質(zhì)譜法分析烘焙對花生過敏原Ara h 1潛在致敏性的影響
2021-03-02 07:05:38周紅菲吳志華陳紅兵
食品科學(xué) 2021年3期
關(guān)鍵詞:結(jié)構(gòu)
周紅菲,吳志華,張 英,陳紅兵
(1.南昌大學(xué) 食品科學(xué)與技術(shù)國家重點(diǎn)實(shí)驗(yàn)室,江西 南昌 330047;2.南昌大學(xué)食品學(xué)院,江西 南昌 330031;3.南昌大學(xué)中德聯(lián)合研究院,江西 南昌 330047)
食物過敏被認(rèn)為是當(dāng)今社會(huì)一個(gè)重大的公共衛(wèi)生問題,受影響的兒童與成年人數(shù)量在快速增長[1]。花生作為“八大食物過敏原”之一[2],引發(fā)的常見臨床過敏癥狀有哮喘、皮膚瘙癢、過敏性鼻炎及口腔過敏綜合癥等。這是由于花生過敏原初次被機(jī)體攝入后,誘導(dǎo)特異性B細(xì)胞產(chǎn)生免疫球蛋白E(immunoglobulin E,IgE)抗體應(yīng)答[3],使機(jī)體處于致敏狀態(tài)。當(dāng)處于致敏狀態(tài)的機(jī)體再次接觸相同過敏原,會(huì)使IgE受體交聯(lián)引發(fā)致敏的肥大細(xì)胞或嗜堿性粒細(xì)胞活化和脫顆粒,釋放生物活性介質(zhì),導(dǎo)致毛細(xì)血管擴(kuò)張、血管通透性增加、平滑肌收縮等[4]。
目前已有17 種花生過敏原被世界衛(wèi)生組織的過敏原命名數(shù)據(jù)庫(http://www.allergen.org/)收錄,其中Ara h 1是花生蛋白中含量最高的過敏原,約占花生總蛋白含量的12%~16%[5]。
烘焙作為常見的花生制品加工方式之一,在改善食物感官品質(zhì)[6]的同時(shí),也會(huì)破壞蛋白質(zhì)的空間結(jié)構(gòu)[7-9]。溫度和熱處理時(shí)間都會(huì)影響蛋白質(zhì)結(jié)構(gòu)的變化程度[10]。有研究表明,當(dāng)溫度在70~80 ℃時(shí),蛋白質(zhì)的二級結(jié)構(gòu)會(huì)發(fā)生部分丟失,并伴隨著三級結(jié)構(gòu)可逆性地展開;當(dāng)溫度達(dá)到90 ℃時(shí),會(huì)有新的分子內(nèi)或分子間作用力形成;當(dāng)溫度接近100 ℃時(shí),蛋白質(zhì)會(huì)發(fā)生一定的聚集[10-11]。Hansen等[12]發(fā)現(xiàn)經(jīng)過熱處理后,花生過敏原的構(gòu)象性表位遭到了破壞,從而喪失了90%的免疫反應(yīng)性。雖然Rao Huan等[13]研究發(fā)現(xiàn),當(dāng)烘焙溫度達(dá)到130 ℃并持續(xù)5 min時(shí),可使花生蛋白的致敏性迅速降低,但多數(shù)研究表明花生蛋白致敏性會(huì)增強(qiáng),因?yàn)楹姹汉蟮鞍踪|(zhì)的二、三級結(jié)構(gòu)均遭到破壞,且蛋白具有更強(qiáng)的抗消化能力[14]。Moghaddam等[15]的研究同樣得出烘焙可以導(dǎo)致花生蛋白致敏性增強(qiáng)的結(jié)論,可能是由于烘焙過程中形成了新的過敏原表位。熱處理引起的空間結(jié)構(gòu)變化,在破壞原有的過敏原表位或生成新的過敏原表位的同時(shí)[16-17],也會(huì)暴露更多的酶切位點(diǎn),從而改變胃腸消化性[18-19]。
本研究通過基質(zhì)輔助激光解吸電離飛行時(shí)間質(zhì)譜(matrix assisted laser desorption ionization-time of flight mass spectrometry,MALDI-TOF-MS)技術(shù)檢測Ara h 1酶解肽段,結(jié)合蛋白空間結(jié)構(gòu)模型,分析烘焙對花生過敏原Ara h 1的結(jié)構(gòu)以及潛在致敏性的影響,以期為揭示加工前后蛋白構(gòu)效關(guān)系提供新的思路。
1 材料與方法
1.1 材料與試劑
鮮花生購自江西省南昌市菜市場,將新鮮花生進(jìn)行帶殼烘焙,條件為170 ℃、35 min。之后將花生去殼、去紅衣并研磨,利用丙酮脫脂,得到花生脫脂粉。
脲、硫脲、3-[3-(膽酰胺丙基)二甲氨基]丙磺酸內(nèi)鹽(3-[(3-cholamidopropyl)dimethylammonio]propanesulfonate,CHAPS)、二硫蘇糖醇(DLdithiothreitol,DTT)、碘乙酰胺(iodoacetamide,IAM)、碳酸氫銨(NH4HCO3)、乙腈(acetonitrile,ACN)(色譜級,純度不低于99.9%);三氟乙酸(trifluoroacetic acid,TFA)(色譜級,純度不低于99%)美國Sigma公司;Trypsin(測序級) 德國Promega公司。
1.2 儀器與設(shè)備
PB-10型pH計(jì) 德國Sartorius公司;真空離心濃縮儀 美國Labconco公司;BR680型酶標(biāo)儀、迷你蛋白電泳儀 美國Bio-Rad公司;SQ-GS800光密度掃描儀北京宇艾奇科技有限公司;MALDI-TOF-MS儀 日本Shimadzu公司。
1.3 方法
1.3.1 蛋白質(zhì)樣品的提取
提取液:7 mol/L脲、2 mol/L硫脲、2 g/100 mL CHAPS、1 g/100 mL DTT,用超純水定容至100 mL,調(diào)節(jié)pH值至8.8。
花生蛋白的提取:取0.3 g花生脫脂粉,加入15 mL提取液,60 ℃超聲波(160 W)提取15 min。之后在20 ℃、9 000 r/min條件下離心20 min,棄去沉淀,用0.45 μm濾膜對上清液進(jìn)行過濾。最終將蛋白溶液凍存于-20 ℃?zhèn)溆谩?/p>
1.3.2 十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳定性分析
采取不連續(xù)體系的十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)分析花生脫脂粉提取液中蛋白組分。將蛋白提取液中花生蛋白質(zhì)量濃度稀釋到1 mg/mL,上樣量為15 μL。Marker上樣量為7 μL,選用質(zhì)量分?jǐn)?shù)為12%分離膠、4%濃縮膠。設(shè)定電泳條件分別為恒流6 mA和12 mA,時(shí)間分別為30 min和90 min。電泳結(jié)束后,進(jìn)行剝膠、染色、脫色等工序。采用SQ-GS800光密度掃描儀,對蛋白電泳凝膠進(jìn)行圖象采集和定性分析。
1.3.3 蛋白膠內(nèi)酶解
從凝膠中切下目的條帶,用超純水沖洗多次。向膠塊中加入脫色液(25 mmol/L NH4HCO3-50% ACN(NH4HCO3與50% ACN體積比為1∶1)),漩渦振蕩10 min,直至膠塊無色透明,吸去脫色液。
在脫色后的膠塊中,加入少量100% ACN,于真空濃縮儀中56 ℃干燥約30 min,使膠塊完全脫水。再利用100 mmol/L DTT和200 mmol/L IAM對蛋白膠塊分別進(jìn)行還原和烷基化。加入少量25 mmol/L NH4HCO3溶液,漩渦振蕩,棄去上清液,再次干燥膠塊。干燥好的膠塊上加入Trypsin酶液(Trypsin與蛋白質(zhì)量比為1∶30~1∶50),4 ℃放置1 h使酶液完全被吸收,另補(bǔ)加10 μL NH4HCO3溶液保濕,37 ℃孵育15 h。
收集肽段混合物。在膠塊內(nèi)加入質(zhì)量分?jǐn)?shù)10% TFA,收集上清液;再向膠塊中加入肽段抽提液(含體積分?jǐn)?shù)60% ACN、質(zhì)量分?jǐn)?shù)0.1% TFA),超聲15 min,收集上清液;合并兩次上清液,置于4 ℃?zhèn)溆谩?/p>
1.3.4 質(zhì)譜分析
先點(diǎn)0.5 μL樣品于MALDI靶板上,再取0.5 μL α-氰基-4-羥基肉桂酸基質(zhì)溶液覆蓋其上,待樣品與基質(zhì)形成的共結(jié)晶薄膜干燥后進(jìn)行檢測。選擇反射模式,每孔采集140 次,MS質(zhì)量掃描范圍:m/z 800~3 000。采用TOF-mix標(biāo)準(zhǔn)品進(jìn)行外標(biāo)法校準(zhǔn),校準(zhǔn)后進(jìn)行質(zhì)譜數(shù)據(jù)采集。
通過Mascot網(wǎng)頁進(jìn)行在線肽指紋圖譜匹配,搜庫參數(shù)設(shè)定:選擇UniProt數(shù)據(jù)庫,種屬為Other green plants,使用酶為Trypsin,允許2 個(gè)漏切位點(diǎn)。質(zhì)譜數(shù)據(jù)及圖譜處理:MALDI-TOF-MS產(chǎn)生的原始數(shù)據(jù)及圖譜均由Biotech MALDI-MS軟件導(dǎo)出,并結(jié)合Trypsin理論酶解肽段分子質(zhì)量確定肽段序列。
2 結(jié)果與分析
2.1 SDS-PAGE條帶分析結(jié)果
通過SDS-PAGE對鮮花生和烘焙花生的蛋白提取液分別進(jìn)行了組分分析,如圖1所示。主要過敏原蛋白Ara h 1位于約64 kDa處,即紅色方框標(biāo)記的條帶。與鮮花生相比,烘焙花生樣品的各過敏原條帶均不同程度變淺,Ara h 1單體含量明顯降低,且在大于180 kDa處出現(xiàn)了高聚物,可以看出烘焙明顯導(dǎo)致了蛋白質(zhì)的聚集。烘焙花生樣品的泳道中也出現(xiàn)較多的彌散狀條帶,說明烘焙也可能造成蛋白質(zhì)分子的降解。
熱加工會(huì)導(dǎo)致花生蛋白的二級和三級結(jié)構(gòu)的改變,表現(xiàn)出部分結(jié)構(gòu)舒展,甚至還有分子聚集現(xiàn)象[11],從而影響蛋白質(zhì)的可溶性。Schmitt等[6]的研究也得到了相似結(jié)果。本次提取蛋白采用的是高離液序列鹽溶液,可以盡可能全面地提取出花生過敏原[20-21]。該提取液中含有的變性劑、還原劑等可以裂解已經(jīng)聚集的蛋白,恢復(fù)蛋白質(zhì)的可溶性,而烘焙花生的Ara h 1含量仍然顯著降低,這可能是由于烘焙的溫度較高、時(shí)間也較長,導(dǎo)致生成物聚集的程度較高。有研究表明,Ara h 1可以通過美拉德反應(yīng)形成共價(jià)交聯(lián)的六聚體[22],且烘焙加工后Ara h 1也會(huì)與Ara h 6形成交聯(lián)[23],產(chǎn)生高聚物。
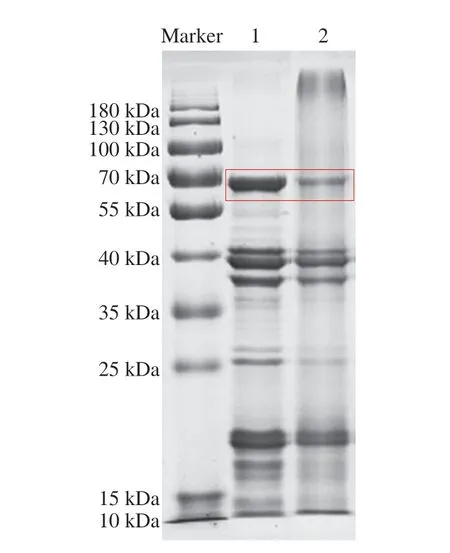
圖1 鮮花生與烘焙花生蛋白的SDS-PAGE圖Fig.1 SDS-PAGE profile of proteins in raw and roasted peanuts
2.2 MALDI-TOF-MS肽段分析結(jié)果
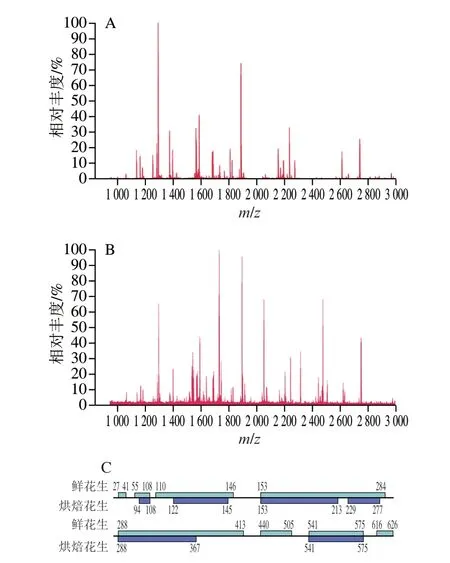
圖2 鮮花生與烘焙花生蛋白的質(zhì)譜圖及氨基酸序列覆蓋區(qū)域Fig.2 Mass spectra and amino acid sequences of proteins in raw and roasted peanuts
對圖1中紅色方框標(biāo)記的泳道1和泳道2蛋白條帶分別進(jìn)行質(zhì)譜鑒定,并搜索Uniprot數(shù)據(jù)庫,均可成功匹配蛋白Ara h 1,蛋白序列登錄號為P43238。鮮花生和烘焙花生樣品蛋白打分分別為120 分和64 分,可能是烘焙加工造成的酶切肽段變化,致使結(jié)果與肽指紋圖譜匹配度下降。如圖2A、B所示,花生經(jīng)烘焙后,MALDI-TOF-MS一級圖譜中肽段分布情況發(fā)生了明顯變化。鮮花生和烘焙花生樣品中共檢測到Ara h 1的71 條肽段,其中鮮花生中檢測到70 條肽段,如圖2C所示,氨基酸覆蓋率達(dá)到79.2%。烘焙加工后,僅檢測到31 條肽段,較鮮花生新增1 條肽段,缺失40 條肽段,氨基酸覆蓋率為43.9%。這可能是由于烘焙使得花生中水分大幅降低[24],蛋白質(zhì)結(jié)構(gòu)變得緊密,掩蓋了部分酶切位點(diǎn),致使很多原先鮮花生中可檢測到的肽段在烘焙樣品中出現(xiàn)了缺失。
2.3 烘焙對蛋白質(zhì)空間結(jié)構(gòu)的影響

表1 鮮花生與烘焙花生肽段差異比較Table 1 Peptide profiles of raw and roasted peanuts

續(xù)表1
質(zhì)譜檢出的肽段及其位置差異在一定程度上可以反映花生烘焙前后蛋白質(zhì)高級結(jié)構(gòu)的變化。如表1所示,在鮮花生和烘焙花生樣品中均檢測到第67~70號肽段,這4 條肽段覆蓋了541IFLAGDKDNVIDQIEKQAKDLAFPGS GEQVEKLIK575氨基酸序列。其胰蛋白酶理論酶切位點(diǎn)有5 個(gè),經(jīng)質(zhì)譜只檢測到4 個(gè),烘焙花生蛋白保持著和鮮花生相同的酶解方式,沒有酶切位點(diǎn)變化。說明這一區(qū)域結(jié)構(gòu)較為穩(wěn)定,烘焙對酶切位點(diǎn)的影響較小。
第56~66號肽段覆蓋了兩段多肽區(qū)域,分別為371Glu-Lys413和440Glu-Lys505。這些肽段在鮮花生均有檢出,在烘焙后的花生中則均未檢出。可能是由于烘焙使得Ara h 1結(jié)構(gòu)趨于緊實(shí)[24],371Glu-Lys413和440Glu-Lys505這兩片區(qū)域的酶切位點(diǎn)集中被遮擋,形成較大分子質(zhì)量的多肽,可能已經(jīng)超出設(shè)定的分子質(zhì)量檢測范圍。出現(xiàn)與此情況類似的區(qū)域還有616Gly-Asn626、27Ser-Arg41和55Gln-Arg91等。
鮮花生中檢出了第28號肽段193IIe-Arg202和第30號肽段195Val-Arg204,未檢出第29號肽段193IIe-Arg204;而烘焙花生中檢出193IIe-Arg204和195Val-Arg204,未檢出193IIe-Arg202,說明烘焙改變了蛋白的結(jié)構(gòu),掩蔽了202Arg與203Ser之間的酶切位點(diǎn),造成漏切,產(chǎn)生了分子質(zhì)量較大的酶解肽段。
肽段199FDQRSR204(第31號肽段)在烘焙后的樣品中未被檢出,則可能是烘焙暴露了更多酶切位點(diǎn),產(chǎn)生分子質(zhì)量更小的肽段,如199FDQR202和203SR204等,這些肽段分子質(zhì)量也在檢測范圍之外,因而未能檢出。

圖3 質(zhì)譜檢測發(fā)現(xiàn)的Ara h 1胰蛋白酶酶切位點(diǎn)Fig.3 Trypsin cleavage sites of Ara h 1 detected by mass spectrometry
利用Swiss-Model蛋白質(zhì)結(jié)構(gòu)同源建模服務(wù)器,在線構(gòu)建Ara h 1的空間結(jié)構(gòu)模型(結(jié)構(gòu)模型僅展示171Arg-Arg586氨基酸序列部分)。Ara h 1是屬于豌豆球蛋白家族的一類貯藏蛋白[25-26],是由3 個(gè)對稱的單體非共價(jià)結(jié)合形成的三聚體結(jié)構(gòu)[18,27]。圖3為Ara h 1單體的空間結(jié)構(gòu),并分別標(biāo)記出了鮮花生與烘焙花生中被胰蛋白酶識別出的酶切位點(diǎn)(C端)。可以發(fā)現(xiàn),靠近Ara h 1單體C末端的α-螺旋上,酶切位點(diǎn)在烘焙前后未發(fā)生變化,說明這一區(qū)域的酶切位點(diǎn)沒有被掩蔽。主要是因?yàn)锳ra h 1單體末端α-螺旋上的疏水性殘基有助于維持結(jié)構(gòu)穩(wěn)定[25],烘焙后這一區(qū)域結(jié)構(gòu)變化不大,且Ara h 1高度穩(wěn)定的三聚體結(jié)構(gòu)的形成也正是由疏水相互作用介導(dǎo)。烘焙花生中漏切的酶切位點(diǎn)主要出現(xiàn)在α-螺旋和β-折疊結(jié)構(gòu)的末端和無規(guī)卷曲中。烘焙過程中,α-螺旋和β-折疊結(jié)構(gòu)可能遭到了一定程度的破壞[28],尤其是α-螺旋結(jié)構(gòu)發(fā)生丟失[29],無規(guī)卷曲含量增多,可能使酶切位點(diǎn)被遮擋。烘焙后酶切位點(diǎn)被遮擋,也會(huì)對蛋白的消化性造成影響[30]。
2.4 蛋白結(jié)構(gòu)變化對過敏原表位的影響
據(jù)報(bào)道,Ara h 1已知的過敏原線性表位有21 個(gè)[31-33]。質(zhì)譜檢出鮮花生的肽段涉及18 個(gè)過敏原線性表位,而烘焙花生中僅檢出其中13 個(gè)。經(jīng)過質(zhì)譜前處理,其中,鮮花生中檢出16 個(gè)過敏原線性表位被胰蛋白酶酶解,烘焙花生中僅發(fā)現(xiàn)12 個(gè)。由表1可知,在烘焙花生中發(fā)生缺失的過敏原表位肽,主要分布在371Glu-Lys413和440Glu-Lys505區(qū)域,由于酶切位點(diǎn)被掩蔽,會(huì)形成較大分子質(zhì)量的多肽。說明在酶解作用下,鮮花生Ara h 1中多達(dá)88%的過敏原線性表位均遭到了破壞,烘焙后Ara h 1的部分酶切位點(diǎn)被掩蔽,過敏原線性表位的被破壞程度下降。由圖4中的Ara h 1結(jié)構(gòu)模型可以看出,Ara h 1單體結(jié)構(gòu)分為了4 個(gè)區(qū)域[25],包括一端的α-螺旋結(jié)構(gòu)、兩個(gè)對稱的反平行β-折疊結(jié)構(gòu)、另一端的α-螺旋結(jié)構(gòu)。大部分的Ara h 1的過敏原線性表位均集中分布在Ara h 1單體與單體接觸區(qū)域。被破壞的過敏原表位與檢出的酶切位點(diǎn)所在區(qū)域發(fā)生重疊。在烘焙之前,過敏原線性表位受到酶解的破壞較為嚴(yán)重,僅小部分過敏原線性表位保存完整性,有10 個(gè)過敏原線性表位被破壞。在烘焙后,有7 個(gè)過敏原線性表位被破壞,少于鮮花生樣品。說明烘焙后,過敏原線性表位被胰蛋白酶酶解破壞的程度降低。在酶解作用下,相比于烘焙前Ara h 1被破壞過敏原線性表位的分布,烘焙后主要集中在Ara h 1單體兩端的α-螺旋結(jié)構(gòu)上,較少破壞兩個(gè)反平行β-折疊區(qū)域的過敏原線性表位。說明烘焙可能造成了反平行β-折疊區(qū)域的結(jié)構(gòu)緊縮,遮擋了相應(yīng)的酶切位點(diǎn),造成消化對過敏原線性表位的破壞程度下降[30],使得Ara h 1潛在致敏性增強(qiáng)。
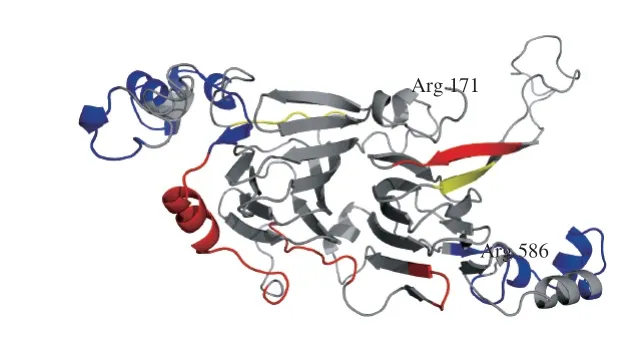
圖4 質(zhì)譜檢測發(fā)現(xiàn)的Ara h 1過敏原線性表位Fig.4 Linear IgE-binding epitopes of Ara h 1 detected by mass spectrometry
Beyer等[34]研究也發(fā)現(xiàn),較高溫度的干熱烘焙可以提高花生3 種主要過敏原的致敏性。烘焙過程中,可能會(huì)產(chǎn)生眾多如伸展、聚集和交聯(lián)等結(jié)構(gòu)性的改變[35],花生組分之間也會(huì)發(fā)生反應(yīng)(如常見的美拉德反應(yīng)),對花生蛋白產(chǎn)生修飾作用。有研究表明,烘焙后花生全蛋白提取物[14]和純化的Ara h 1致敏性均會(huì)增強(qiáng)[22],可能是由于美拉德反應(yīng)對蛋白的修飾增強(qiáng)了過敏原表位耐胃腸消化的能力。另外,有研究也發(fā)現(xiàn)熱加工會(huì)改變蛋白質(zhì)二級與三級結(jié)構(gòu),不僅會(huì)影響對過敏原線性表位的破壞程度,也會(huì)對過敏原構(gòu)象型表位產(chǎn)生掩蓋、暴露或破壞等影響[35],從而影響潛在致敏性。因此,烘焙導(dǎo)致的Ara h 1結(jié)構(gòu)變化,使得部分過敏原線性表位未被酶解破壞,可能是烘焙后花生潛在致敏性增強(qiáng)的重要原因之一。
3 結(jié) 論
本研究分析了烘焙對花生主要過敏原Ara h 1結(jié)構(gòu)的影響,結(jié)果發(fā)現(xiàn)烘焙加工后,Ara h 1會(huì)發(fā)生聚集和降解。結(jié)合質(zhì)譜和空間模型構(gòu)象,對加工后Ara h 1的高級結(jié)構(gòu)進(jìn)行了分析,發(fā)現(xiàn)烘焙也可以改變Ara h 1的高級結(jié)構(gòu)。高級結(jié)構(gòu)的變化掩蓋了部分酶切位點(diǎn),從而減輕了酶解對過敏原線性表位的破壞程度,這可能導(dǎo)致烘焙加工后Ara h 1致敏性強(qiáng)于鮮花生。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學(xué)評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學(xué)生數(shù)理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現(xiàn)代企業(yè)(2015年9期)2015-02-28 18:56:50
