走馬胎活性組分對肝癌HepG2 細胞DUSPs/MAPK 信號通路的影響
2021-03-09 10:11:22廖長秀瑩黃桂坤潘勇韋建開
中成藥 2021年2期
關鍵詞:肝癌
賀 珊 廖長秀 羅 瑩黃桂坤潘 勇韋建開
(1.右江民族醫學院藥學院,廣西百色 533000;2.廣西肝膽疾病臨床醫學研究中心,廣西百色 533000;3.右江民族醫學院基礎醫學院,廣西百色 533000)
走馬胎Ardisia gigantifolia Stapf 為紫金牛科紫金牛屬植物走馬胎的干燥根莖,是廣西民間應用較多的藥物。有祛風濕、壯筋骨、活血祛瘀的功效,主治風濕筋骨疼痛、跌打損傷、產后血瘀和癰疽潰瘍等[1-2]。研究發現走馬胎主要成分有三萜皂苷類、沒食子酸等[3],其中三萜皂苷類成分對多種腫瘤細胞均具有較強的抗腫瘤作用[4-6]。課題組前期研究發現走馬胎水提液和醇提液及活性組分群均可明顯抑制肝癌細胞增殖、侵襲和轉移,并誘導其凋亡[7-8],但其抗肝癌機制尚不清楚。課題組前期采用基因表達譜芯片篩查發現走馬胎水提液可顯著升高肝癌細胞雙特異性磷酸酶(dual specificity protein phosphatase,DUSPs)家族DUSP1、 DUSP4 和DUSP5 mRNA 表達水平(分別為8.5 倍、6.5 倍和19 倍)。DUSPs 家族中大部分成員為絲裂原激活的蛋白激酶(mitogen-activated protein kinase,MAPK)信號通路的負向調節劑,參與腫瘤細胞的增殖、凋亡、侵襲轉移和耐藥等過程[9-10]。本研究觀察走馬胎活性組分對肝癌細胞DUSP1/4/5 mRNA 和蛋白表達及其下游ERK/JNK/p38-MAPK信號通路的影響,以期明確走馬胎活性組分抗肝癌的可能分子機制。
1 材料
1.1 細胞株 人肝癌HepG2細胞購于中國科學院細胞庫。
1.2 試劑 走馬胎購于廣西玉林中藥材市場,由右江民族醫學院覃道光副教授鑒定為走馬胎根莖。胎牛血清(烏拉圭Lonsera 公司,批號MI09331);DMEM 高糖培養基(美國 Gibco 公司,批號8117208);索拉非尼(美國MedChemExpress 公司,批號 10003);胰蛋白酶-EDTA 消化液(0.25%,不含酚紅)、青鏈霉素混合液和MTT(北京索萊寶科技有限公司,批號分別為20160422、20160525、822A054)。RNA 提取試劑盒(美國 ZYMO RESEARCH 公司,批號ZRC00362);FastKing 一步法除基因組cDNA 第一鏈合成預混試劑和熒光定量PCR 試劑盒[天根生化科技(北京)有限公司,批號分別為S7614、S7717];PCR 引物由生工生物工程(上海)股份有限公司合成。BCA 蛋白定量試劑盒(增強型,批號為080919191105)及Western blot 常規試劑購于上海碧云天生物技術有限公司;兔抗DUSP1/4/5 單抗(英國Abcam 公司,批號分別為GR201669-3、GR266778-1 和GR201795-8);兔抗ERK、p-ERK、JNK、p-JNK,p38、p-p38(美國 Cell Signaling Technology 公司,批號分別為04、07、4、4、20、10);鼠抗GADPH 單抗、山羊抗鼠二抗、山羊抗兔二抗(北京中杉金橋生物技術有限公司,批號分別為TA-08、ZB-2305、ZB-2301)。
2 方法
2.1 分組及給藥 走馬胎活性組分的提取分離和總三萜皂苷含量測定參照文獻[8]。走馬胎藥材粉碎后加入6 倍量60%乙醇回流提取3 次,每次2 h,合并3 次提取液,減壓回收乙醇,并冷凍干燥為干粉,為制得的走馬胎乙醇提取物。經水飽和正丁醇后上D101 大孔樹脂,分別用蒸餾水、30%乙醇、50% 乙醇、70% 乙醇、95% 乙醇進行洗脫,各部分洗脫液經減壓回收乙醇和真空冷凍干燥處理后得到干粉。經MTT 實驗發現70%乙醇洗脫部位抗肝癌活性最強。70% 乙醇洗脫組分加甲醇溶解后,加于小孔樹脂凝膠柱,分別用蒸餾水、30%甲醇、50%甲醇、70% 甲醇、100% 甲醇進行洗脫,各部分洗脫液經減壓回收得到干粉。再經MTT 實驗發現小孔樹脂凝膠柱70%甲醇洗脫組分抗肝癌活性最強,硅膠薄層分析證明小孔樹脂凝膠柱70%甲醇洗脫成分主要成分為三萜皂苷類,以齊墩果酸為對照品,紫外分光光度計檢測其總三萜皂苷含量為39.50%。小孔樹脂凝膠柱70%甲醇洗脫組分即為本研究采用的走馬胎活性組分,和索拉非尼用DMSO 分別制備成4、8 mg/mL 和5 mmol/L,臨用前以完全培養基(含10%胎牛血清、1%青鏈霉素混合液的DMEM 培養基)按1 ∶1 000 稀釋為終濃度4 μg/mL、8 μg/mL、5 μmol/L 的培養液(0.1% DMSO)。實驗分為4 組,空白對照組、索拉非尼組、走馬胎活性組分(4、8 μg/mL)。取對數生長的HepG2 細胞以每孔3×105個細胞鋪于6孔板,貼壁培養24 h 后加藥處理,分別加入含不同質量濃度走馬胎活性組分(4、8 μg/mL)或5 μmol/L的索拉非尼的完全培養基,空白對照組加入等體積的含0.1% DMSO 完全培養基。繼續培養24、72 h,分別提取HepG2 細胞RNA 和蛋白進行后續實驗。
2.2 RT-PCR 檢測DUSPs 家族mRNA 表達 按RNA 提取試劑盒說明提取總RNA,用紫外分光光度法檢測總RNA 純度和濃度,取1 μg RNA 按試劑盒說明進行逆轉錄,取合成的cDNA 進行實時熒光定量PCR,以GAPDH 為內參,引物由Primer Premier 5.0 軟件設計,引物序列、產物大小見表1。反應體系按試劑盒說明,反應條件均為,預變性95 ℃15 min,變性95 ℃10 s,退火/延伸60 ℃30 s,共40 個循環(Roche LightCycler96 實時熒光定量PCR 儀擴增)。根據相對定量計算公式=(E1為目的基因的擴增效率,E2為內參的擴增效率,ΔCt1為目的基因對照組平均Ct與樣本Ct差值,ΔCt2為內參基因樣本Ct與對照組平均Ct差值)對每個基因標準曲線擴增效率進行校正后,分析實驗結果。實驗重復3次。取PCR 產物進行1.5%瓊脂糖電泳判斷產物的特異性。
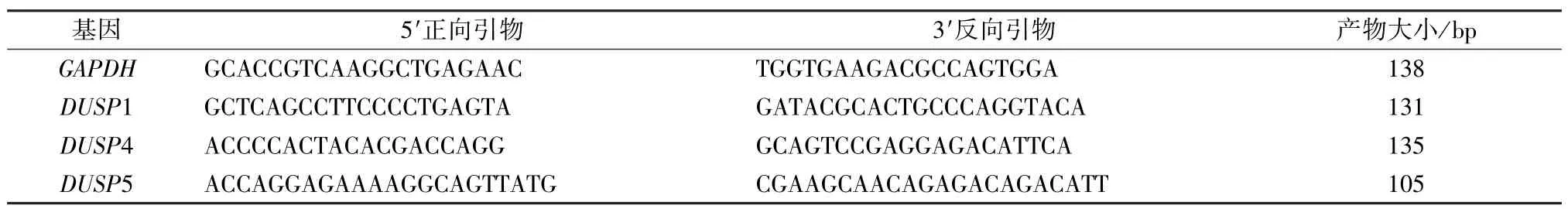
表1 實時定量PCR 引物序列及產物大小Tab.1 Primer sequences and product sizes of real-time quantitative PCR
2.3 Western blot 檢測蛋白表達 提取細胞總蛋白,用BCA 法測蛋白濃度。各組樣品加入5×上樣緩沖液,震蕩混勻后,加熱煮沸5 min 使蛋白充分變性。SDS-PAGE 垂直電泳后,轉膜。轉膜后BSA封閉液室溫封閉1 h。然后孵育一抗(內參抗體為小鼠抗GADPH 單抗,目的抗體為兔抗DUSP1、DUSP4、DUSP5、ERK、p-ERK、JNK、p-JNK、p38、p-p38 單抗,稀釋比均為1 ∶1 000),4 ℃搖床過夜。TBST 洗3 次,每次10 min。進一步室溫孵育二抗(內參蛋白二抗為山羊抗小鼠IgG 辣根酶標記,目的蛋白二抗為山羊抗兔IgG 辣根酶標記,稀釋比均為1 ∶5 000),室溫孵育2 h。TBST 洗脫二抗(5 min/次,大于5 次),然后暗室曝光。蛋白相對表達量=(目的蛋白灰度值給藥組/相對應的內參蛋白灰度值)/(目的蛋白灰度值空白組/相對應的內參蛋白灰度值)。
2.4 統計學分析 采用SPSS 20.0 軟件進行數據分析,數據以()表示,多組間均數比較采用單因素方差分析,組間兩兩比較采用LSD 檢驗,方差不齊采用Dunnett’s T3 檢驗。以P<0.05 為差異有統計學意義。
3 結果
3.1 走馬胎活性組分對肝癌HepG2 細胞DUSP1/4/5 mRNA 和蛋白表達的影響 與空白對照組比較,走馬胎活性組分低質量濃度組(4 μg/mL)作用24 h 后可升高HepG2 細胞DUSP1 mRNA,作用72 h HepG2 細胞后使DUSP1/4/5 mRNA 表達均明顯升高(P<0.05 或P<0.01);走馬胎活性組分高質量濃度組(8 μg/mL)作用24、72 h 可使HepG2 細胞DUSP 1/4/5 mRNA 表達水平均明顯升高(P<0.01)。且隨作用時間和走馬胎活性組分濃度增加,HepG2 細胞DUSP1、 DUSP4 mRNA 增加更為明顯。走馬胎活性組分低和高質量濃度組不論作用24 h 還是72 h 均能顯著升高DUSP1/4/5 蛋白表達,與空白對照組比較,差異有統計學意義(P<0.01)。
與空白對照組比較,5 μmol/L 索拉非尼處理HepG2 細胞24 h DUSP1 mRNA 表達升高(P <0.05),作用72 h 則可升高DUSP4、 DUSP5 mRNA表達(P <0.01)。處理24、72 h 均可明顯升高DUSP1/4/5 蛋白表達,與空白對照組比較,差異有統計學意義(P<0.01)。見圖1~2。
3.2 走馬胎活性組分對肝癌HepG2 細胞MAPK 信號通路蛋白表達的影響 與空白對照組比較,走馬胎活性組分(4、8 μg/mL)作用24、72 h 的HepG2 細胞ERK、JNK 和p38 磷酸化蛋白表達均明顯降低,差異性有統計學意義(P<0.01)。且隨走馬胎活性組分濃度和作用時間增加,HepG2 細胞ERK 磷酸化蛋白表達降低更為明顯,見圖3。5 μmol/L索拉非尼處理HepG2 細胞24、72 h 對ERK、JNK 和p38 磷酸化水平均有所降低,與空白對照組比較,差異有統計學意義(P<0.01)。
4 討論
MAPK 是細胞內一類絲氨酸/蘇氨酸蛋白激酶,MAPKs 信號轉導通路存在于大多數細胞內,可將細胞內的信號轉導到細胞外,參與細胞的生長和分化、調節細胞周期和凋亡以及血管的生成等[11]。目前已經確定有4 條并行的MAPK 相關信號轉導通路ERK1/2、JNK、p38 和ERK5 通路,通過調節肝癌細胞的增殖、凋亡和侵襲轉移[12-15],在肝癌發生過程中起著重要的調控作用,成為肝癌防治的重要作用靶點。本研究發現隨著走馬胎活性組分濃度增加和作用時間延長,MAPK 信號通路中ERK、JNK/MAPK 和p38 磷酸化蛋白表達明顯降低。提示走馬胎活性組分可能通過調控ERK/JNK/p-38 蛋白的磷酸化水平從而達到抑制肝癌發生發展的作用。
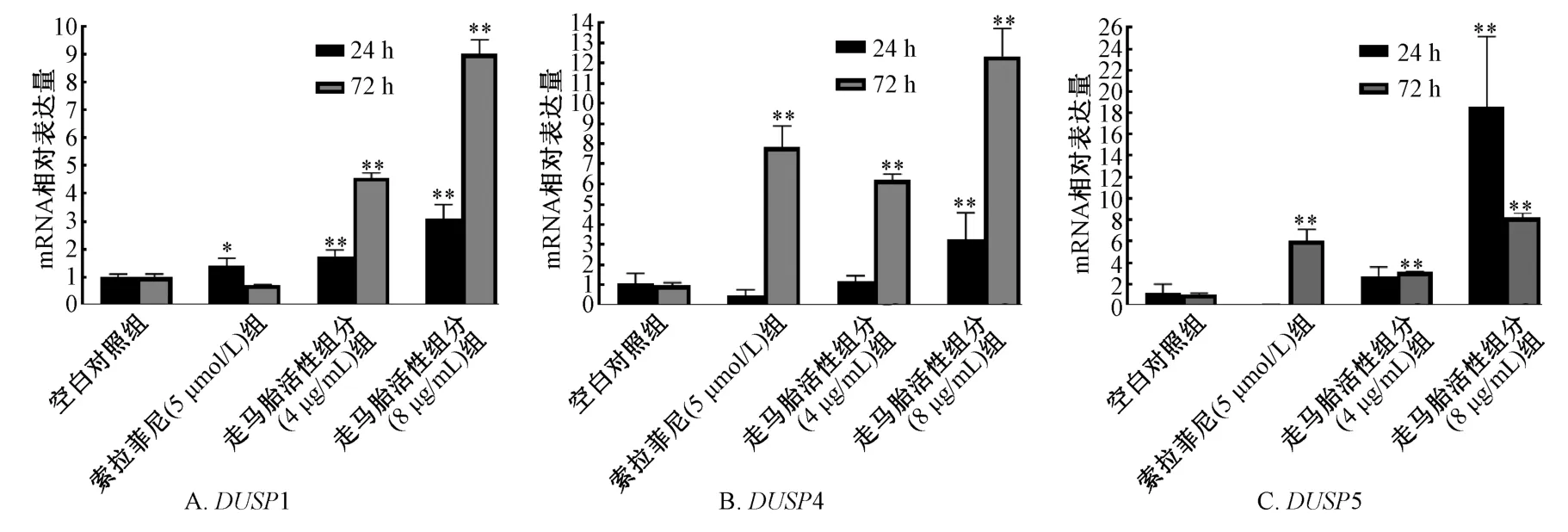
圖1 走馬胎活性組分對HepG2 細胞DUSP1/4/5 mRNA 表達的影響Fig.1 Effects of active components of A.gigantifolia on the expression of DUSP1/4/5 mRNA in HepG2 cells
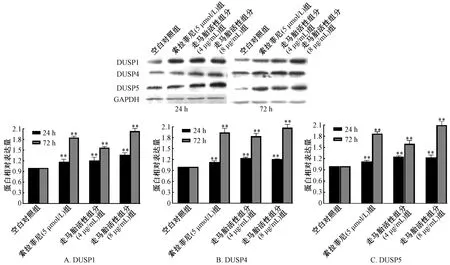
圖2 走馬胎活性組分對HepG2 細胞DUSP1/4/5 蛋白表達的影響Fig.2 Effects of active components of A.gigantifolia on the expression of DUSP1/4/5 protein in HepG2 cells
DUSP1/4/5 是蛋白質酪氨酸磷酸酶(protein tyrosine phosphatases,PTPs)超家族的成員,其在氨基酸一級結構上有PTP 保守序列,在一個底物內可同時使磷酸酪氨酸殘基、磷酸絲氨酸/蘇氨酸殘基去磷酸化,使MAPK 通路失活,調控腫瘤細胞的增殖、凋亡、侵襲和轉移等[16]。有研究報道DUSP1 低表達與低分化和肝細胞癌進展相關,而DUSP1 高表達則可抑制肝癌,延長肝癌病人生存期,機制可能與抑制p38 MAPK 磷酸化和促進p53磷酸化有關[17]。另有研究表明DUSP4 基因組的缺失可增加胰腺上皮內腫瘤的侵襲性,提示DUSP4可能抑制胰腺癌的侵襲[18]。Staege 等[19]發現霍奇金淋巴瘤細胞株中DUSP5 高表達進而調控促凋亡因子BCL2L11 而達到抗腫瘤的作用[19]。本實驗研究發現,隨著走馬胎活性組分濃度和作用時間的增加,DUSP1/4/5 的mRNA 和蛋白表達與對照組比較,都有不同程度的升高,呈現劑量-時間依賴性。結合文獻報道和本實驗對MAPK 信號通路關鍵蛋白的研究,提示走馬胎活性組分可能是通過升高DUSP1/4/5 的表達,使MAPK 信號通路失活,從而抑制肝癌的發生發展。
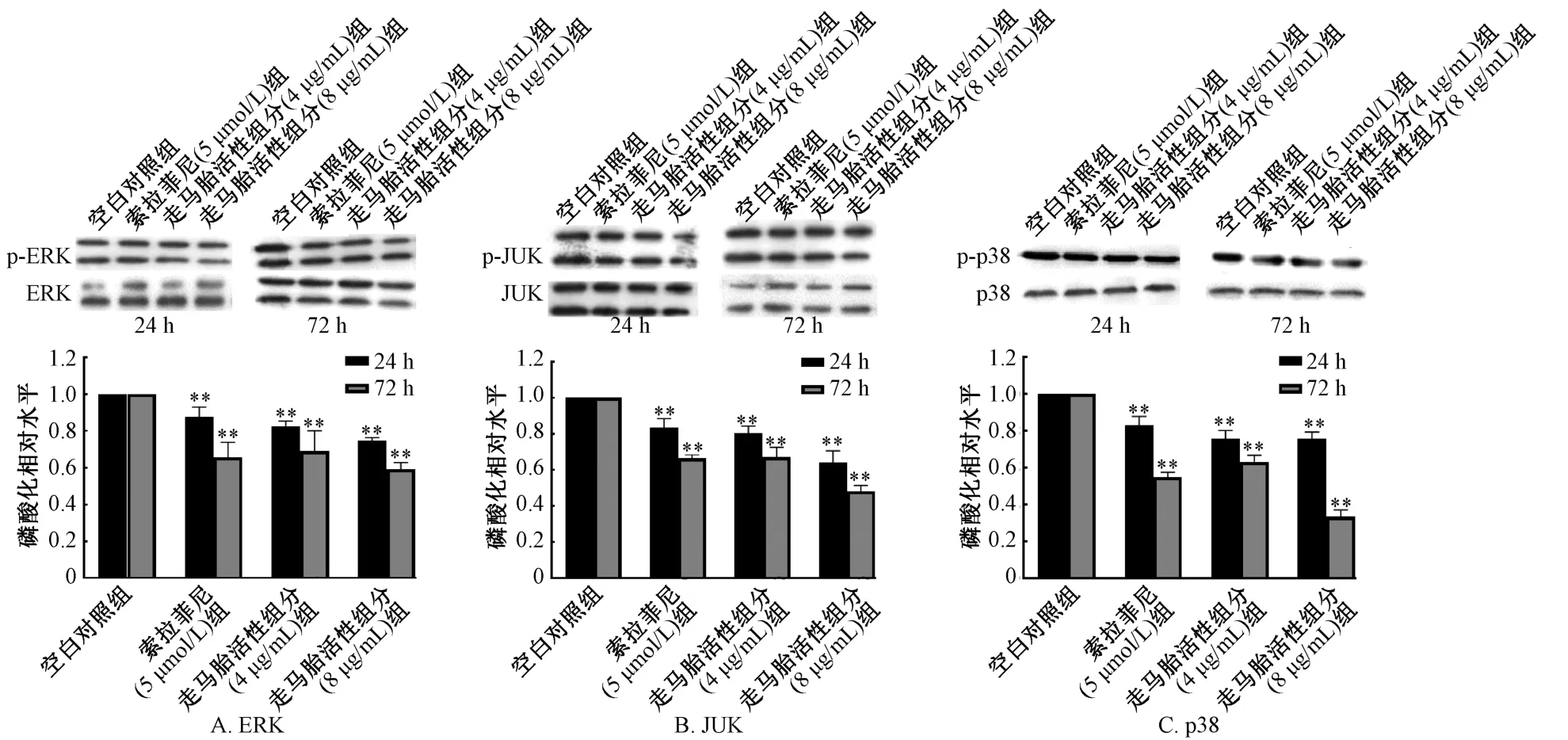
圖3 走馬胎活性組分對HepG2 細胞MAPK 信號通路磷酸化蛋白的影響Fig.3 Effects of active components of A.gigantifolia on the expression of phosphorylation protein of MAPK signal pathway in HepG2 cells
由以上可知,走馬胎抗腫瘤活性組分的抗肝癌作用可能是通過影響DUSPs 家族成員(DUSP1、DUSP4 和DUSP5)mRNA 和蛋白的表達水平,進而影響下游的ERK/JNK/p38-MAPK 信號通路實現的,但具體的相互作用機制,有待下一步通過DUSP1/4/5 激動劑和抑制劑或通過siRNA 和過表達質粒慢病毒轉染調控DUSP1/4/5 表達等實驗進行驗證。
猜你喜歡
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
中成藥(2016年8期)2016-05-17 06:08:14
癌癥進展(2016年12期)2016-03-20 13:16:17
罕少疾病雜志(2016年5期)2016-03-11 16:34:44
吉林大學學報(醫學版)(2015年1期)2015-12-17 07:47:28
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
中國醫藥導報(2015年26期)2015-02-28 22:07:59
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:34
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:24
