2年生云南松實生苗生物量家系間變異研究*
2021-02-24 09:01:56魏巍蔡年輝李根前鄭畹許玉蘭李孫玲陳詩王磊孫睿
西部林業科學 2021年1期
關鍵詞:分配
魏巍,蔡年輝,李根前,鄭畹,許玉蘭,李孫玲,陳詩,王磊,孫睿
(1.云南省林業和草原科學院,云南 昆明 650201;2.西南林業大學西南山地森林資源保育與利用教育部重點實驗室,云南 昆明 650224)
植物生長受環境中可用資源的限制和影響,理想的生物量分配理論假定植物將生物量分配到能夠獲得最受限制資源的器官,以便實現最快的生長[1]。濃密的下層植物或干旱貧瘠的生境可以影響更新層幼苗的生存,而苗木生物量的分配策略,決定了其競爭能力和將來的生長狀況[2]。特定條件下,苗木對各構件器官生物量的投入和比例平衡能力,可視作一組苗木質量評價指標或待選擇的優良性狀。
云南松(Pinusyunnanensis)具有適應性強、耐干旱瘠薄等優良特性,是中國西南地區人工造林的重要先鋒樹種,該樹種占其主要分布區云南省全省林地面積的19.56%、森林總蓄積量的14.28%[3-5]。但是其天然更新的幼苗均存在“蹲苗期”,即幼苗地上部分生長較慢,很大程度影響著造林成活率和保存率[6]。優質壯苗是營造培育優質高產人工林的基礎[7],實際林業生產中一般根據苗高、地徑或綜合的苗木質量指標對苗木進行分級使用[8]。不同層次的選育工作和苗木分類生產需強化具有生物學特性的變異規律研究,摸清遺傳變異規律和主要性狀的遺傳參數,從而選出不同生態區的優良種源、林分和家系[9]。當前,云南松的遺傳改良水平仍處于較為初級的階段,良種使用率尤其優良家系種苗使用率還較低。進行各層次遺傳變異的測定勢在必行,而家系層次的遺傳變異研究是遺傳改良工作的重要組成部分。
因此,本研究對20個半同胞家系的云南松實生苗生物量累積與分配在家系層次的變異規律進行研究,以期為科學評價云南松苗木質量和發揮遺傳改良的潛在增益提供理論依據。
1 材料與方法
1.1 試驗材料
本試驗所用半同胞家系云南松種子采自云南省永仁縣林業局國營白馬河林場云南松母樹林基地內20個云南松健康單株,每株之間的最小水平距離大于100 m,家系平均千粒重和標準差為15.036±2.676g。
1.2 試驗設計
本研究采用單因素對比設計,除了種子來自不同的母樹外,其他條件均相同。田間排列采用完全區組法,并結合順序錯位法進行排列,設3次重復,每次重復由1個區組組成,1個區組劃分為20個小區,每個家系為1個小區,完整試驗共包含60個小區。
1.3 育苗方法及苗木管理
試驗地設置在西南林業大學育苗圃(102°45′41″ E,25°04′00″ N),海拔1 945 m,屬北亞熱帶半濕潤高原季風氣候,年平均氣溫14.7 ℃,年降水量700~1 100 mm,年平均相對濕度68.2%。苗床采用高床設計,寬1.0 m,下層生土,播種前2 d噴灑0.5%高錳酸鉀溶液及10.0%多菌靈溶液對下層土消毒,上層鋪4~5 cm取自昆明樹木園的云南松林表土。按照DB53/T249-2008《林木育苗技術規程》[10]進行種子調制處理并用馬勃菌拌種。育苗當年的5月按設計進行點播,每個家系播種14行、每行16株,株行距皆為5 cm×5 cm。播種后采用0.5~1.0 cm厚細粒云南松林下土覆蓋,噴灌裝置霧狀澆水20 min,土壤濕透蓋塑料拱膜。苗期第1—2月保持苗床土壤濕潤無雜草。大部分苗木長出真葉前,為防止苗木灼傷或猝倒,根據膜內溫度定期開膜透風降溫,此后去除薄膜。為預防常見病害,在苗木完全木質化前每隔20~30 d用0.5%多菌靈溶液噴灑1次,遇蟲害則用1%敵百蟲溶液噴灑。
1.4 數據調查
到2 a生苗木生長結束時,即播種后第2年12月底,從每個家系中隨機抽取20株苗木,采用全挖法測定生物量。挖出后每株分根、莖、葉不同器官按家系和單株編號帶回實驗室,105 ℃殺青30 min,80 ℃恒溫下烘干至恒定質量,精確至0.001 g。
1.5 家系生物量變異及遺傳特性
1.5.1 方差分量百分比
方差分量百分比可以體現數據集變異來源,本研究借鑒群體或居群遺傳多樣性分析手段[11-14],將其應用在家系層面變異特性分析。方差分量百分比Pσf可以表示家系變異占總變異的比例。
Pσf=σf/σt
①
式中:Pσf為方差分量百分比,σf和σt分別為家系間和總體的方差分量。
1.5.2 表型分化系數
表型分化系數Vσf可以表示家系間表型性狀變異的值。
Vσf=σf/(σf+σe)
②
式中:σe為家系內的方差分量,即環境方差分量。
1.5.3 變異系數
變異系數(coefficient of variation,下文皆以“CV”表示)是表示一組數據變異程度大小的指標之一,為標準差S與平均數的比值的百分數,可以反映單位均值上的離散程度,常用在2個總體均值不等的離散程度的比較上。由于不同家系表型性狀指標的均值有所差異,本研究采用CV來輔助評價不同家系各表型性狀的變異程度。根據式③和式④計算不同家系各表型性狀的平均值和標準差S,并依式⑤計算CV。
③
④

⑤
式中:x為生物量指標實測值,n為每個家系測定的株數。
1.5.4 半同胞家系遺傳力
苗木的表型性狀傳遞給其后代的潛力稱為遺傳傳遞力或遺傳力,本研究采用的遺傳力估算方法,如式⑥:
⑥
式中:Sf為家系間離差平方和,Se為家系內離差平方和,r為供測家系總數,n為單個生物量指標供測樣品的總數,F值為家系間和家系內供測生物量的均方之比值。
1.5.5 遺傳增益
遺傳增益(△G)在林木遺傳育種的分析應用中實際上是表示人工選擇取得改良效果的指標之一[15],即入選親本的子代平均表現值距親本群體平均型值的離差R與親本群體平均型值的百分比。根據代算可得:
⑦
式中:S′為選擇差,hf2為遺傳力。本研究借鑒林木遺傳育種遺傳增益△G這個指標,并假設選擇后的選擇差為家系表型性狀的1倍標準差,估算在此選擇強度下的遺傳增益,模擬選擇后的改良效果。試驗數據用Excel 2013進行統計,SPSS 13.0進行家系生物量指標的分析比較。
2 結果與分析
2.1 2 a生實生苗生物量累積、分配與生產力關系
2 a生實生苗生產力與各構件器官生物量的累積密切相關,由圖1可知,實生苗各構件器官生物量累積量隨總生物量變大而呈現明顯的增加趨勢。

圖1 2 a生苗木不同構件器官生物量累積量與總生物量大小的關系Fig.1 The relations between modular biomass effort and total biomass of two-year P.yunnanensis
除生物量的累積外,不同構件器官的生物量分配比例還隨總生物量的變化呈現一定的規律。由圖2可知,如總生物量分配于針葉和莖生長的比例分布范圍,隨苗木總生物量的增大而逐漸減小,生物量較高的實生苗,針葉生物量分配的范圍僅在50%附近,莖生物量分配的范圍在40%附近;生物量較低的實生苗,其根系的分配比例較小,高生產力的苗木根系分配比例普遍高于生產力較低的苗木;不同生產力苗木的地上與地下生物量之比有所不同,較低生產力苗木的地上地下生物量比普遍高于較高生產力的苗木,較高生產力的苗木,其地上地下生物量比大部分處于較低水平,在5~10 g之內分布廣泛。

圖2 2 a生苗木不同構件器官生物量分配比例與總生物量大小的關系Fig.2 The relations between modular biomass effort ratio and total biomass of two-year P.yunnanensis
2.2 2 a生實生苗生物量變異概況
2 a生苗木各構件器官生物量累積量分別為,根干重(0.774±0.281)g、莖干重(3.119±1.488)g、葉干重(3.609±1.586)g、地上干重(6.728±2.908)g、全株干重(7.503±3.425)g。多重比較結果表明(表1),在P<0.05的顯著水平下各構件器官生物量累積量可分為2組,全株干重最大的為11號、12號、14號、16號家系,最小的為1號、6號、8號家系;地上干重最大的為12號、14號、16號家系,最小的為1號、6號、8號家系;徑干重最大的為2號、8號、11號、15號家系,最小的為6號家系;葉干重最大的為2號、10號、11號、12號、14號、16號家系,最小的為1號家系;根干重最大的為12號、16號家系,最小的為1號家系。由此可知,12號家系生物量積累最多,6號家系生物量積累最少。

表1 2 a生云南松各家系生物量累積Tab.1 Biomass effort among families of two-year P.yunnanensis g
2 a生苗木各構件器官生物量分配比例分別為,根分配(9.32±3.86)%、莖分配(41.75±7.62)%、葉分配(48.93±7.68)%、地上地下生物量比(13.96±5.43)。多重比較結果(表2)表明,在P<0.05的顯著水平下根系生物量分配比例可分為2組,最大為12號、16號家系,最小為1號家系。在P<0.05的顯著水平下,莖分配、葉分配和地上地下生物量比可以分為3組。徑分配最大的為1號、3號家系,最小的為6號、12號、16號家系;葉分配最大的為10號家系,最小的為17號家系;地上地下生物量比最大的為5號、8號、9號家系,最小的為12號家系。

表2 2 a生云南松各家系生物量分配Tab.2 Biomass allocation among families of two-year P.yunnanensis
2.3 2 a生實生苗生物量變異來源
如表3所示,各構件器官和總生物量的累積量在家系間的方差分量占總方差的百分比范圍在12.770 3%~33.028 5%,表明其生物量累積量在家系間有一定程度的變異,其中葉干重變異受家系層次的影響最小,莖干重受家系層次的影響最大。總生物量對各構件器官生長分配比例的方差分量占總方差的百分比范圍在17.7424%~39.0348%,其中,家系對根分配變異的影響最大,對地上地下生物量比的變異影響最小。
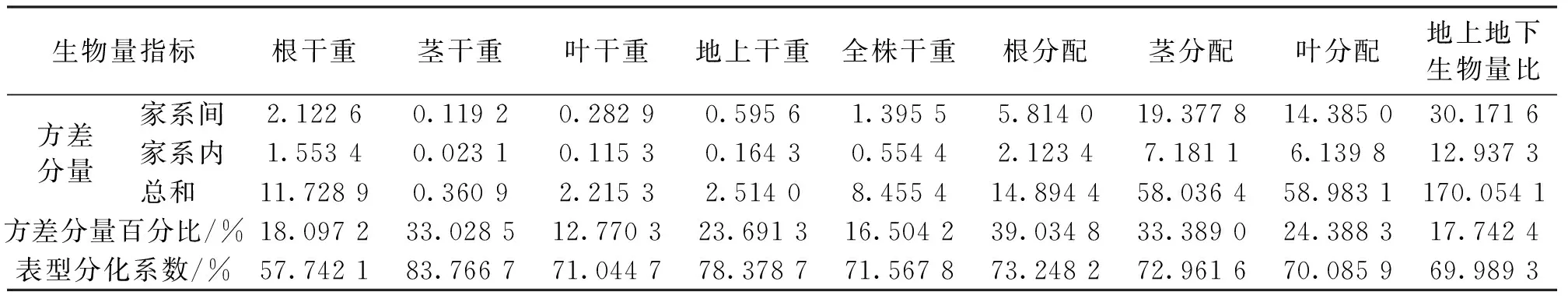
表3 2 a生云南松各家系生物量的方差分量與表型分化系數Tab.3 Variance components and differentiation coefficients of phenotypic traits for biomass among families of two-year P.yunnanensis
2.4 2 a生實生苗生物量變異特征
如表4所示,2 a生各構件器官生物量累積量的平均CV為39.70%~55.40%。其中,根干重變異最為豐富,11號家系根干重的變異程度最小,16號家系的變異程度最大;葉干重的分化較小,CV為39.70%,其2號家系的變異程度最小,9號家系的變異程度最大。

表4 2 a生云南松各家系生物量的變異系數Tab.4 Coefficient of variations for biomass among families of two-year P.yunnanensis %
構件器官生物量分配比例和地上地下生物量比的平均CV依次為39.12%、11.25%、7.78%和44.36%。根分配比例的變異程度最低,12號家系根分配比例的CV最小(4.52%),4號家系最大(22.14%);地上地下生物量比的變異最為豐富,其19號家系的CV最小(18.07%),2號家系CV最大(48.17%)。
2.5 2 a生實生苗生物量遺傳特性
如表5所示,2 a生云南松苗木各構件器官生物量累積量的遺傳力在0.39~0.45的范圍,遺傳力都略小于0.50,遺傳力低于中等水平,表明變異受遺傳因素的影響水平較低,其中根干重的遺傳力最高,葉干重最低。

表5 2 a生云南松生物量的遺傳參數估算Tab.5 Estimates of genetic parameters for biomass of two-year seedling
生物量分配比例和地上地下生物量比的家系遺傳力都較生物量累積指標的遺傳力高,在0.46~0.62范圍,僅莖分配比例的遺傳力低于中等水平,其余指標的變異受遺傳因素的控制力較大,葉分配的家系遺傳力最高(0.62)。
如果通過人工選擇,各構件器官的累積與分配能在不同程度獲得遺傳增益。選擇地上地下生物量比和根干重可以獲得的增益較高,分別為14.21%和10.26%;選擇根分配能獲得的增益最小,僅為5.51%。
3 結論與討論
2 a生云南松實生苗各構件器官生物量累積與生產力的關系十分明確,即構件器官生物量累積越多,苗木的總生物量累積就越多,生產力也就越大。但總生物量大小與不同構件器官生物量分配比例有著各種微妙的關系。不同構件器官間生物量分配比例的不同會影響各構件器官生物量的累積,也會一定程度影響苗木的形態特征,最終影響苗木的生產力[16]。以往學者也在群落、個體等水平十分關注有關植物形態指標的變異規律,在草本植物、針葉樹種的形態指標調控和變異規律等方面開展相關研究[17-19]。作為可以描述植物形態的指標之一,地上地下生物量比可用于對苗木質量、林木生長特點、碳循環分配等問題進行探討,因此了解這類性狀在家系間的變異規律十分有必要。本研究發現絕大部分與苗木生產力和生態適應能力緊密相關的指標,在家系間存在具有統計學意義的變異現象,且家系間變異占主導地位。根生物量、莖生物量、葉生物量、地上生物量、總生物量、根分配比例、莖分配比例、葉分配比例、地上地下生物量比等9個生物量累積與分配指標經多重比較分析,均可以在P<0.05的顯著水平分為2組以上,具備家系間選育的差異基礎。所有指標中,生物量大小有關的指標及地上地下生物量比指標在家系間的變異較為豐富。但生物量大小有關的指標遺傳力較小且普遍低于中等水平,生物量分配比例有關的指標和地上地下生物量比遺傳力高于中等水平,將地上地下生物量比作為選育指標的遺傳增益最高。本研究發現生產力較高的苗木,地上地下生物量比處于較低水平。同時有研究表明,莖根比、高徑比等一些與苗木形態相關的參數,可以作為體現苗木活力水平的指標,如莖根比小的苗木活力水平高,造林后生長表現也較為良好[20]。因此,如以2 a生云南松苗木為材料開展早期選育時,宜選擇地上地下生物量比小、生物量較大的家系。
李蓮芳等[6]用不同苗齡云南松苗木進行造林效果的研究,認為苗齡較大、高徑生長量更大并解除“蹲苗”的苗木,有助于解決造林后一段時間高生長弱、競爭力低的問題,而且苗齡較大苗木的生長性狀分化程度也較高,提供更好的選育基礎。通過對比其他研究中云南松不同苗齡苗木的生物量情況,發現隨著苗木生長,最合理的生物量累積與分配指標可能有所變動。例如陳詩等[21]的研究中,育苗第1年由于苗木尚小,個體間地上部分對空間的競爭較小,苗木個體和家系將主要在爭奪水肥等生態限制因子的能力上產生更多的分化。而隨著苗齡和密度的增大,2、3 a生苗木為獲取生存資源,對地上部分的分配比例要比1 a生苗木更大,隨之將帶來對光資源競爭能力的強烈分化[22-23]。因此,建議繼續深入地摸清生物量各指標在時間上的遺傳變異規律,以便科學篩選不同苗齡苗木的選育指標,確定部分優良性狀的最佳考察時間[24]。
本研究中2 a生云南松苗根系生物量分配比例均低于蔡年輝等[25]、汪夢婷等[26]研究中2 a生苗木,而地上部分分配比例則較其他報道大。這可能與苗木種植密度、光水肥等管理措施有關。苗木的地上部分負責吸收太陽能進行光合作用,地下部分吸收水分和養分,兩個構件器官共同作用了植物與外界的交流和平衡,生物量的大小與比例變化,是植物形態對生態因子的可塑性具體體現[27]。本研究的播種密度為5 cm×5 cm,小于其他研究的5 cm×10 cm,因此,可能分配更多的生物量用于對地上空間的爭奪。
作為西南地區重要的陽性先鋒樹種,云南松的天然更新是森林演替和可持續管理的重要環節,本研究初步探討了苗木生物量在田間管理情況下家系間的變異規律。繼續挖掘自然狀態下云南松幼苗、幼樹的個體或群體生物量分配等生理生態規律,或能為該樹種的森林經營、苗木生產和遺傳改造提供更加豐富的基礎參考[28-32]。
猜你喜歡
天水行政學院學報(2022年4期)2022-11-18 09:02:36
艦船科學技術(2022年13期)2022-08-11 09:30:02
鐵道通信信號(2020年9期)2020-02-06 09:15:22
漢語世界(The World of Chinese)(2019年3期)2019-07-01 02:37:48
數學大王·趣味邏輯(2019年5期)2019-06-13 20:27:43
小學科學(學生版)(2019年5期)2019-05-21 01:00:18
中學生數理化·中考版(2018年10期)2018-12-07 00:44:52
經濟技術協作信息(2018年30期)2018-11-22 06:20:24
中央社會主義學院學報(2017年1期)2017-04-16 05:34:07
中國衛生(2014年12期)2014-11-12 13:12:40
