雜交蘭花色花香生物合成途徑的轉錄組分析
2021-03-10 01:40:48樊榮輝林榕燕葉秀仙鐘淮欽
西北植物學報 2021年1期
孔 蘭,樊榮輝,林榕燕,葉秀仙,林 兵,鐘淮欽*
(1 福建省農業科學院 作物研究所, 福州 350013; 2 福建省特色花卉工程技術研究中心, 福州 350013)
花色、花香作為觀賞植物重要的表型性狀,決定了蘭花的觀賞價值和經濟價值[1-2]。雜交蘭(Cymbidiumhybrid)是蘭屬中的國蘭和大花蕙蘭(Cymbidiumhybridium)雜交培育而成的一類蘭花的特稱,繼承了國蘭的幽香和大花蕙蘭花大、色艷、花箭出架等優異特性,植株適中,花期長,具有廣闊的市場前景[3-5]。此外,雜交蘭花色和花香多樣,也是研究花色和花香形成機制的理想材料[6-7]。
目前,國內外已開展了觀賞型植物花色素和香氣成分分析[8-9]。不同花色雜交蘭色素成分檢測發現,花青素決定了雜交蘭紅色素和紫色色素的積累,而類胡蘿卜素影響了黃色色素的積累[10-11]。陳藝荃等[12]應用頂空-固相微萃取/氣相色譜-質譜聯用技術分析了7個雜交蘭品種的花香成分,發現萜烯類化合物是其主要花香成分,不同品種間花香成分存在顯著差異。上述研究表明,雜交蘭不同品種的花色素和花香成分生物合成存在異同。
前人基于轉錄組數據,分析了蝴蝶蘭[13]、文心蘭[14]、卡特蘭[15]、春蘭[16]和建蘭[17]等蘭科植物花色素和花香成分生物合成的主要功能基因及其差異表達,建立了花色花香形成的分子調控網絡。但對雜交蘭特定的花色和花香、相關分子機制仍待研究。
雜交蘭K18為黃色花,其花香成分包括石竹烯、α-香柑油烯、反式橙花叔醇、β-姜黃烯和β-羅勒烯等化合物,主成分為倍半萜;K24的花為紫紅色,其花香成分包括桉油精、正己醇和反式法呢醛等化合物,主成分為單萜[12]。因此,本研究以K18和K24為材料,采用RNA-Seq技術獲得雜交蘭不同花期的花朵轉錄組數據,分析雜交蘭不同花色/花香相關基因的表達調控變化,為深入研究其分子機制以及蘭花品種改良提供有效信息。
1 材料和方法
1.1 材 料
黃花清香型K18(‘玉鳳’)為本實驗室引進品種,紫紅花淡香型K24(‘福韻丹霞’)是本實驗室大花蕙蘭和墨蘭雜交而來的自育品種,均種植于福建省農科院作物所花卉研究室苗圃中。分別采集K18和K24小花蕾期、半開期和盛花期的花朵(圖1),-80 ℃冷凍保存,用于后續RNA-seq分析。每個樣品3個生物學重復。
1.2 方 法
1.2.1 RNA提取及測序使用TaKaRa植物RNA提取試劑盒(Cat#9769)提取雜交蘭不同花期的總RNA。利用1%瓊脂糖凝膠電泳檢測RNA降解和污染;分別使用NanoPhotometer分光光度計、Qubit?2.0熒光計和Agilent bioanalyzer 2100系統檢測RNA純度、濃度和RNA完整性;根據NEB-Next?UltraTMRNA文庫制備試劑盒提供的說明書構建18個不同花期樣品(含3個生物學重復)的測序文庫,并利用Illumina高通量測序平臺進行測序,生成兩端測序Reads。測序工作由北京諾禾致源生物公司完成。

A~ C. K18小花蕾期、半開期和盛花期;D~ F. K24小花蕾期、半開期和盛花期圖1 K18 和 K24 不同花期表型A-C. Bud stage, half flowering stage and flowering stage in K18;D-F. Bud stage, half flowering stage and flowering stage in K24Fig.1 Phenotypes of K18 and K24 at different flowering stages
1.2.2 功能注釋和表達差異分析采用Trinity[18]軟件對轉錄組Clean reads進行拼接獲得轉錄本序列。根據序列相似性,比對到Nr、Nt、Swiss-Prot、KEGG、GO(e<1e-5)、Pfam(e<0.01)和COG(e<0.001)七大數據庫中,獲得相應基因的功能注釋。基于Nr和Pfam獲得的蛋白功能注釋結果,利用KAAS[19]軟件對基因進行KO注釋,完成KEGG代謝通路分類。采用DESeq R[20]軟件包分析不同花發育時期的差異表達基因,并通過多重假設檢驗校正。readcount數值為基因差異表達的分析數據,校正后的P<0.05且|log2(foldchange)|>1設置為基因顯著差異表達的篩選閾值。使用KOBAS[21]軟件進行差異表達基因的KEGG通路富集分析,校正后P<0.05的Pathway為顯著富集的代謝通路。
1.2.3 qRT-PCR驗證以雜交蘭小花蕾、半開期和盛花期花朵的總RNA為模板,按照PrimeScript RT reagent kit with gDNA Eeaser試劑盒進行反轉錄合成cDNA。參照TB GreenTMPremix Ex TaqTM試劑盒說明書,以cDNA為模板,在ABI7500 PCR儀上完成qRT-PCR。用2-ΔΔCt方法計算每個基因在不同樣品中的相對表達量。Actin為內參基因。
1.2.4 花色素苷成分檢測參照徐怡倩等[22]的方法,應用超高效液相色譜-四級桿飛行時間質譜聯用儀(UHPLC-QTOF-MS)檢測雜交蘭花色素苷成分。
2 結果與分析
2.1 轉錄組數據組裝與功能注釋
K18和K24樣品測序的原始數據經質控和處理,獲得446.52 M有效數據,共66.97 Gb,Q30在92.00%以上,GC含量均大于46.11%,表明測序數據可靠。有效數據組裝后獲得787 529和336 170條轉錄本(transcript)和單基因(unigene);其中unigene平均長度為599 bp,N50長度為868 bp,組裝完整性較高,可用于后續分析。通過公共數據庫比對,36.13% unigene獲得功能注釋(表1)。其中,Nr數據庫匹配到的unigene數目最多,為86 272個(25.66%);GO數據庫匹配到50 174個unigene(14.92%);KEGG數據庫中得到18 789個unigene(5.58%)注釋。
2.2 轉錄組差異表達基因分析及KEGG功能分類
對K18和K24不同花期的差異表達基因(differently expressed gene,DEG)比較結果(圖2)發現,K18中,較盛花期分別有11 914和1 082個基因在小花蕾期和半開期差異表達;K24中,較盛花期分別有6 793和459個基因在小花蕾期和半開期差異表達。K18-1 vs K18-3或K24-1 vs K24-3比較組中涵蓋了全部的DEG,且上調基因數目明顯低于下調基因數目。此外,本研究比較了小花蕾期和盛花
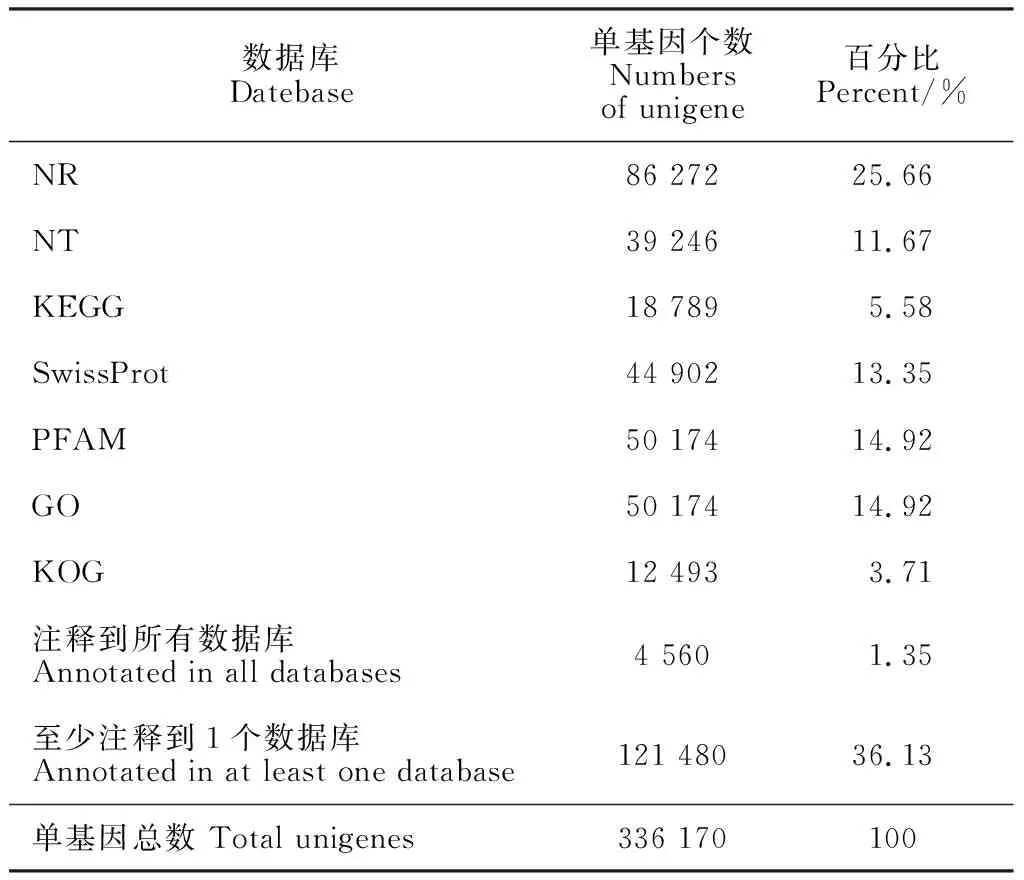
表1 功能注釋結果統計

圖2 雜交蘭不同時期差異表達基因的分布Fig.2 Distribution of DEGs among different flower developmental stages
期 K18 與 K24 之間的DEG,其中K18-1 vs K24-1獲得17 920個DEG,K18-3 vs K24-3獲得13 161個DEG。
分別對K18-1 vs K18-3和K24-1 vs K24-3的11 914個和6 793個DEG比對到KEGG數據庫中。K18-1 vs K18-3的DEG在淀粉和蔗糖代謝(ko00500,228)富集最顯著,其次是類胡蘿卜素生物合成(ko00906,50)、亞油酸代謝(ko00591,47)和萜類骨架生物合成(ko00900,66)等。K24-1 vs K24-3富集最多的是苯丙烷類生物合成(ko00940,130)、萜類骨架生物合成(ko00900,59)和α-亞麻酸代謝(ko00592,71)等。已明確的是,類胡蘿卜素生物合成途徑、類黃酮生物合成途徑介導了花色的形成,而萜類骨架生物合成途徑、倍半萜和三萜生物合成途徑與花香直接相關。
2.3 花色素生物合成相關結構基因分析
13個花色相關差異表達基因參與類胡蘿卜素生物合成途徑(表2)。如圖3,A顯示,與小花蕾期相比,K18中八氫番茄紅素合成酶基因(PSY)、八氫番茄紅素脫氫酶基因(PDS)和ζ-胡蘿卜素異構酶基因(Z-ISO)等的表達量在盛花期均上調;番茄紅素環化酶基因(LYCE和LYCB)和β-胡蘿卜素羥化酶基因(BCH1)的表達量下調。K24盛花期與小花蕾期相比,上調的基因有PSY、玉米黃質環氧酶基因(ZEP)和VDE;下調的基因有PDS、Z-ISO和BCH1等。
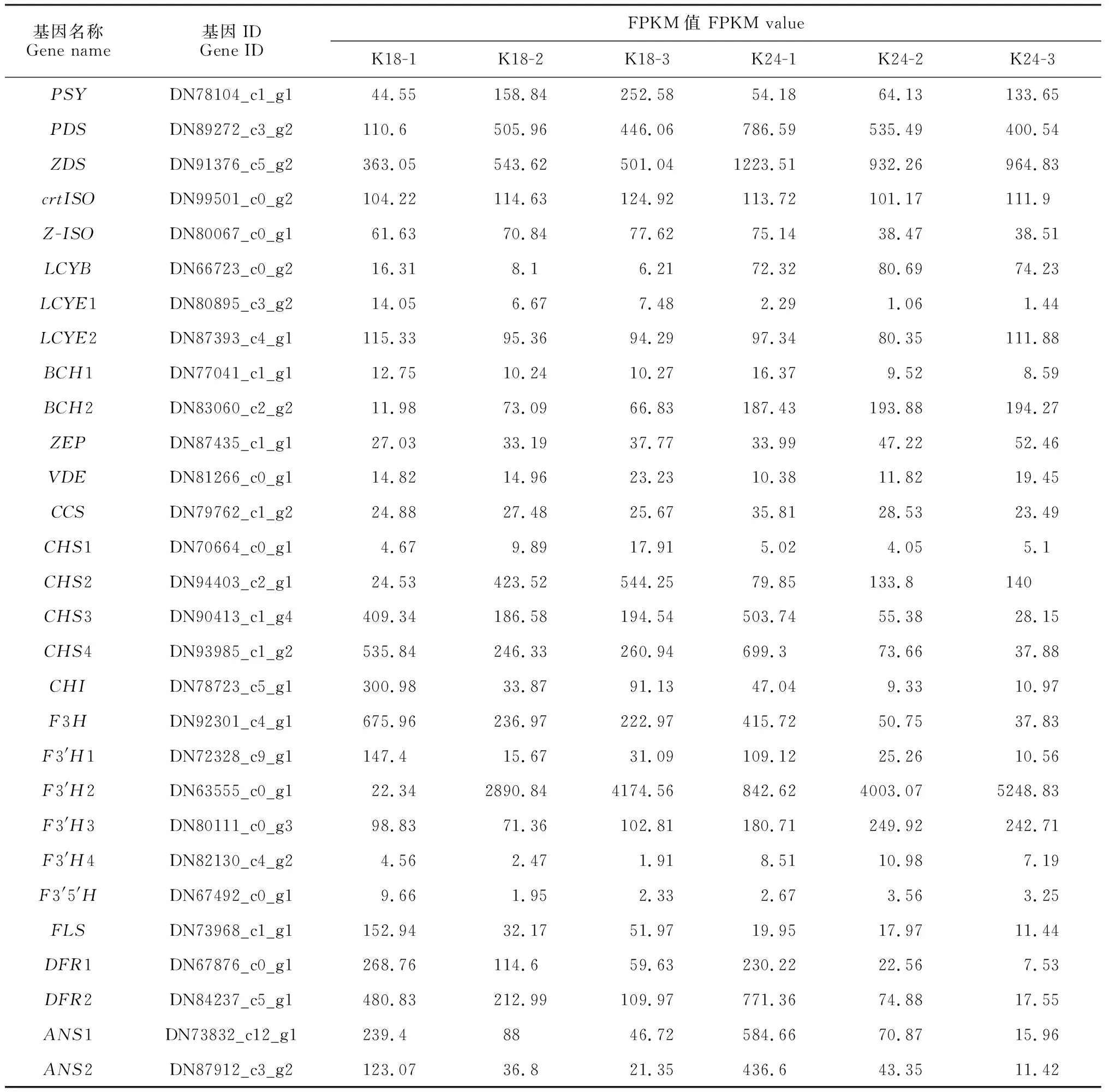
表2 花色結構基因的差異表達
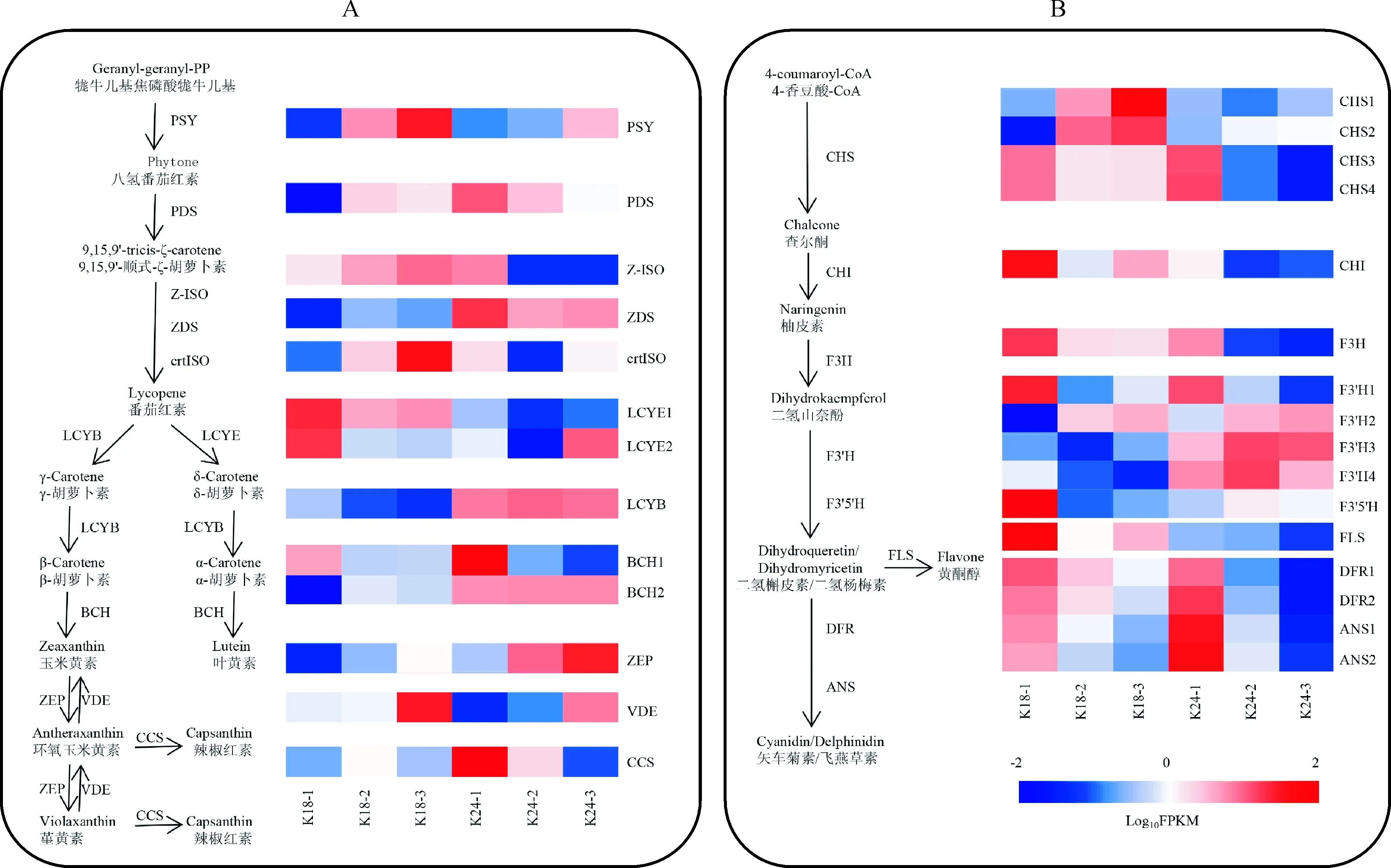
A. 類胡蘿卜素生物合成途徑中結構基因的表達譜;B. 類黃酮生物合成途徑中結構基因的表達譜圖3 K18 和 K24 中花色素結構基因的表達譜A. The expression profile of structural genes involved in carotenoid biosynthesis; B. The expression profile of structural genes involved in anthocyanin biosynthesisFig.3 Expression profiles of structural genes associated with pigment biosynthesis in K18 and K24 at three flower developmental stages
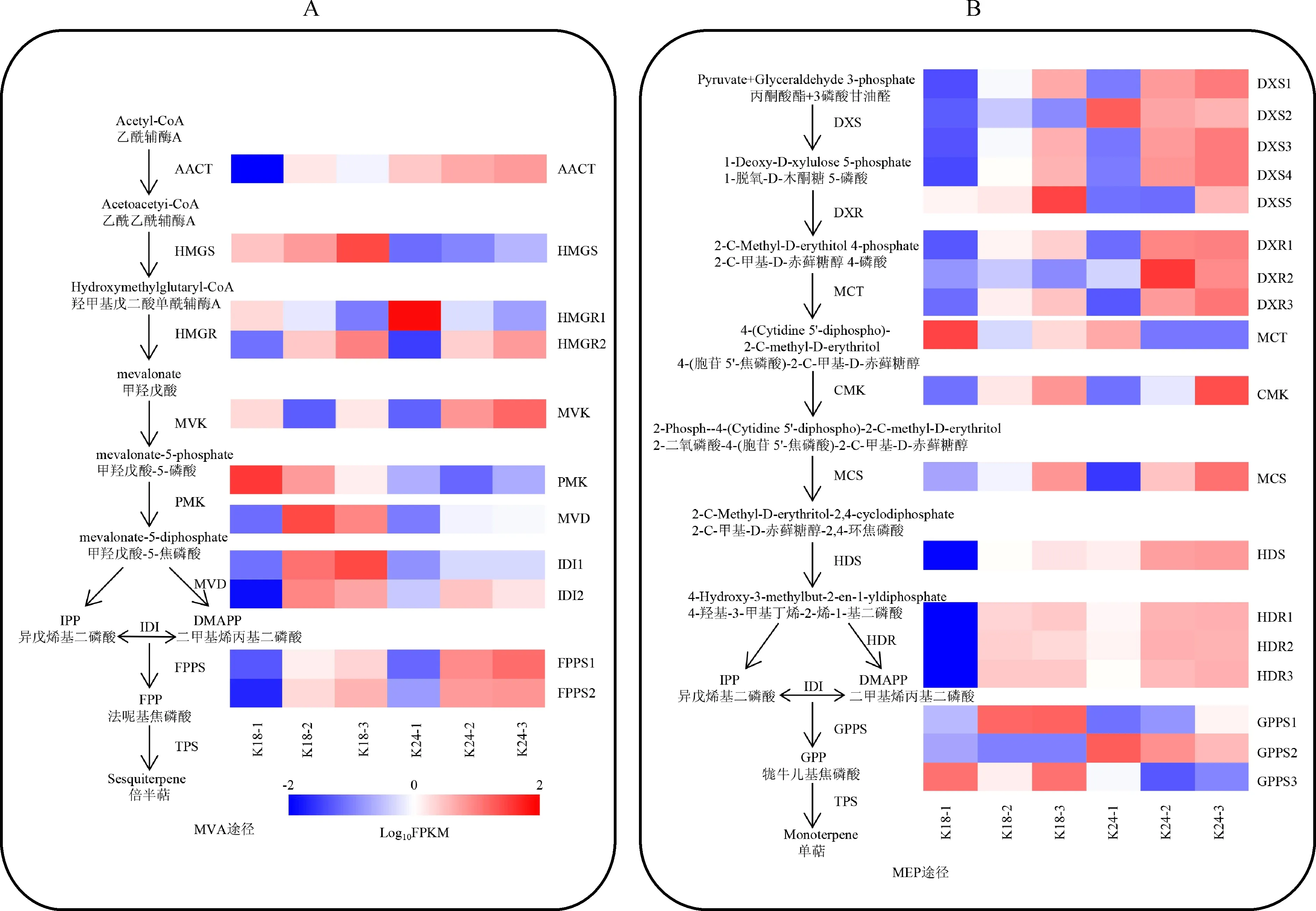
A. 倍半萜生物合成途徑中結構基因的表達譜;B. 單萜生物合成途徑中結構基因的表達譜圖4 K18 和 K24 中萜類合成相關結構基因的表達譜A. Expression profile of structural genes involved in sesquiterpene biosynthesis; B. Expression profile of structural genes involved in monoterpene biosynthesisFig.4 Expression profiles of structural genes associated with terpenoid biosynthesis in K18 and K24
16個差異表達基因參與類黃酮生物合成途徑(表2)。除類黃酮-3′,5′羥化酶基因(F3′5′H)外,其他類黃酮生物合成關鍵基因在 K18 和 K24 中均表現相似的表達模式。其中,查爾酮異構酶基因(CHI)、黃烷酮3-羥基酶基因(F3H)和黃酮醇合酶基因(FLS)等均表現下調,查耳酮合酶基因(CHS)和類黃酮-3′羥化酶基因(F3′H)既有上調表達也有下調表達;F3′5′H在K18的小花蕾期表達水平最高,在K24的3個時期均無明顯差異表達(圖3,B)。
2.4 雜交蘭萜類生物合成相關的結構基因
28個花香結構基因參與萜類骨架生物合成途徑,其不同花發育時期的表達水平具有顯著差異(圖4,表3)。甲羥戊酸(MVA)途徑中羥甲基戊二酰輔酶基因(HMGS)、羥基-3-甲基戊二酰輔酶A還原酶基因(HMGR2)、甲羥戊酸-5-焦磷酸合酶基因(MVD)及異戊烯基焦磷酸異構酶基因(IDI)的表達水平在半開期或盛花期顯著上調,且K18的表達量明顯高于K24(圖4,A)。2-C-甲基-D-赤蘚醇-4-磷酸(MEP)途徑中除2-C-甲基-D-赤蘚糖醇4-磷酸胞苷基轉移酶基因(MCT)和牻牛兒基焦磷酸合酶基因(GPPS)外,其他酶基因在盛花期高表達,且在K24中優勢表達(圖4,B)。
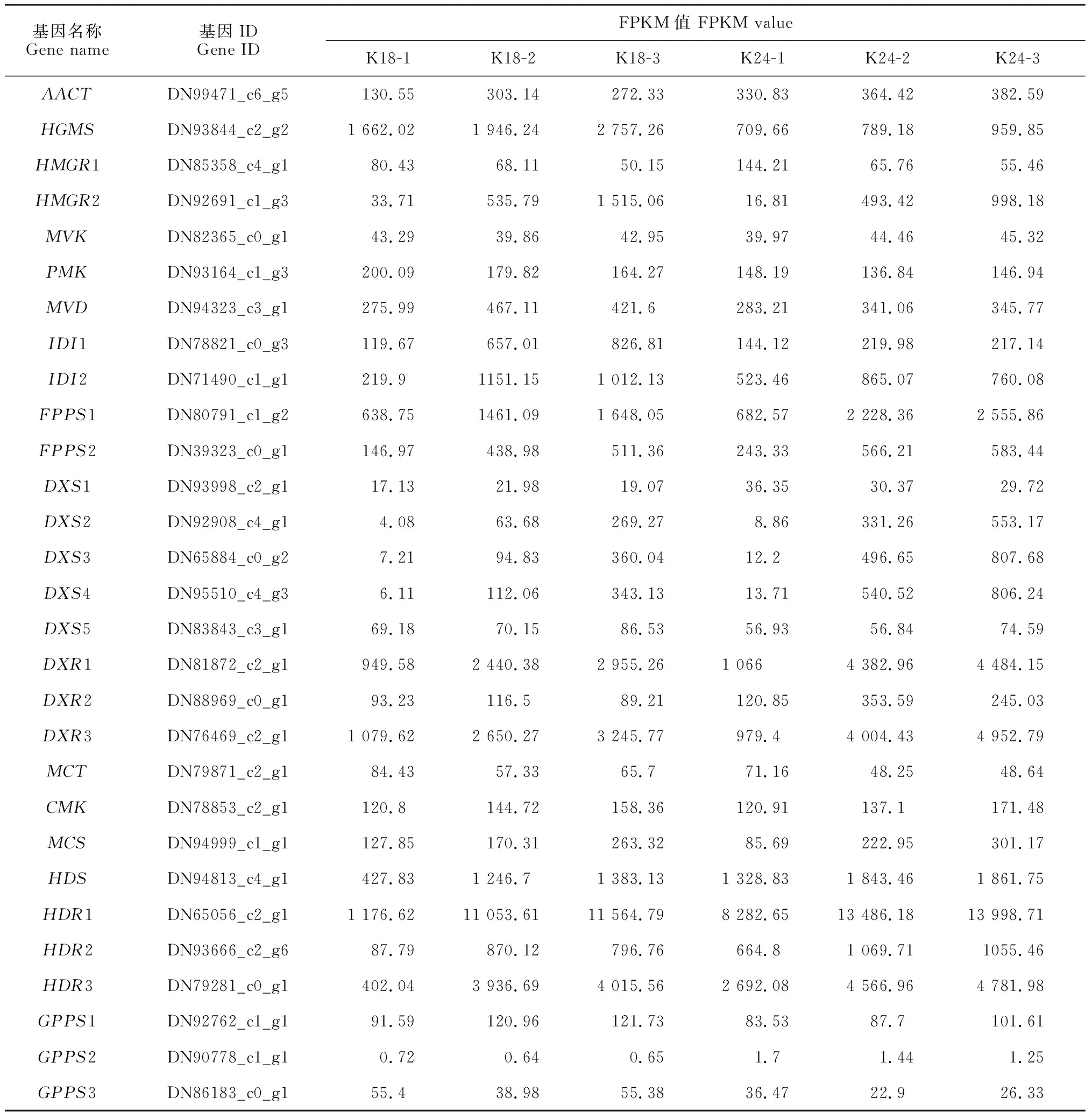
表3 花香結構基因的差異表達
此外,本研究篩選到112個差異表達的萜烯合酶基因(TPS),多數TPS的表達水平在盛花期最高,與香氣釋放呈正相關。其中,倍半萜生物合成途徑中DN78169_c2_g2(alpha-humulene synthase-like)、DN99176_c3_g1(Beta-cubebene synthase)、DN80925_c0_g1(germacrene D synthase)和單萜生物合成途徑中DN64363_c2_g4(beta-ocimene synthase)、DN72253_c0_g1(terpene synthase 10-like)和DN83821_c0_g4(linalool synthase)在K18盛花期的轉錄水平顯著上調,且高于K24(>100倍)。而單萜合酶基因DN65004_c3_g1、DN87421_c2_g1(alpha-terpineol synthase)和DN78962_c1_g1(terpene synthase 10-like)在K24盛花期的轉錄水平高于K18。
2.5 花色素苷成分檢測
對雜交蘭K18和K24盛花期花瓣花色素苷進行定量定性分析,結果顯示,共有18種花色素苷,花色素苷總含量存在顯著差異,K24中花色苷含量為608.74 μg·g-1,K18花色苷含量僅為K24的20%。K24花中色素物質主要成分為矢車菊素苷,使其花色呈紅色;K18中飛燕草素苷含量相對較高(表4)。
2.6 花色花香結構基因的qRT-PCR分析
對16個花色花香結構基因進行qRT-PCR分析,其表達結果與RNA-Seq的測序結果相似,驗證了RNA-Seq數據的可靠性(圖5)。花色結構基因LYCE、F3H和FLS在小花蕾期表達量最高,且K18中表達水平高于K24;ANS則在K24的小花蕾期顯著表達。花香結構基因DXS2和TPS1-8在盛花期的表達水平明顯高于小花蕾期和半開期,其中TPS1、TPS2和TPS5-8在K18盛花期中的表達量約為K24的100倍以上。

表4 K18和K24中花色素苷成分鑒定

K18-1、K24-1. 小花蕾期;K18-2、K24-2. 半開期;K18-3、K24-3. 盛花期圖5 雜交蘭花色花香合成相關基因的表達情況K18-1, K24-1. Bud; K18-2, K24-2. Half flowering; K18-3, K24-3. Full floweringFig.5 Expression analysis of genes involved in pigment biosynthesis and terpenoid biosynthesis in Cymbidium hybrid
3 討 論
本研究對2個雜交蘭品種的不同時期花進行轉錄組測序,共獲得了336 170個unigene,其中36.13%的unigene成功注釋到數據庫。基于KEGG通路富集和表達譜分析,獲得了58個花色花香相關差異表達基因及其在不同發育階段花中的表達豐度(FPKM值),這些關鍵差異基因在雜交蘭花色花香形成過程中具有重要作用。
類胡蘿卜素和類黃酮是植物的兩大呈色物質,其生物合成途徑得到了系統的研究[23-25]。其中類黃酮生物合成途徑結構基因CHS、CHI、F3H、F3′H、F3′5′H、FLS、DFR和ANS已相繼被克隆和功能分析,這些關鍵結構基因的高表達可影響色素的積累,進而決定從淺黃到紅色或藍紫色等花色的呈現[26]。前人研究表明[27-29],ANS的表達可促進紅色花的形成,而F3′5′H和FLS的高表達可促進黃酮醇物質的積累,從而使花呈現黃色。本研究發現類黃酮生物合成途徑結構基因整體上在小花蕾期表達量最高,盛花期顯著降低,與文心蘭花色相關基因的表達模式相似[14]。然而,CHI、F3′5′H、FLS和ANS的表達水平在 K18 和 K24 間存在顯著差異(圖3,B),其中ANS在K24的表達量明顯較高,可能是K24花瓣中花色苷含量高于K18的原因之一;K18花中雖含有少量飛燕草素和飛燕草3-蕓香糖苷,但其花朵顏色為黃色,轉錄組數據和qPCR結果顯示CHI、F3′5′H和FLS在K18中顯著高表達,推測K18通過CHI、F3′5′H、FLS途徑積累了大量黃酮醇而不是花青素,從而影響了花朵顏色的決定。此外,前期課題組研究發現K18花中未檢測出類胡蘿卜素,轉錄組數據顯示其類胡蘿卜素生物合成途徑除上游結構基因PSY、PDS、ZDS、ctrISO表達較高外,下游結構基因的表達水平偏低,這可能是造成類胡蘿卜素合成受阻的原因。
萜烯類化合物是雜交蘭主要的花香物質。目前春蘭[16]、臘梅[30]、百合[31]、姜花[32]等花卉中萜類化合物生物合成途徑已研究得較為清楚。Ramya等[16]指出FPPS、HMGS、HMGR和AACT的表達量與春蘭倍半萜物質金合歡醇的合成和釋放呈正相關。本研究發現K18倍半萜合成途徑的HMGS、HMGR2、MVD、IDI和FPPS的表達水平均在盛花期最高(圖4,A),與香氣釋放規律一致,推測這些基因的表達為K18倍半萜物質的合成提供上游底物。與K18相比,K24的香氣較弱,而單萜合成途徑的DXR和DXS在K24的表達量明顯高于K18(圖4,B),前人研究結果表明DXR和DXS的表達促進了單萜類物質的合成[33-34],推測DXR和DXS是K24主要花香成分單萜物質合成的關鍵基因。此外,本研究中TPS的表達水平隨花開放進程總體呈上升趨勢,且在K18中的表達量高于K24,解釋了K18比K24香氣更清香的原因。
目前,雖已克隆了較多的花色和花香相關基因,但這些基因多集中在模式植物中,且它們的分子調控機理還有待完善。本研究通過雜交蘭不同時期花轉錄組測序,挖掘到大量與類黃酮生物合成途徑和萜類生物合成途徑相關的酶基因,豐富了相關途徑的基因庫。在后續研究中可進一步結合蛋白組學和代謝組學分析驗證相關基因轉錄水平與蛋白組水平和代謝組水平的一致性。另外,花青素和花香物質的合成由調節因子和結構基因共同決定的[32,35-36],后期可結合調控因子對結構基因的轉錄調控和轉基因分析,闡明雜交蘭花色花香相關基因的生物學功能和分子機制,為今后雜交蘭花色花香的定向改良和新品種選育提供了可靠依據。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
音樂探索(2022年2期)2022-05-30 21:01:37
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
航空世界(2020年10期)2020-01-19 14:36:20
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
湖南教育·C版(2018年3期)2018-06-05 16:54:36
