目舒丸對兔眼睫狀體平滑肌細胞內舒縮傳導信號的影響
2021-03-07 08:21:12宿蕾艷張莎莎張明明魏春秀方素萍王穎胡瑛莊曾淵
中國中醫眼科雜志 2021年12期
宿蕾艷,張莎莎,張明明,魏春秀,方素萍,王穎,胡瑛,莊曾淵
眼部調節和集合功能異常,睫狀肌和內直肌功能紊亂,可導致神經支配的異常或肌肉的過度緊張,引起視疲勞。眼的調節功能是依賴睫狀體平滑肌的收縮和舒張來實現的,與細胞內游離鈣水平關系密切。大鼠肉瘤(rat sarcoma,RAS)同源基因家族成員A(ras homolog gene family member A,RhoA)及蛋白激酶C(protein kinase C,PKC)是平滑肌收縮的關鍵信號蛋白[1-3]。中醫認為視疲勞屬于“肝勞”,其核心病機是陰血不足、筋膜攣急。目舒丸為中國中醫科學院眼科醫院的院內制劑,具有活血養血,解痙止痛的功效,臨床上用于治療視疲勞,療效確切[4-5]。本實驗擬在細胞水平觀察目舒丸對兔眼睫狀體平滑肌細胞舒縮的影響,研究目舒丸治療視疲勞的機制,為其臨床合理應用提供數據支持。
1 材料與方法
1.1 實驗動物
實驗用3 月齡新西蘭大白兔,10 只,體重2.5~3.0 kg,購于北京維通利華實驗動物技術有限公司。飼養條件:12 h/12 h 交替光暗,溫度25℃,濕度55%左右,自由飲食、飲水,動物房環境保持安靜、通風良好。
1.2 主要試劑與儀器
氯化乙酰膽堿(上海梯希愛化成工業發展有限公司,批號:A0084);二氧化碳培養箱(美國Thermo公司,型號:HERA CELL 150);超凈工作臺(北京東聯哈爾儀器制造有限公司,型號:SCB-1360);倒置顯微鏡(美國,型號:OLYMPUS B071);冷凍離心機(美國Sigma 公司,型號:1-14PK),酶標儀(美國BioRad 公司,型號:1450);實時熒光定量聚合酶鏈式反應儀(美國BIO-RAD,型號:CFX96),基因擴增儀(美國BIO-RAD,型號:C1000)。
1.3 取材
取上述新西蘭大白兔,無菌條件下摘取兔眼球,用含雙抗的磷酸緩沖鹽溶液(phosphate buffered saline,PBS)沖洗,沿角膜緣后3 mm 處切開眼球,去除晶體,剪除虹膜,取下完整睫狀肌帶,沖洗后剪碎,加入3 mL V 型膠原酶消化2 h,終止消化,1500 r/min離心5 min,于沉淀中加入平滑肌細胞培養液,吹打均勻,得到原代兔眼睫狀體平滑肌細胞,置于37℃、5%CO2培養箱中培養。2~3 d 后細胞傳代培養,傳代得到第3 代睫狀體平滑肌細胞可用于后續相關實驗。
目舒丸溶液配置:稱取160 mg 目舒丸顆粒加入10 mL 細胞培養液充分溶解,之后使用0.45 μm 微孔濾膜進行過濾,得到濃度為16 mg/mL 的目舒丸濃溶液,濃溶液用于稀釋不同終濃度目舒丸溶液。
1.4 分組與給藥
將1.3 得到的第3 代睫狀體平滑肌細胞以1×106個/mL 均勻接種于5 個6 孔板中,鋪板24 h 后,隨機選出27 個接種孔,并隨機分為9組,每組3 個孔,分別命名為A組~I組。將1.3 得到的目舒丸溶液稀釋成7 個濃度,依次為8、4、2、1、0.5、0.25、0.125 mg/mL 并分別加入到A~G組中,繼續培養細胞10 min。H組予氯化乙酰膽堿(0.5 mmol/L)繼續培養2 h,I組予目舒丸溶液(1.0 mg/mL)繼續培養24 h,之后予氯化乙酰膽堿(0.5 mmol/L)培養2 h。
1.5 兔眼睫狀體平滑肌細胞長度測定
A組~G組在給藥前及給藥后的0.5、1、2、4 和10 min 這6 個時間點,分別在相差顯微鏡下用測微器測定細胞直徑,每個接種孔觀察6 個視野,每個視野測10 個細胞,取其平均值進行統計分析。
1.6 兔眼睫狀體平滑肌細胞內PKC、RhoA 基因表達
將H組和I組細胞離心,收集各組細胞,進行實時熒光定量聚合酶鏈式反應(real-time fluorescent quantitative polymerase chain reaction,RT-qPCR)。按照總核糖核酸(Ribonucleic Acid,RNA)抽提試劑說明書提取總RNA(表1)。反轉錄:各組樣品取1 μg總RNA,按M-MLV First Strand RT Kit 試劑盒說明書進行cDNA 反轉錄合成。聚合酶鏈式反應(polymerase chain reaction,PCR)檢測:以上述反轉錄cDNA 為模板,應用熒光定量PCR 檢測細胞中PKC、RhoA 的基因表達。25 μL 反應體系:SYBR Premix Ex Taq Ⅱ12.5 μL,上下游引物各1 μL,模板2 μL,滅菌雙蒸水補足至25 μL。擴增條件為預變性95℃3 min,變性94℃30 s,退火72℃3 min,以上2 步擴增循環。每個循環結束后收集熒光信號,反應結束后確認擴增曲線和融解曲線。再根據2-ΔΔCt公示計算目的基因的表達量。
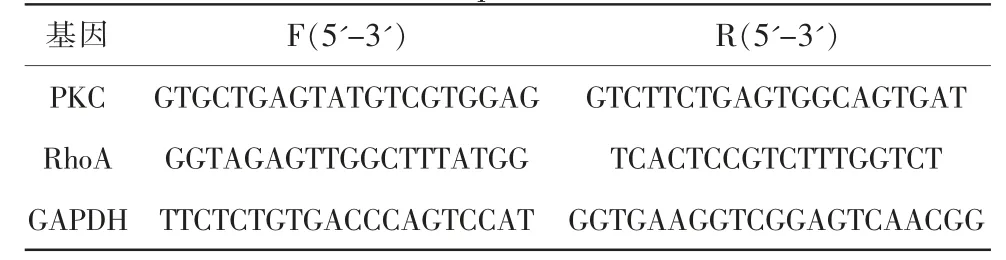
表1 RT-qPCR 引物序列
1.7 統計學方法
采用SPSS20.0 系統軟件進行統計處理。計量資料用均值±標準差()表示,多組間比較采用單因素方差分析,各組間兩兩比較采用LSD-t 檢驗,組內比較采用配對t 檢驗。以P<0.05 為差異有統計學意義。
2 結果
2.1 兔眼睫狀體平滑肌細胞形態及鑒定
兔眼睫狀體平滑肌細胞體外培養12 h 后,細胞開始貼壁,剛貼壁的細胞呈梭形,細胞相對較小,培養2 d 后,貼壁細胞增多,并逐漸伸展,原代細胞形態不一,多呈梭型,有少數細胞突起,呈單個生長或幾個細胞以突起相連接(圖1A)。
兔眼睫狀體平滑肌細胞免疫組化鑒定:培養的睫狀體平滑肌細胞為α-actin 免疫反應陽性,陽性物質位于胞漿內,呈棕黃色,胞核不著色;倒置顯微鏡下可見胞核無色,胞漿呈棕黃色(圖1B)。
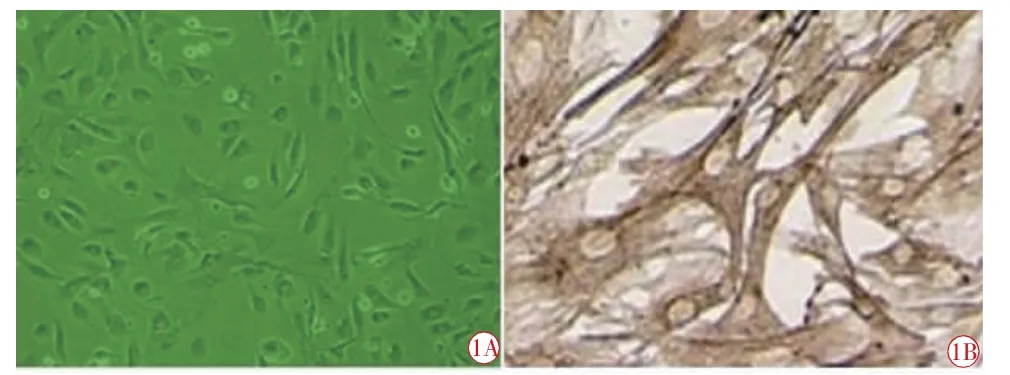
圖1 兔眼睫狀體平滑肌細胞形態及組化鑒定圖。1A 原代兔睫狀體平滑肌細胞形態圖(×200);1B 兔眼睫狀體平滑肌細胞為αactin 免疫反應陽性(×400),陽性物質位于胞漿內,呈棕黃色,細胞核不著色
2.2 兔眼睫狀體平滑肌細胞直徑比較
給藥前后比較:A組給藥后所有時間點細胞直徑較給藥前均縮短,差異均有統計學意義(t0.5min=6.347,t1min=6.582,t2min=10.870,t4min=11.480,t10min=5.005,均P=0.000)。B組給藥后所有時間點細胞直徑變化均無統計學意義(P>0.05)。C組給藥后4、10 min 細胞直徑均伸長,差異均有統計學意義(t4min=2.086,P=0.039;t10min=2.300,P=0.024);余時間點細胞直徑變化均無統計學意義(P>0.05)。D組給藥后1、2、4、10 min細胞直徑均伸長,差異均有統計學意義(t1min=2.794,P=0.006;t2min=3.294,P=0.001;t4min=3.109,P=0.003;t10min=2.821,P=0.006);給藥后0.5 min 細胞直徑變化無統計學意義(P>0.05)。E組給藥后2、4、10 min 細胞直徑均伸長,差異均有統計學意義(t2min=2.389,P=0.019;t4min=3.271,P=0.002;t10min=3.983,P=0.000);余時間點細胞直徑變化均無統計學意義(P>0.05)。F組給藥后所有時間點細胞直徑變化均無統計學意義(P>0.05)。G組給藥后所有時間點細胞直徑變化均無統計學意義(P>0.05)(表2)。
組間比較:與A組給藥后各時間點比較,B~G組細胞直徑均伸長,差異均有統計學意義(P<0.05)。與B組給藥后10 min 比較,E組細胞直徑縮短,差異有統計學意義(P<0.05),余組均無統計學意義(P>0.05);與余時間點比較,C~G組細胞直徑變化均無統計學意義(P>0.05)。與C組給藥后0.5 min 比較,E組細胞直徑縮短,差異有統計學意義(P<0.05),余組均無統計學意義(P>0.05);與給藥后1 min,D、G組細胞直徑伸長,差異有統計學意義(P<0.05),余組細胞直徑變化均無統計學意義(P>0.05);與余時間點比較,D~G組細胞直徑變化均無統計學意義(P>0.05)。與D 給藥后10 min 比較,E組細胞直徑伸長,差異有統計學意義(P<0.05),余組均無統計學意義(P>0.05);與余時間點比較,E~G組細胞直徑變化均無統計學意義(P>0.05)。與E組給藥后0.5 min 比較,F組細胞直徑伸長,差異有統計學意義(P<0.05),G組細胞直徑變化無統計學意義(P>0.05);與余時間點比較,F、G組細胞直徑變化均無統計學意義(P>0.05)。與F組給藥后2 min 比較,G組細胞直徑伸長,差異有統計學意義(P<0.05);與余時間點比較,G組細胞直徑變化均無統計學意義(P>0.05)(表2、3)。
表2 不同濃度目舒丸溶液干預前、后兔眼睫狀體平滑肌細胞直徑比較(,n=3,μm)
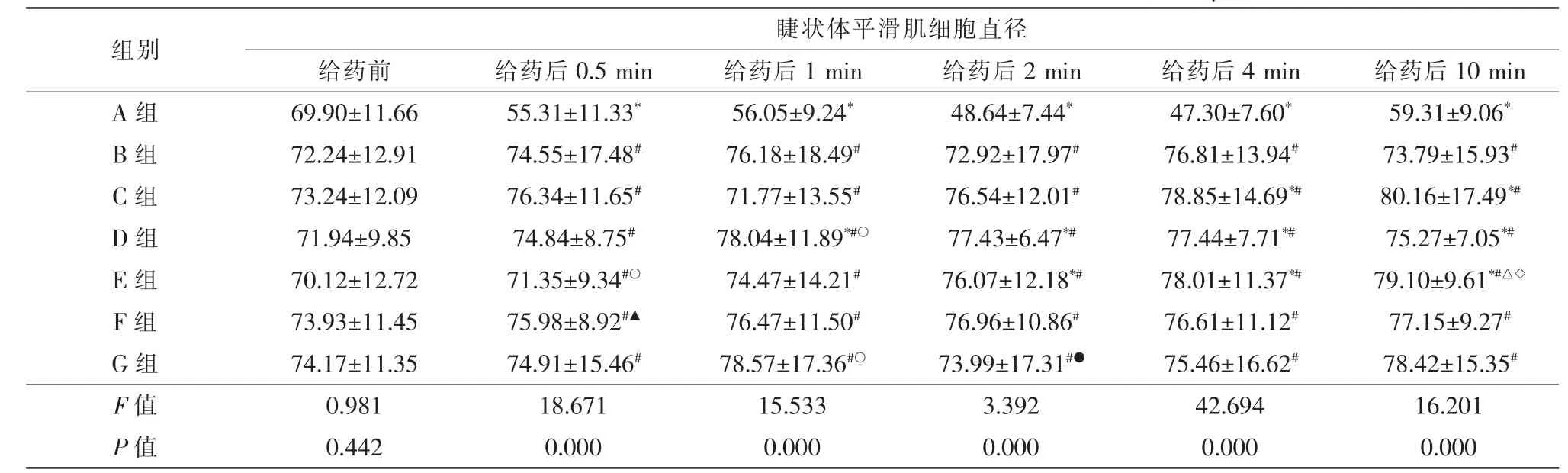
表2 不同濃度目舒丸溶液干預前、后兔眼睫狀體平滑肌細胞直徑比較(,n=3,μm)
注:* 與同組組給藥前比較,P<0.05;# 與A組比較,P<0.05;△與B組比較,P<0.05;○與C組比較,P<0.05;◇與D組比較,P<0.05;▲與E組比較,P<0.05;●與F組比較,P<0.05
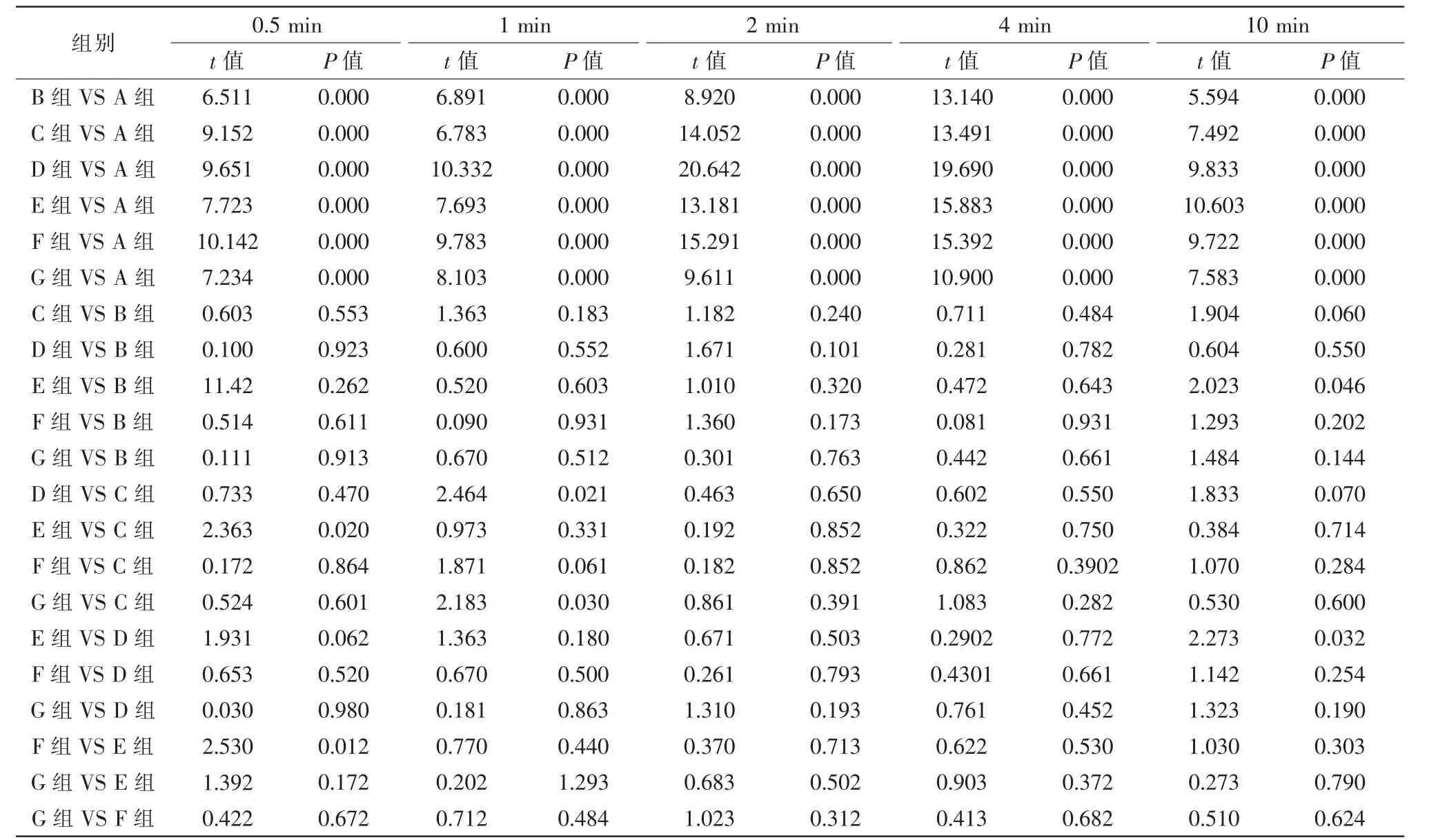
表3 不同濃度目舒丸溶液干預后各時間點兔眼睫狀體平滑肌細胞直徑統計值
2.3 兔眼睫狀體平滑肌細胞內PKC 及RhoA 基因表達
與H組比較,I組的PKC 及RhoA 表達量降低,差異均有統計學意義(tPKC=-3.566,P=0.023;tRhoA=-8.895,P=0.001)(表4)。
表4 RT-PCR 檢測結果(,n=3)
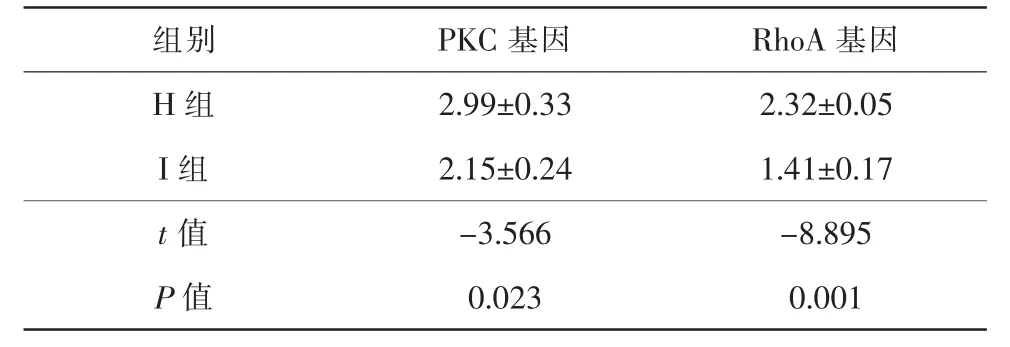
表4 RT-PCR 檢測結果(,n=3)
3 討論
中醫將視疲勞歸于“肝勞”,肝血耗損、過度使用腦力及目力,都會導致目竅失養,無法久視。調節痙攣是視疲勞的常見原因,現代工作方式造成調節痙攣發生的普遍性,但臨床除了屈光矯正,并無有效治療方法。
目舒丸為中國中醫科學院眼科醫院莊曾淵教授多年經驗方加工而成,治療眼視疲勞,癥見眼珠隱痛、頭額悶痛、不能久視、視力模糊、常欲閉目,臨床療效較好。本實驗設定了6 個時間水平,考察7 個目舒丸劑量梯度來觀察其對眼睫狀體平滑肌細胞舒縮作用時間、劑量效應關系,及對睫狀體平滑肌細胞內與收縮有關的鈣依賴性及非鈣依賴性信號傳導因子的影響。
原代培養的兔睫狀體平滑肌細胞具有很好的收縮功能,是研究睫狀體生理及藥理作用的良好實驗材料。本研究采用組織膠原酶消化法成功獲得原代兔睫狀肌細胞,細胞呈梭形,α-actin 染色強陽性,與已有國外文獻一致[6-7]。原代睫狀體平滑肌細胞給予不同濃度目舒丸干預一定時長后發現,目舒丸可顯著影響睫狀肌細胞的收縮狀態,并呈現一定的時間依賴性,尤其是1.0 mg/mL 目舒丸干預細胞10 min內,發現其能顯著舒張平滑肌。
RhoA 是單體G 蛋白中結構較為保守的蛋白,其C 端可以參與正確定位蛋白與翻譯后的修飾,N 端則具有可以與鳥苷三磷酸(guanosine-5′-triphosphate,GTP)結合并參與水解的大部分氨基酸[8]。RhoA 結構中存在具有催化功能的氨基酸,這些氨基酸主要在GTP 水解過程中發生作用。肌動蛋白聚合受到RhoA 蛋白的影響,促使細胞粘附,肌動球蛋白收縮等。目前,參與細胞動力學的重要信號通路是RhoA-ROCK[9]。RhoA 激活Rho 相關激酶,促進細胞粘附、改變細胞形態以及促使細胞骨架拉伸,Rho 激酶(Rho-associated kinase,ROCK)活化狀態持續,激活肌球蛋白輕鏈磷酸化會促使ROCK 進一步拉伸細胞骨架,同時加強RhoA-ROCK 的耦合。ROCK 激活可以促使肌球蛋白輕鏈(myosin light chain,MLC)磷酸化而產生肌絲收縮作用,同時活化后ROCK 的底物是磷酸化肌球蛋白輕鏈激酶(myosin light-chain phosphatase,MLCP)。磷酸化是在接受Rho/ROCK 的活化信號后產生的,失活后,同時也使磷酸化的MLC 脫磷酸失活狀態的終止,從而提高胞漿內磷酸化MLC 水平,增加肌球-肌動蛋白交聯。在鈣水平相同的條件下,MLC 磷酸化的程度增加,細胞收縮強度增加,說明收縮蛋白對鈣的敏感性的增加,這一過程稱為鈣敏感性調節。
PKC 是一種存在于胞質的依賴鈣和磷脂酶的蛋白激酶,其活性影響著細胞的增殖、凋亡等生物學活動[9]。另外,PKC 可以通過p44/42MAPK 信號級聯反應使鈣黏蛋白(cadherin,CaD)和P-鈣粘蛋白(Pcadherin,CaP)發生磷酸化,促使它們對肌球蛋白ATP 酶活性的抑制作用消失,從而調節平滑肌細胞的收縮[10-11]。
本研究發現,在加入氯化乙酰膽堿24 h 前加入目舒丸,發生作用后,與單純加入氯化乙酰膽堿組相比,細胞內PKC 及RhoA 表達量下降,推測目舒丸可通過降低睫狀體平滑肌細胞內PKC 及RhoA 的表達,減少非鈣依賴通道引起的睫狀體平滑肌細胞收縮,進而改善睫狀體平滑肌過度收縮引起的眼部調節功能異常。
綜上,本研究發現目舒丸可通過降低睫狀體平滑肌細胞內PKC 及RhoA 的表達,減少非鈣依賴通道引起的睫狀體平滑肌細胞收縮,進而改善睫狀體平滑肌過度收縮引起的眼部調節功能異常。
猜你喜歡
新少年(2022年9期)2022-09-17 07:10:54
音樂探索(2022年2期)2022-05-30 21:01:37
小天使·一年級語數英綜合(2020年6期)2020-12-16 02:56:41
文苑(2020年12期)2020-04-13 00:54:10
中學生數理化·中考版(2019年12期)2019-09-23 06:23:28
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
北極光(2014年8期)2015-03-30 02:50:51
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
