雅安周公山不同發育階段峨眉含笑的枝葉性狀
2021-03-04 02:01:08趙安玖陳智超呂佳瑤李曜汐
浙江農林大學學報 2021年1期
彭 希,趙安玖,陳智超,呂佳瑤,李曜汐
(四川農業大學 林學院,四川 成都 611130)
植物功能性狀是指對植物個體生存與發展有著一定影響的植物特征,與植物自身的生存策略密切相關[1?2]。植物的生物學特性間接反映植物各功能性狀間的權衡方式,影響著植物在群落中的生存與發展。在植物的各種器官間,小枝和葉片是植物器官分支系統中最敏感的部分,在植物的不同發育階段,小枝和葉性狀是植物與環境相互作用的結果[3]。植物如何通過調整小枝內各構件之間的生長關系,來適應不斷變化的環境,是研究植物生態策略的重要內容[4]。作為植物光合作用重要的器官,葉片能夠將光能轉化為自身發育所需要的物質,其大小直接影響著植物個體的發育模式以及對光能的捕獲和轉化能力[5]。小枝是植物葉片直接著生的器官,它能夠傳輸莖運輸過來的養分及葉光合作用產生的同化物質,與植物在空間上的開展情況密切相關,并決定著葉片的投資方式[6]。小枝與葉片之間的關系是植物個體在不同發育階段的生態策略體現[7]。不同物種對環境的適應情況不同,因此生存策略方式也不同,即快速高效策略和慢速高質量策略[8?10]。物種內存在不同發育階段的植物個體,它們在環境中獲得的資源情況不同。為了提高對環境的適應性,植物枝葉可能會在不同發育階段有著不同的投資方式。植物小枝與葉片對環境變化的敏感性強,植物個體在不同發育階段的生態策略容易在其性狀上表現出來[10]。種群空間格局指種群個體在空間上的分布情況,是與外界環境相互作用的結果,在一定程度上反映種群的生態策略方式[11]。目前,關于種群空間格局的研究主要集中在不同物種間、物種內不同發育階段等方面[11?12],在植物性狀間空間格局的分析較少。地統計學作為研究植物空間格局的重要方法,可以真實反映植物的空間變化[13]。SIEFERT[14]采用半方差變異函數對農田植物葉功能性狀的空間格局進行了研究,發現葉功能性狀具有中等空間自相關性,植物功能性狀在空間上是連續變化的,進而能夠預測植物性狀的空間變化情況。探究植物枝葉性狀的空間變化有助于理解植物對環境的適應策略。峨眉含笑Michelia wilsonii是多年生常綠喬木,主要生長于氣候濕潤,海拔600~2 000 m的常綠闊葉林中。雖然峨眉含笑的結實量較多,但其自然更新困難,分布區域較小,現已瀕臨滅絕,為國家Ⅱ級保護樹種。目前,對于峨眉含笑的研究主要集中在群落結構、凋落物特征等方面[15?16],對其生態策略方式還不清楚。因此,本研究以四川省雅安周公山峨眉含笑優勢闊葉林為基礎,選取種群內不同發育階段個體為研究對象,分析枝葉性狀在空間上的自相關性,以期為了解峨眉含笑枝葉性狀之間的資源權衡方式和生態策略方式提供依據。
1 研究地區與研究方法
1.1 研究區概況
研究區位于四川省雅安周公山國家森林公園內 (29°58′09″N,103°02′58″E)。該區屬亞熱帶季風性濕潤氣候,降水量大,年均降水量為1 774.3 mm;濕度大,年平均相對濕度79%;氣溫較低,年平均氣溫14.0 ℃,海拔900~1 180 m;土壤類型以山地黃壤為主。研究區內植被豐富,以常綠闊葉林為主,除峨眉含笑優勢樹種外,伴生有杉木Cunninghamia lanceolata、華中櫻桃Cerasus conradinae、栗Castanea mollissima、日本杜英Elaeocarpus japonicus、刺楸Kalopanax septemlobus、燈臺Bothrocaryum controversum、南酸棗Choerospondias axillaris等喬木樹種,林分郁閉度為0.8~0.9;林下主要有水竹Phyllostachys heteroclada、姬蕨Hypolepis punctata及一些禾草等植物,蓋度達80%以上[15]。
1.2 樣地設置
在峨眉含笑種群分布較多的地區,建立1塊200 m×200 m的固定樣地,并將樣地劃分為100個20 m×20 m的樣方。逐一對樣方內所有胸徑≥5 cm的峨眉含笑個體進行掛牌登記,并記錄胸徑、樹高等信息,以便進行后期采樣工作。
1.3 枝葉采集
于2018年7月采集樣地內峨眉含笑個體的葉片和枝條。根據研究區域峨眉含笑種群的年齡結構及數量動態特征[16],將其劃分為小樹(胸徑5~15 cm)、中樹(胸徑15~25 cm)和大樹(胸徑>25 cm) 3個階段。定義當年生小枝為1年生小枝,沿著1年生小枝依次向內為2年生、3年生小枝。為了保證每株樹采集的1、2年生枝條和葉來自同一個樹枝,本研究在樹冠南部枝條分叉處直徑約1 cm粗的地方剪取1個樹枝,從樹枝上采集6個能明顯區分1、2年生的小枝。采集小枝上成熟的當年生葉片15片,并將1、2年生小枝中均勻生長的部分截斷,將其保存在自封袋中,做好標記帶回實驗室進行后續指標測定。共采集20株小樹、59株中樹、34株大樹的枝葉。
1.4 指標測定
葉片鮮質量使用電子天平(精度0.000 1 g)稱量;將稱量過后的葉片放入烘箱中,在105 ℃下烘20 min進行殺青,然后在80 ℃下烘48 h,用電子天平稱其干質量(精度0.000 1 g)。用葉面積(LI-3100C,LI COR,美國)掃描儀測定葉片后,在PS6中計算葉片面積;用游標卡尺測量葉片厚度(精度0.01 mm)。葉干鮮比=葉片干質量(g)/葉片鮮質量(g);比葉面積=葉片面積(cm2)/葉片干質量(g)。將所有小枝浸入去離子水中8~12 h,待小枝達到飽和狀態后將其取出,用排水法測量小枝體積。將測量完體積的小枝,用吸水紙吸去表面水分后用電子天平稱量(精度0.000 1 g),獲得小枝鮮質量,然后將其放入烘箱中80 ℃烘48 h,用電子天平稱量(精度0.000 1 g),獲得小枝干質量。小枝干鮮比=小枝干質量(g)/小枝鮮質量(g);小枝密度=小枝干質量(g)/小枝體積(cm3)。
1.5 數據處理
采用單因素方差分析對峨眉含笑種群不同發育階段枝葉性狀進行差異分析;采用標準化主軸分析計算枝葉性狀間的相關性。為了使枝葉性狀呈正態分布,對其進行了對數轉換(以10為底數)。數據分析在R語言的smatR包中進行。
通過主成分分析獲得每株樹木第1軸枝葉性狀的得分值,然后采用半方差函數法對峨眉含笑種群不同發育階段枝葉性狀主成分得分進行空間格局分析[17?18]。其計算公式為:
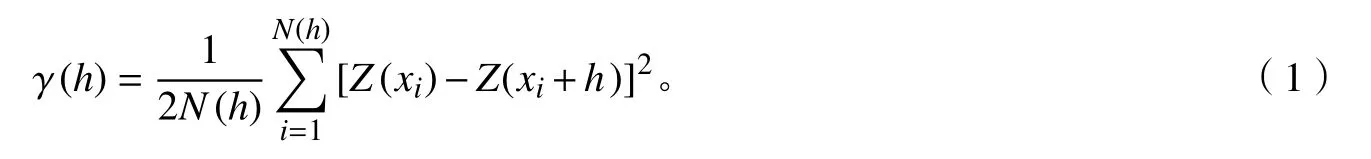
式(1)中:γ(h)為林木個體間距為h時的半方差函數值;N(h)為林木個體間距為h時的樣本對數;Z(xi)為林木個體i在位置點xi處的實測值;Z(xi+h)為林木個體在位置點xi+h處的實測值。通過半方差值進行模型擬合,繪制半方差函數圖。
本研究選取4種模型進行擬合(球狀模型、指數模型、高斯模型和線性模型),選取模型決定系數(R2)最大和殘差最小的模型。通過最優模型獲得基臺值(C0+C)、偏基臺值(C)、塊金值(C0)和變程(A),求得結構方差比[C/(C0+C)]來表示研究區變量的最大變異程度。一般認為結構方差>75%時,空間自相關性強;結構方差位于25%~75%時,空間自相關性中等,結構方差<25%時,空間自相關弱,此時不宜采用克里格(Kringing)插值進行變量預測。
2 結果與分析
2.1 峨眉含笑不同發育階段枝葉性狀特征
不同發育階段葉鮮質量、葉干質量和2年生小枝密度差異顯著(P<0.05),大樹與中樹、小樹之間葉面積、比葉面積、1年生小枝密度差異顯著(P<0.05),中樹和小樹之間葉干鮮比和2年生小枝干鮮比差異顯著(P<0.05),其余性狀差異不顯著(P>0.05)(表1)。葉鮮質量、葉干質量、葉面積、葉厚、1年生小枝干鮮比、1年生小枝密度隨著林木胸徑的增加呈顯著增加(P<0.05),比葉面積隨著林木胸徑的增加顯著降低(P<0.05)(表2)。主成分分析前2軸的解釋總方差為56.99%,其中第1軸的解釋方差為39.95%,第2軸的解釋方差為17.04%(圖1)。對枝葉性狀進行排序,葉性狀與第1排序軸的相關性較高,枝性狀與第2排序軸相關性較好。在第1排序軸中,除比葉面積外,從左到右,枝葉性狀增加,幼樹有著較高的比葉面積值,而大樹除比葉面積外,其他枝葉性狀較高;在第2排序軸中,從下到上,枝性狀逐漸增加,葉性狀逐漸減小,幼樹有著較低的枝葉性狀。這意味著更高發育階段的個體有著更高的枝葉性狀。
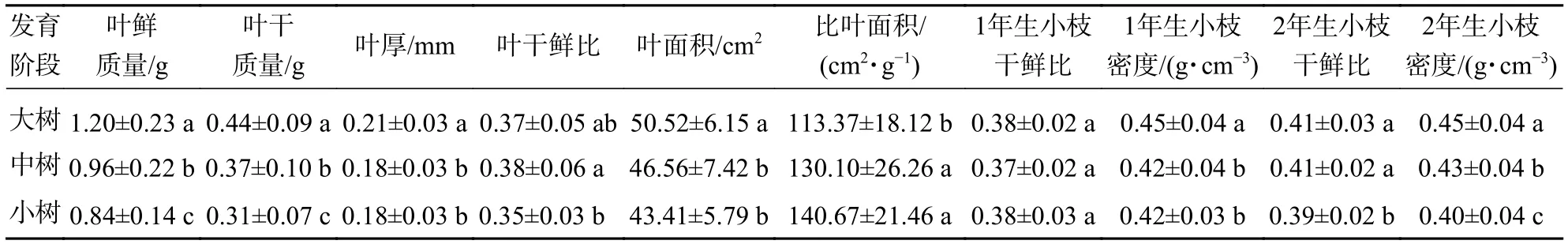
表1 不同發育階段枝葉性狀特征Table 1 Traits of twig and leaf in different development stages

表2 枝葉性狀與胸徑之間的相關系數Table 2 Correlation coefficient between twig, leaf traits and DBH
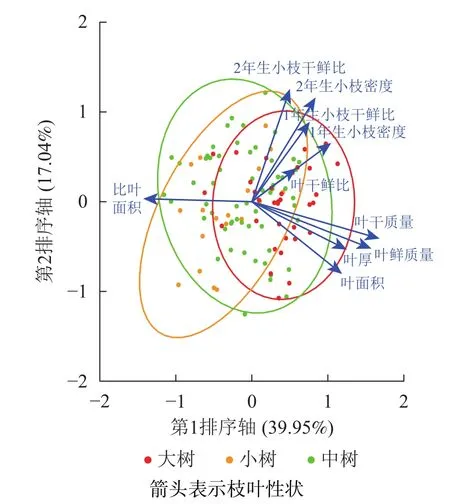
圖1 不同發育階段枝葉性狀主成分排序圖Figure 1 PCA ordination of twig and leaf traits at different developmental stages
2.2 不同發育階段枝葉性狀相互關系
標準化主軸分析發現(表3):葉性狀之間及枝性狀之間有著較強的相關關系,葉性狀與莖性狀之間相關性較弱,僅葉鮮質量和葉干質量與小枝性狀具有相關性。主成分分析(PCA)也發現 (圖1):葉性狀與小枝性狀存在正交關系。從表4~6可以看出:不同發育階段葉性狀間相關顯著(P<0.05),其中,大樹和中樹階段葉性狀間相關性均比小樹階的段相關性強;不同發育階段枝性狀之間相關性從大到小依次為小樹、中樹、大樹。枝葉性狀之間相關性,除了小樹階段葉干質量和1年生小枝密度,大樹階段葉鮮質量和1年生小枝干鮮比、1年生小枝密度,中、小樹階段比葉面積和1年生小枝密度,中樹階段葉厚和1年生小枝密度顯著相關外(P<0.05),其余均不顯著(P>0.05)。
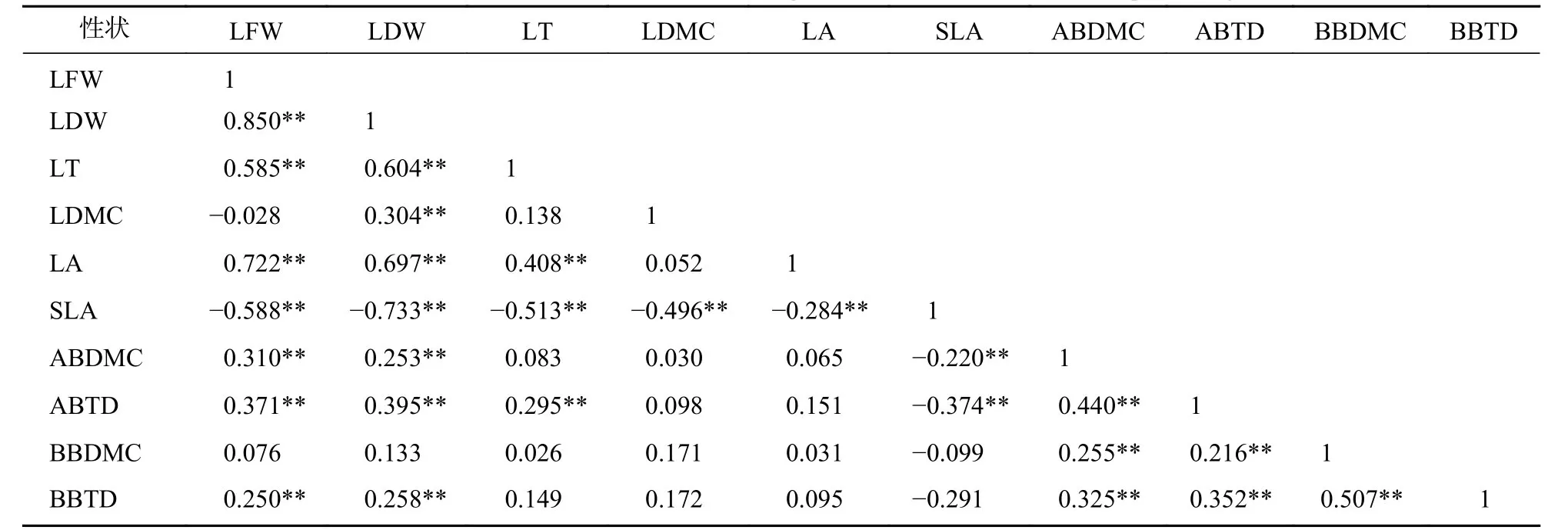
表3 整個發育階段枝葉性狀之間的相關系數Table 3 Correlation coefficient between the twig and leaf traits in the whole development stage
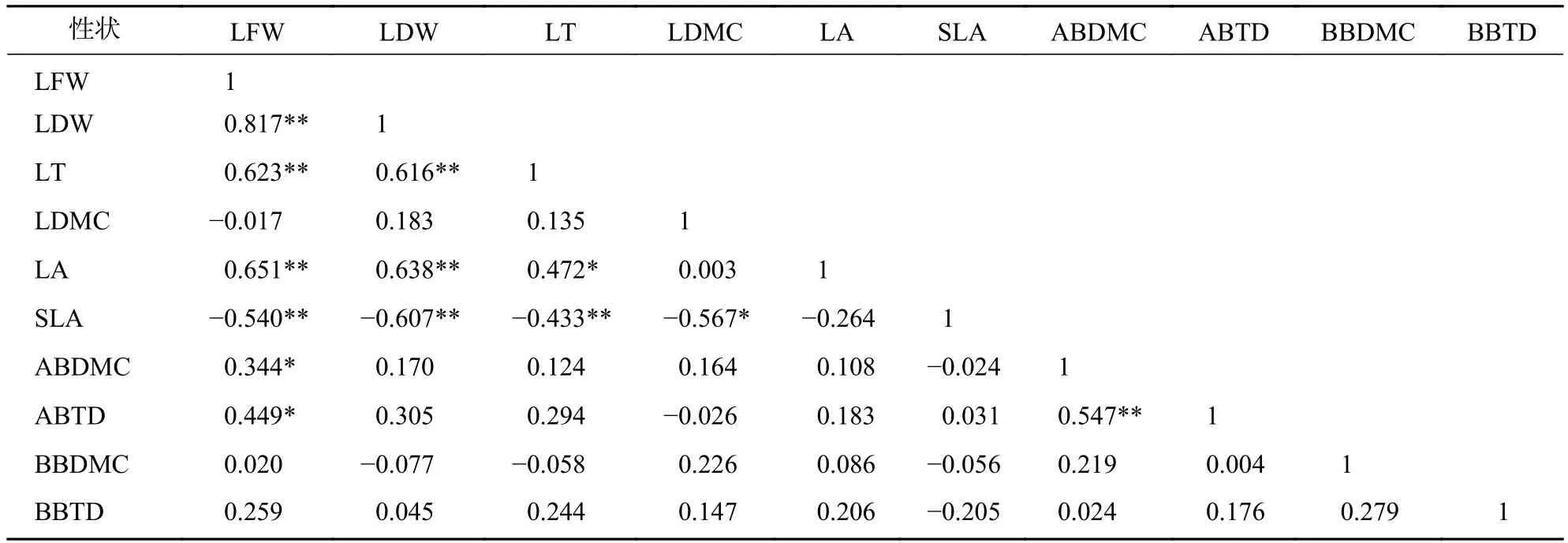
表4 大樹階段枝葉性狀之間的相關系數Table 4 Correlation coefficient between the twig and leaf traits in big tree stage
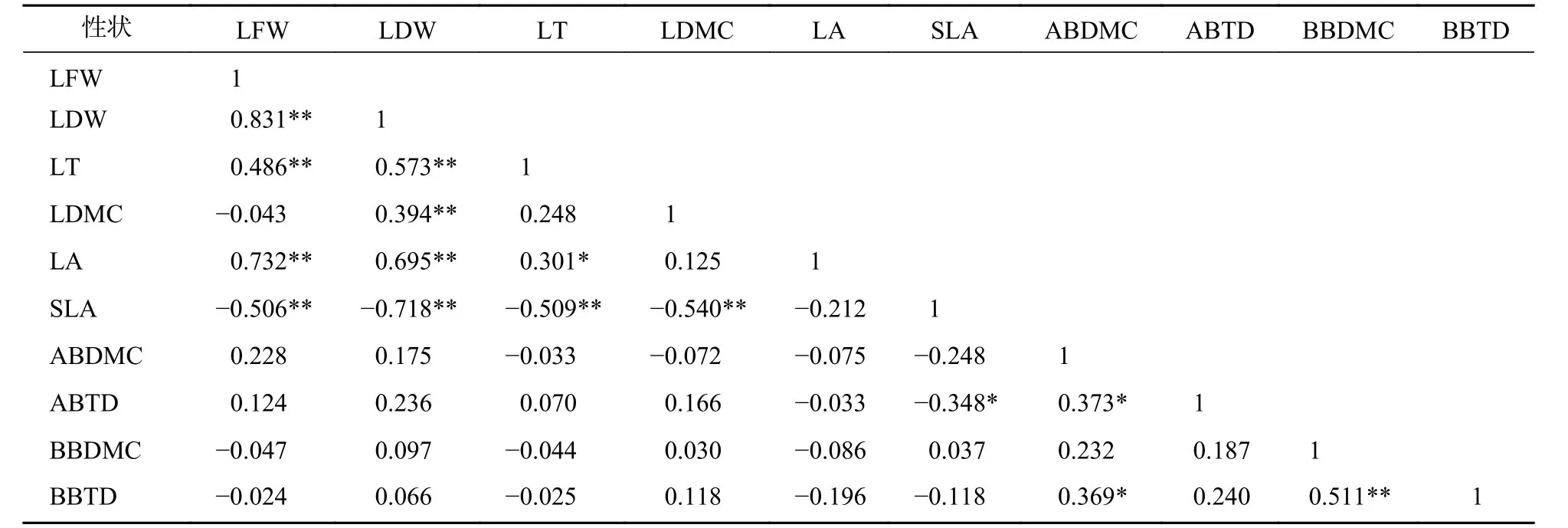
表5 中樹階段枝葉性狀之間的相關系數Table 5 Correlation coefficient between the twig and leaf traits in middle tree stage
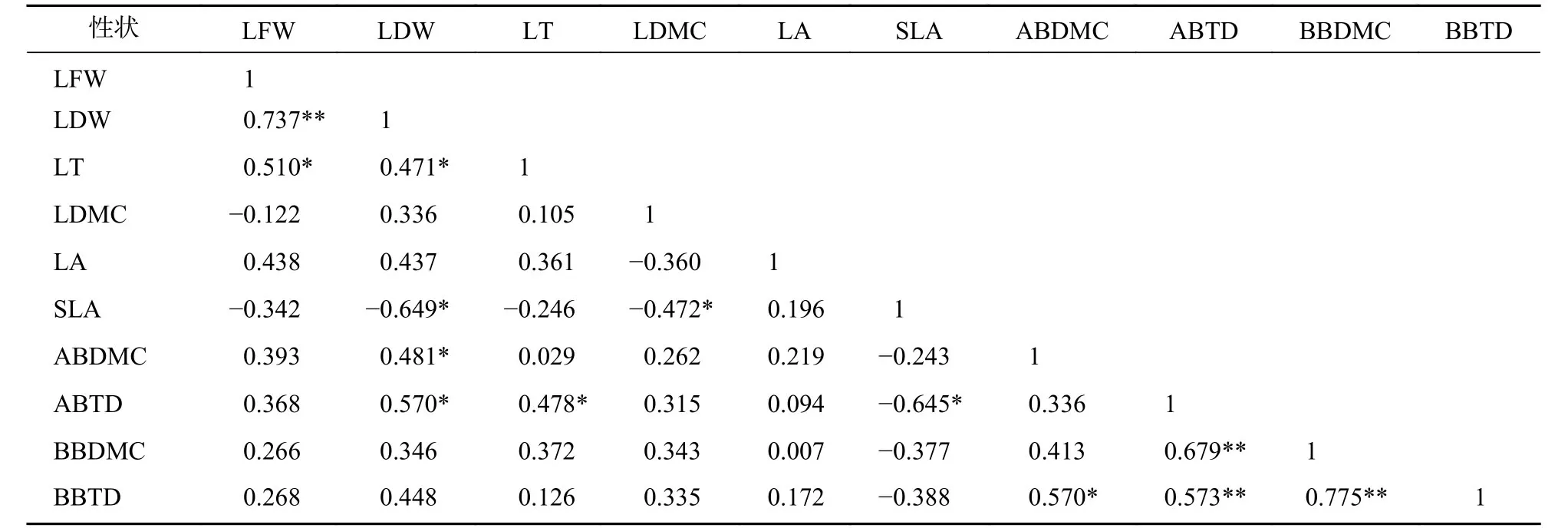
表6 小樹階段枝葉性狀之間的相關系數Table 6 Correlation coefficient between the twig and leaf traits in small tree stage
2.3 不同發育階段枝葉性狀半方差分析
在不同發育階段枝葉性狀中,選擇第1軸的林木個體枝葉性狀主成分得分進行半變異函數方差分析(表7和圖2)。對整體林分而言,單木整體性狀、葉性狀和枝性狀的結構方差比分別為25.2%、31.8%、0,單木整體性狀和葉性狀具有中等程度的空間自相關,枝性狀空間自相關性弱,擬合最優模型為線性,說明其性狀表現呈隨機分布。在大樹階段,單木整體、葉性狀和枝性狀的結構方差比分別為50.5%、50.4%和50.5%,具有中等的空間自相關程度,有效變程分別為75.60、73.70和212.70 m,擬合最優模型分別為球狀模型、球狀模型和指數模型,呈聚集分布,枝性狀的有效變程最大,說明枝性狀的空間連續性更大。在中樹和小樹階段,模型擬合效果差,且結構方差低,說明小樹階段枝葉性狀空間自相關性弱,不宜采用克里格插值(Kringing)空間預測。
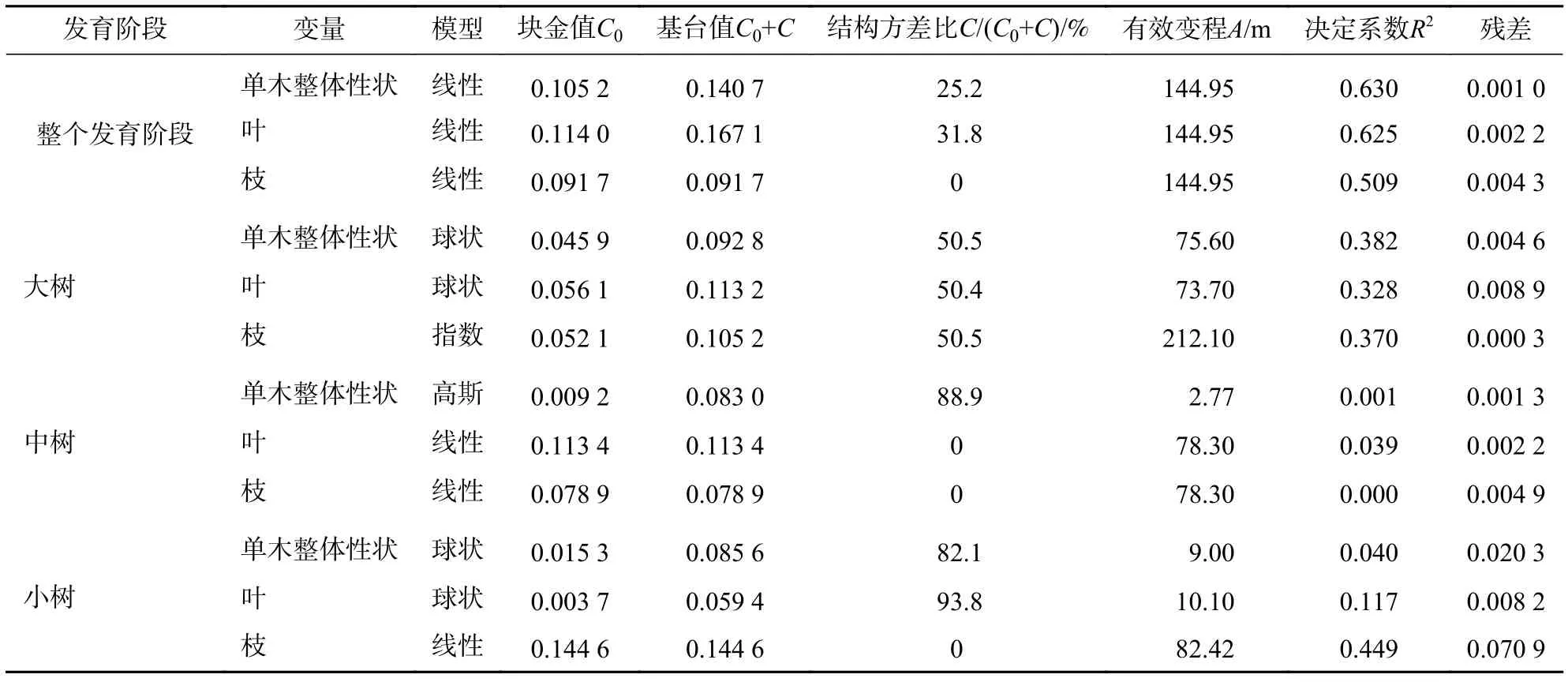
表7 枝葉性狀半方差模型及參數Table 7 Isotropic semi-variogram model and parameters of twig and leaf traits
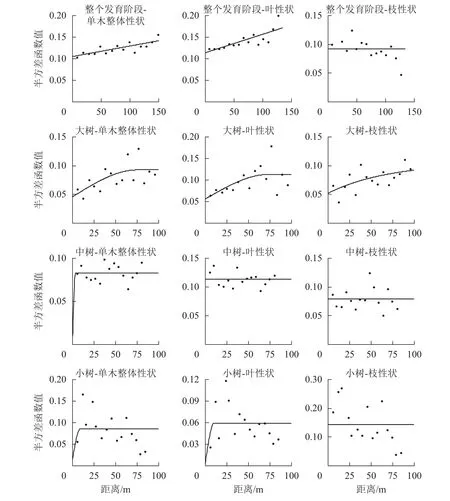
圖2 枝葉性狀半方差函數圖Figure 2 Isotropic semi-variogram of twig and leaf functional traits
3 討論
3.1 不同發育階段枝葉性狀的差異
植物之間的生存策略方式會在植物的性狀上表現出來。比葉面積與植物的光合利用效率有關[19],本研究中比葉面積隨著林木大小增加顯著降低,這與其他不同物種及生境下得到的研究結果一致[20]。耿夢婭等[21]通過對不同發育階段葉性狀的研究發現:發育后期的植物個體有著更大更厚的葉片,且不同的發育階段葉片性狀差異顯著,與本研究結果相符。較高的林木在進行長距離水分運輸時,必須克服阻力問題,而較高的莖干鮮比將有利于水分的運輸[22]。本研究發現:枝干鮮比與林木大小的關系不顯著,可能是研究區內濕度高,水資源沒有成為限制植物生長的環境因子,這與HE等[23]的研究結果一致。研究還發現:隨著林木的發育,小枝密度逐漸增大,高密度的小枝除了能夠加強對外界環境的抵抗外,還能夠承載更大更多的葉片;較低的小枝密度意味著有更低構造細胞的成本,有利于小樹階段枝條的快速生長[24]。這反映了種群的生態策略方式從小樹到大樹階段由快收益向慢收益轉變,這是物種的生存策略所決定的。自然界中,無論從單個物種的發育階段還是不同生活型的物種來看,大部分矮小的林木有著小的葉片,高大的林木個體葉片和枝密度更高[25],與本研究結果相符。
3.2 枝葉性狀間的相關性
本研究發現:在小枝與葉片的10個性狀中,葉性狀間的相關性強,與FORTUNEL等[26]的研究結果一致。部分性狀間在小樹階段相關性較低或不顯著,可能是在激烈的競爭環境下,由于生存策略的優化方式,不同器官間的權衡方式在各性狀間獨立運行的原因[27]。小枝與葉功能性狀相關性較弱,僅小枝密度與葉鮮質量和干質量呈正相關,然而MéNDEZ-ALONZO等[28]研究發現:莖密度與水分傳導速率呈顯著負相關,本研究結果與其相反,原因是莖的密度遠大于小枝密度,足以支持林木生長的需要,因此更應該考慮長距離運輸過程中高密度組織細胞帶來的水分運輸問題,這是莖與小枝不同的功能作用造成的。對莖葉經濟譜的研究發現[9,29]:莖經濟譜與葉經濟譜呈正交狀態,本研究中小枝與葉性狀在排序軸呈正交狀態,這是造成枝葉性狀相關性弱的重要原因。葉片質量與枝密度呈顯著正相關,較大的枝密度意味著小枝能夠承受更多的質量負荷。對不同發育階段葉片質量與枝密度之間的相關性分析發現:這種現象在大樹階段表現得更加顯著,其原因是小枝密度與葉生物量密切相關[6],本研究也發現大樹有著更大更厚的葉。通過研究4個枝性狀之間的關系,兩兩之間呈正相關,較高的小枝密度需要更多的結構組織,這種現象在不同的發育階段都有著不同程度的表現,小樹階段小枝性狀間相關性更大,這主要是因為小樹階段的生物學特征決定的。關于莖經濟譜[9]的研究表明:莖密度與莖干鮮比密切相關,本研究中小枝性狀間也出現這種相關性。對于莖經濟譜理論,同樣也能適用于小枝部分性狀的研究。
3.3 不同發育階段枝葉性狀空間特征
在地統計學分析中,整體林分的枝葉性狀分布模式以隨機分布為主,其空間自相關性為中等,其原因是天然林中闊葉樹種的分布方式常以隨機分布為主[29]。大樹階段,枝葉性狀以聚集分布為主,劉妍妍[30]通過對闊葉林的空間格局分析發現:具有較大葉片的個體通常以聚集形式分布,與本研究相符。葉片和小枝性狀是影響植物冠幅大小的重要因素,能夠影響成年植物冠層的結構形式及發展狀態,進而影響植物對資源的獲取能力。大樹階段枝性狀主成分得分的有效影響變程均大于整體和葉性狀,說明枝條的開展情況比葉的大小更能加劇大樹個體之間的影響。中、小樹階段枝葉性狀的擬合效果和結構方差比較小,不成連續變化狀態,中、小樹階段林木枝葉性狀的預測不宜采用克里格(Kringing)插值估算,與SIEFERT[14]的研究不一致,主要原因可能是中、小樹階段的空間格局分布方式為隨機分布,林木間的距離較遠,空間自相關性弱。若要預測樣地整體功能性狀的表現,需要測量更多林木個體的功能性狀值。
綜上所述,不同發育階段枝葉性狀差異顯著,大樹傾向于高質量的投資策略,小樹的投資策略為快速投資;不同發育階段葉性狀間和小枝性狀間相關性顯著;峨眉含笑種群大樹階段枝葉性狀具有空間自相關性。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
