葛氏鱸塘鱧(Perccottus glenii)繁殖生物學及人工催產實驗
2021-02-25 02:40:28張玉勇何寶全曹頂臣白慶利何立川
水產學雜志 2021年6期
關鍵詞:生長
張玉勇,何寶全,曹頂臣,白慶利,何立川
(1.中國水產科學研究院黑龍江水產研究所,黑龍江 哈爾濱 150070;2.東北林業大學野生動物資源學院,黑龍江 哈爾濱 150040)
近些年,東北地區鯉(Cyprinus carpio)、鰱(Hypophthalmichthys molitrix)和草魚(Ctenopharyngodon idella)等大宗魚類塘口價格低迷,漁民養殖積極性深受影響,養殖規模不斷收縮。然而隨著人們消費觀念轉變和需求多樣性的增加,過去漁業生產中并未受到重視的小型魚類,越來越受到消費者的青睞,成為魚中上品,價格較高,市場前景看好。因此,小型特色經濟魚類的開發越來越成為水產養殖品種結構調整、提質增效的重要著力點。
葛氏鱸塘鱧(Perccottus glenii)屬鱸形目沙塘鱧科鱸塘鱧屬,俗稱“老頭魚”、“還陽魚”等,在我國是東北地區特有小型魚類[1,2]。隨著人們對其“肉質細嫩、味道鮮美、口感獨特、肌間刺少”等商品特性的認可,葛氏鱸塘鱧市場價格迅速上揚,體質量100 g左右的個體由原來的不足20 元/kg 升至40~60 元/kg。目前,受苗種制約,葛氏鱸塘鱧人工養殖規模小,市場銷售主要以江河、水庫和湖泊等自然水域捕撈為主,該魚苗種市場前景被普遍看好。葛氏鱸塘鱧人工繁殖的相關研究雖然已有開展并取得一定成績[3,4],但要實現規模化生產相關研究還需要深入。本研究在葛氏鱸塘鱧相關繁殖生物學研究的基礎上開展幾種藥物的催產實驗,以期為葛氏鱸塘鱧苗種規模化生產提供數據支撐。
1 材料與方法
1.1 材料
于2018 年5 月3 日、15 日、21 日,分三批次利用地籠在水產科學研究院黑龍江水產研究所渤海實驗基地場區蘆葦蕩捕獲野生葛氏鱸塘鱧,放入土池暫養,投喂小型雜魚雜蝦。
1.2 方法
生長及性比數據:采集葛氏鱸塘鱧體長和體質量數據,計算體長體質量關系。通過外形進行性比統計,對于外形難以區分的個體通過解剖的方式進行鑒別。并利用二項式檢驗來判斷雌雄分布是否符合概率為0.5 的二項分布。
體質量體長關系式:W=aLb;式中W 為體質量(g),L 為體長(cm)。
繁殖力測定:解剖葛氏鱸塘鱧雌魚50 尾,稱量體質量和卵巢重(0.1 g),再取0.1~1.0 g 的卵巢,稱重后計數其卵粒數,計算絕對繁殖力和相對繁殖力。
絕對懷卵量=單位體質量卵巢含有的卵粒數×卵巢重;
相對懷卵量=絕對懷卵量/體質量;
性成熟系數(GSI)=(性腺質量/空殼體質量)×100%。
人工催產:五月底,水溫達13℃時,解剖發現部分葛氏鱸塘鱧雄魚成熟較好,鏡檢發現其能夠產生少量游離精子,故無須注射催產藥物。解剖雌魚沒有發現卵巢進入Ⅴ期。將挑選的雌親魚隨機分成4組,每組30 尾,分別放入平列槽中(L 4 m×W 50 cm×H 0.4 m)。對照組魚注射生理鹽水,3 個實驗組分別注射促黃體素釋放激素類似物(LHRH-A2)、地毆酮(DOM)以及二者混合物。采用胸鰭基部單次腹腔注射,劑量等見表1。
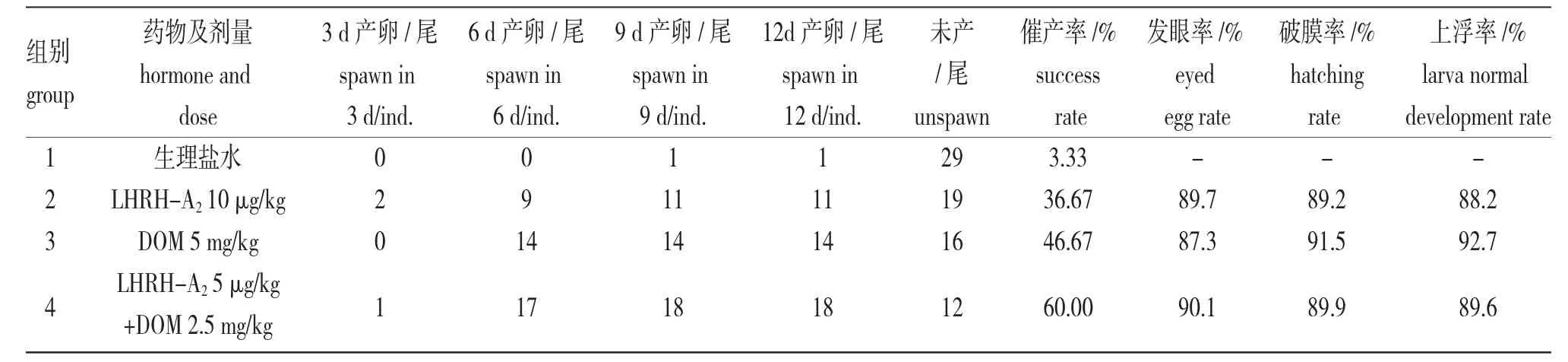
表1 葛氏鱸塘鱧人工催產效率Tab.1 Artificial propagation efficiency of Amur sleeper Perccottus glenii
催產后的雌魚分別放入已布好集卵巢的產卵池,并按照雌雄比2∶3 的比例配置挑選好的雄性親魚。放置親魚后水體交換量增大,流速0.3~0.5 m/s。從注射藥物算起,每3 d 檢查一次集卵巢是否有魚卵分布。產卵后將集卵巢移入室內平列槽中流水孵化。光照控制在500 lux 以下、流速0.1~0.2 m/s。催產率、發眼率、孵化率和上浮率的統計方式如下:
催產率(%)=產卵的雌魚數/催產的總雌魚數×100;
發眼率(%)=發眼卵數/產卵總數×100;
孵化率(%)=破膜仔魚數/發眼卵總數×100;
上浮率(%)=上浮仔魚數/ 破膜總仔魚數×100。
2 結果與分析
2.1 生長、性比
采捕到的葛氏鱸塘鱧群體體長變化在5.80~19.42 cm 之間,其中以全長12.00~15.00 cm 占優勢(33.8%)。雌性群體的平均體長為(14.61±9.24)cm,平均體質量為(55.8±18.5)g;雄性群體的平均體長為(15.23±6.46)cm,平均體質量為(61.8±11.9)g。雌、雄性群體體長體質量關系曲線分別為W=0.0326 L3.0595(R2=0.9860)和W=0.0274L3.0008(R2=0.9471)。
三批次群體性別鑒定結果顯示,葛氏鱸塘鱧雌雄性比分別為1∶1.091(n=69)、1∶0.970(n=132)、1∶0.938(n=93),三批次的整體雌雄性比為1∶0.986(n=294)。二項式檢驗結果顯示:葛氏鱸塘鱧繁殖群體雌雄性別分布服從平均概率0.5 的二項分布(P=1.00>0.05),即雌雄分布均勻。
2.2 繁殖力
用于測定繁殖力的樣本體長分布范圍為9.10~16.77 cm,年齡分布為2+齡~5+齡。由表2 可見,葛氏鱸塘鱧的GSI 區間為6.95%~13.3%,最高值出現在3+齡組;絕對懷卵量與年齡呈正比,從2+齡的2 140 粒增至5+齡的8 680 粒;相對懷卵量區間為86.5~103.1 粒/g,最高值仍出現在3+齡組。

表2 葛氏鱸塘鱧雌魚繁殖力測定Tab.2 Fecundity of female Amur sleeper(Perccottus glenii)
2.3 人工催產
葛氏鱸塘鱧剛產出的卵為球型,立即吸水膨脹后呈紡錘形,黏性較強,一端粘附在魚巢的避光面。卵的長軸:(4.5±0.7)mm,短軸:(1.2±0.4)mm。卵呈淡黃色,卵膜透明,用解剖鏡可以清晰觀測胚胎發育(圖1)。

圖1 葛氏鱸塘鱧受精卵、發眼卵及待破膜胚胎Fig.1 The fertilized eggs,eye-eggs and larva in egg membrane of Amur sleeper(Perccottus glenii)
由表1 可知,催產藥物對葛氏鱸塘鱧的催產效果明顯,藥物間差異很大,注射藥物3d 后開始發揮作用,6 d 或9 d 時效果最佳,但持續統計到12 d時,產卵親魚數量沒有新增。最終催產效率由高至低依次為:LHRH-A2+DOM>DOM>LHRH-A2,三組的催產率分別達到60.00%、46.67%和46.67%。藥物催產沒有明顯影響葛氏鱸塘鱧胚胎及苗種的早期發育。發眼率、孵化率和上浮率均維持在較高水平,其中,DOM 組發眼率最低,為87.3%,LHRH-A2+DOM組最高,為90.1%;LHRH-A2組的破膜率最低,為89.2%,DOM 組最高,為91.5%;LHRH-A2組上浮率最低,為88.2%,DOM 組最高,為92.7%。
3 討論
3.1 生長與性別
不同文獻記載的葛氏鱸塘鱧生長數據差異較大。在俄羅斯的Gusinoe 湖泊中捕獲的3+齡葛氏鱸塘鱧的標準體長為11.0 cm,而在Bodrog 河洪泛區,3+的標準體長僅有6.2 cm[5,6]。這種較大的生長差異可能是氣候、食物資源、密度等環境差異造成。事實上,同一物種存在較大生長變異是一種常見的現象,廣泛存在于多種魚類中[7-9]。本研究中,3+齡葛氏鱸塘鱧體長為11.5 cm,這主要得益于其生存環境。其棲息沼澤區域上游是鮭鱒魚養殖實驗區,殘餌和糞便等的排入導致沼澤地小型底棲生物和小型雜魚雜蝦等天然餌料生物極為豐富,導致較高的生長效率。
魚類生長具有性成熟前生長迅速,而性成熟后生長緩慢的特點[10],這主要是由于魚類性成熟后所消耗的餌料大部分不再用于體長的生長,而要保證性腺發育和成熟,以及越冬物質的積累[11]。但測量發現,葛氏鱸塘鱧在性成熟后仍會持續保持快速生長,這表明不能用性成熟來劃分該魚的生長階段。根據W=aLb中的b 值可以判別魚類的生長式型,即b=3 時為等速生長,b>或<3 時為非等速生長。等速生長是指魚類生長過程中體型和比例均保持不變[12]。魚類生長受環境變化的影響,使得b 值很少會等于3。在實際研究中,如果b 值接近3,就認為魚類的生長接近等速的[13]。本研究中不論雌性還是雄性b 值均接近3,所以葛氏鱸塘鱧是接近等速生長。這是由于標本采集的時間臨近葛氏鱸塘鱧的產卵期,2 齡時已經達到性成熟。此時魚體所獲能量主要用于性腺發育,為生殖做準備,所以表現出了較高的肥滿度指標。在多數魚類中,進入繁殖期的雌性生長指標會高于同齡的雄性,但葛氏鱸塘鱧并沒有發現雌性生長方面的二態性,這或許與該魚繁殖過程中的雄性護巢行為需要能量消耗相關,更大的雄性體型也有利于守護巢穴。但這種雌雄生長的差異是否會隨著年齡或棲息地等的不同而變化需要進一步的研究。
利用地籠三批次共捕獲葛氏鱸塘鱧294 尾,不論作為一個整體還是分批次統計分析,均顯示性比符合1∶1 分布。考慮到葛氏鱸塘鱧雄魚護巢的繁殖習性,1∶1 的雌雄比例分布將非常有利于葛氏鱸塘鱧群體的最大化擴張,最大程度上保證后代的延續。
外形對比和解剖觀察發現,雌雄異體的葛氏鱸塘鱧兩性差異主要表現在:在非繁殖期,雌魚體色比雄魚略淺,進入繁殖期,雌、雄魚體色都會加深,但雄魚體色明顯深于雌魚;繁殖期葛氏鱸塘鱧雄性生殖凸頂端較尖,雌性的較為鈍平(圖2);雌魚卵巢一對,成熟的卵巢呈淡黃色至金黃色,充滿腹腔。雄魚的精巢成對,性成熟的精巢為白色,前粗后細彎曲管狀,表面光滑。
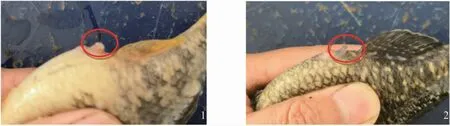
圖2 葛氏鱸塘鱧雌雄生殖凸形態Fig.2 Reproductive convex morphology of female and male Amur sleeper(Perccottus glenii)
3.2 繁殖與催產
資料顯示:葛氏鱸塘鱧已經成為近幾十年來東歐和中歐最具入侵性的魚類之一[14]。其分布范圍從原分布地俄羅斯東西伯利亞、中國東北部和朝鮮半島北部,迅速拓展到中東歐地區[15]。顯然,葛氏鱸塘鱧的生物學特征是促進其傳播的主要原因。而繁殖力或者繁殖習性更是一個品種最重要的擴張武器。本研究顯示,葛氏鱸塘鱧的絕對繁殖力在2 140~8 680 粒/尾,平均值為5 410 粒/尾。這一數據要高于金廣海(1 370~2 673 粒/ 尾)的報道[4],而低于Litvinov 報道[16]的數據。造成這種不同水域個體繁殖力差異的主要原因與這種魚的產卵策略密切相關。分布范圍比較廣的物種在生活史上往往會表現出很大的種內差異,這些差異是對當地環境條件的一種積極響應[17]。總體來看,葛氏鱸塘鱧繁殖力相對較低,考慮到能量分配,這應與該魚通過護巢等繁殖策略來提高繁殖的成功率密切相關。
葛氏鱸塘鱧繁殖期間水溫較低,而低溫條件下,性腺發育較慢,性腺同步性差,這對規模化養殖非常不利。為了提高親魚暫養成活率,促使其集中產卵,人工藥物催產繁殖成為必要手段。一般來說,溫度肯定會影響藥效的作用,因此試驗期間,一直耐心等到水溫穩定在12℃以上才開始進行藥物注射。實驗結果表明,LHRH-A2和DOM 這兩種常用的魚類催產藥物對葛氏鱸塘鱧催產效果較好,其中兩種藥物搭配使用效果最佳。同時各組別催產所獲受精卵在后期的發眼率、孵化率和上浮率均較高。考慮到低溫對藥效的不利影響,結合水溫調控來進行藥物催產效果應更佳。此外,考慮到葛氏鱸塘鱧的應激反應,建議選擇一次性藥物注射。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
