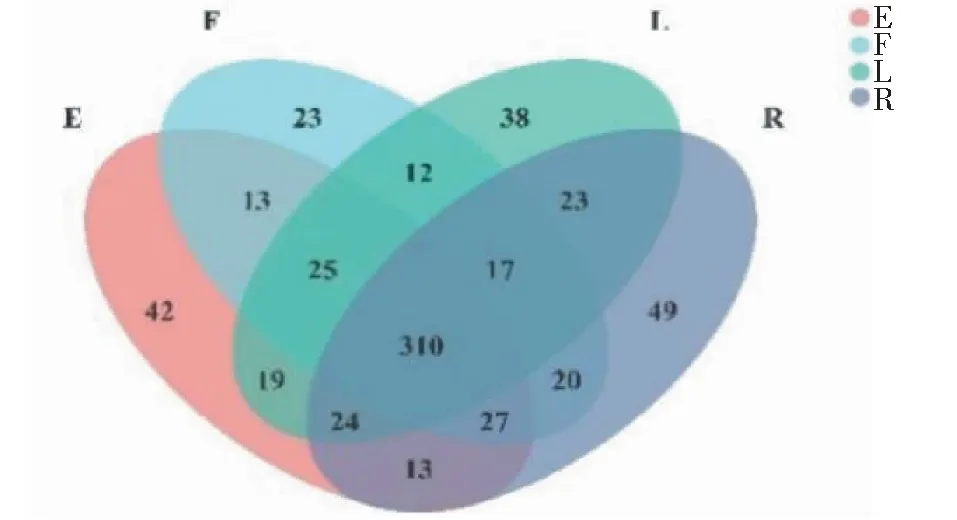
圖4 不同果實形狀番茄植株莖部內生細菌屬分類水平的Venn圖分析結果Fig.4 Venn diagram of endophytic bacteria at genus level in stem of tomato plant with different fruit shapes
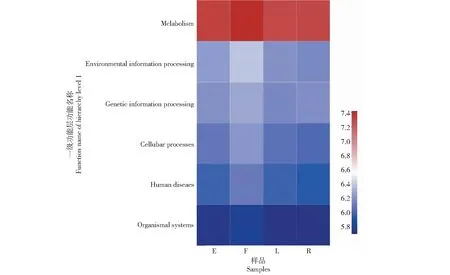
圖5 預測功能基因在不同番茄植株莖部內生細菌間的差異(一級功能層)Fig.5 Predicting differences in functional genes among endophytic bacteria in the stems of different tomato plants(hierarchy level 1)
對4種果實形狀番茄的植株莖部內生細菌基因進行二級功能預測分析,結果(圖6)發現其二級功能層均主要由全貌總述圖(Global and overview maps)、碳水化合物代謝(Carbohydrate metabolism)、氨基酸代謝(Amino acid metabolism)、能量代謝(Energy metabolism)、輔因子和維生素的代謝(Metabolism of cofactors and vitamins)、膜轉運(Membrane transport)、信號傳導(Signal transduction)、異生物質的生物降解和代謝(Xenobiotics biodegradation and metabolism)和細胞群落—原核生物(Cellular community-prokaryotes)等46個子功能組成,其中,果實圓球形番茄植株莖部內生細菌的全貌總述圖、氨基酸代謝、輔因子和維生素的代謝、類脂物代謝作用(Lipid metabolism)和翻譯(Translation)等15個二級功能層預測基因拷貝數占比較多;果實扁平形番茄植株莖部內生細菌的碳水化合物代謝、能量代謝、膜轉運、信號傳導和細胞群落—原核生物等14個二級功能層預測基因拷貝數占比較多;果實長形番茄植株莖部內生細菌的異生物質生物降解和代謝、細胞生長和死亡(Cell growth and death)、癌癥:概述(Cancer:overview)和神經退行性疾病(Neurodegenerative disease)等13個二級功能層預測基因拷貝數占比較多;果實橢圓形番茄植株莖部內生細菌的老化(Aging)、轉運和分解代謝(Transport and catabolism)、免疫系統(Immune system)和免疫性疾病(Immune disease)4個二級功能層預測基因拷貝數占比最多。
綜上所述,雖然4種果實形狀番茄植株莖部內生細菌基因的二級功能層預測基因種類相近,但基因拷貝數四者間存在明顯差異。由此推測,不同果實形狀番茄植株間內生細菌群落組成、結構及其功能基因種類和豐度的差異可能是造成植株內部合成代謝途徑、占比、營養成分、酶和激素種類及其含量水平產生差異的因素,進而使得不同番茄品種果實呈現不同的形狀。
3 討 論
已有研究表明,內生細菌寄居于植物組織中,與寄主植物建立了和諧互作的良好關系,具有固氮、促進植物生長、抗病蟲害及抗病原真菌等作用,還可改變酶活性,進而調節寄主植物的新陳代謝[26-29];在涉及番茄果實形狀的4個基因中,SUN基因編碼IQD家族的鈣調素結合蛋白影響果實伸長,OVATE基因編碼OVATE家族蛋白調控轉錄而影響果實伸長,LC基因主要控制分生組織大小及腔室數量,FAS基因編碼YABBY家族調控腔室數量誘導果實扁平或卵圓形伸長,但番茄果實形狀除受品種基因調控外,還受營養、內源激素水平和環境(光照、溫度和水分)等因素影響[30]。近年來,隨著人們認知水平的不斷提升,已發現植物內源激素的形成與微生物間存在緊密聯系,如IAA和CK含量與酸桿菌門和綠彎菌門細菌豐度占比間呈顯著正相關,與厚壁菌門細菌豐度占比呈顯著負相關;ABA含量與放線菌門細菌豐度占比呈顯著正相關,與變形菌門、綠彎菌門細菌豐度占比呈顯著負相關[31]。本研究結果與上述研究結果相似。
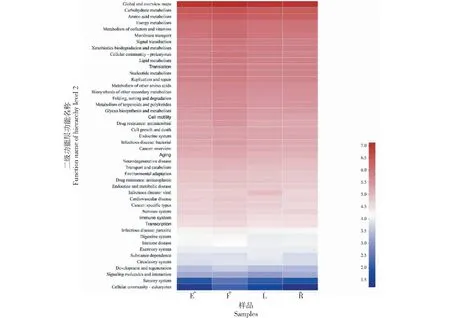
圖6 預測功能基因在不同番茄植株莖部內生細菌間的差異(二級功能層)Fig.6 Predicting differences in functional genes among endophytic bacteria in the stems of different tomato plants(hierarchy level 2)
本研究結果表明,在門分類水平,不同果實形狀番茄植株莖部的內生細菌群落結構相似,優勢細菌門類均由放線菌門、變形菌門、厚壁菌門、擬桿菌門、酸桿菌門和綠彎菌門細菌組成,但優勢細菌門的豐度占比在不同果實形狀番茄植株莖部表現各異。其中,果實長形和扁平形番茄植株莖部中豐度占比最高的優勢內生細菌門為變形菌門,占比分別達48.15%和46.97%;果實橢圓形和圓球形番茄植株莖部中豐度占比最高的優勢內生細菌門為放線菌門,占比分別達45.22%和49.28%,而變形菌門細菌的豐度占比僅分別為34.28%和28.53%。由于ABA含量與放線菌門細菌豐度占比呈正相關、與變形菌門細菌豐度占比呈負相關,由此推測,相比于果實橢圓形和圓球形番茄品種,果實長形和扁平形番茄品種植株莖部可能會形成ABA含量更高的環境;內源ABA可抑制番茄果實縱向生長[32],因此ABA可調控番茄果實形狀傾向于形成橢圓形或圓球形。
乳酸桿菌屬[33]、寡養單胞菌屬[34]、葡萄球菌屬[35-36]和短小桿菌屬[37-38]細菌是果實橢圓形或圓球形番茄品種植株莖部特有的優勢內生細菌屬。其中,乳酸桿菌屬細菌是一類通過產生乳酸降低內生環境pH、有益于植株健康的微生物,寡養單胞菌屬細菌具有ACC脫氨酶活性,可調控乙烯的合成,進而增強植株抗逆性,葡萄球菌屬可將難溶性有機/無機磷轉化成有利于植株吸收的可溶性磷酸鹽,改善作物磷素營養供應,提高土壤肥力,短小桿菌屬具有固氮能力,可提高植株氮利用率,增加果實產量的效果。本研究中,在屬分類水平,青枯菌屬、假諾卡氏菌屬、短頸細菌屬、短狀桿菌屬、芽孢桿菌屬和紅球菌屬細菌均是4種果實形狀番茄的植株莖部中共有優勢內生細菌屬;果實形狀不同番茄的植株莖部中也富集了各自特有優勢內生細菌屬,其中,果實橢圓形番茄的植株莖部特有優勢內生細菌屬為乳酸桿菌屬、甲基桿菌—甲基羅布氏菌屬、擬無枝酸菌屬、寡養單胞菌屬和未分類產堿桿菌屬;果實扁平形番茄的植株莖部特有優勢內生細菌屬為鞘脂單胞菌屬、未分類鹽單胞菌屬、擬諾卡氏菌屬、norank_f__Mitochondria、鞘氨醇桿菌屬和Massilia;果實長形番茄的植株莖部特有優勢內生細菌屬為Leucobacter、未分類黃色單胞菌屬和地微桿菌屬;果實圓球形番茄的植株莖部特有優勢內生細菌屬為棒狀桿菌屬、魏斯氏菌屬、葡萄球菌屬、unclassified_f__Rhodanobacteraceae、短小桿菌屬。由此推斷,本研究中果實橢圓形或圓球形番茄植株莖部能富集有別于果實長形或扁平型番茄植株莖部的特有優勢內生細菌屬,因此可形成諸如內源激素種類與含量、汁液pH及植株內部營養水平等環境因子有異于果實長形或扁平型番茄植株的內部環境,進一步誘導果實形狀不同番茄品種中不同功能基因表現出顯性或隱性狀態,促使該環境下“啟動”的功能基因調控形成不同形狀的番茄果實。
4 結 論
4種果實形狀番茄的植株莖部均富集共有優勢內生細菌屬,各自還富集了特有優勢內生細菌屬,但共有優勢內生細菌屬的豐度占比各異。其中,特有優勢內生細菌屬及其代謝功能參與調控或影響番茄果實形狀的形成進程。因此,番茄植株莖部內生細菌群落特征有望作為指示番茄果實形狀的生物學指標。
