基于2b-RAD簡化基因組測序的甜瓜遺傳多樣性分析
2021-03-10 08:21:56曹燕燕刁倩楠陳幼源張永平
西北植物學報 2021年1期
關鍵詞:分析
曹燕燕,刁倩楠,陳幼源,張永平
(上海市農業科學院 園藝研究所,上海市設施園藝技術重點實驗室,上海 201403)
甜瓜(CucumismeloL.)為葫蘆科甜瓜屬一年生蔓性草本植物,是國內外重要的園藝和經濟作物。甜瓜果實不僅味美香甜,多汁爽口,而且具有很高的營養和藥用價值,因此深受廣大消費者的喜愛,特別是在夏季水果市場中占有十分重要的地位。為了滿足市民多元化高質量的消費需求,迫切需要育種工作者培育出更多新的高品質甜瓜品種。甜瓜種質資源是進行新品種選育以及開展科研工作的重要基礎,因此加強甜瓜特異種質資源的開發和利用對促進甜瓜產業的發展和科研水平的提升具有重要意義[1]。
作為葫蘆科植物中表型變異最為豐富的作物之一,甜瓜在株型、葉片和果實等形態特征方面具有廣泛的多樣性,尤其是果皮顏色、果面覆紋、果肉顏色、果肉硬度、可溶性固形物含量以及風味等的果實表型的變異最為豐富[2-4]。目前,關于甜瓜遺傳多樣性的研究國內外已有報道。例如,Escribano S.等[5]研究了不同皮色希臘甜瓜的遺傳多樣性;王煒勇等[6]利用表型性狀研究了浙江省沿海地區27份薄皮甜瓜地方品種的遺傳多樣性;胡建斌等[7]對中國各地257份甜瓜地方種質資源果實與種子的形態性狀進行了遺傳多樣性分析;王吉明等[8]基于果實性狀對從國外引進的100份野生甜瓜種質資源的遺傳多樣性進行了研究;閆洪朗等[9]對江浙滬地區的65個甜瓜品種果實性狀進行了遺傳多樣性分析。以上這些報道都是基于表型性狀進行甜瓜遺傳多樣性研究。
分子標記技術是評價種質資源遺傳多樣性的重要手段。目前已有利用分子標記研究甜瓜遺傳多樣性的報道,所用標記主要涉及AFLP、SRAP、RAPD、SSR以及ISSR等[10-14]。SNP標記作為目前遺傳學研究中最為流行的分子標記,其與傳統分子標記相比具有在基因組中分布廣泛、密度高、遺傳穩定以及易于分析等優勢。2b-RAD是一種基于IIB型限制性核酸內切酶的簡化基因組測序技術,也是近年來在非模式生物中進行SNP開發最經濟有效的測序方法之一,具有酶切DNA片段大小一致、文庫構建簡單快速、標簽密度易于調節、重復性和準確性高以及成本低廉等優點[15]。該技術已在果蠅、扇貝、線蟲、對蝦、擬南芥、水稻、玉米和茶樹等物種的基因型分型、高精度連鎖圖譜構建、群體遺傳結構和遺傳多樣性分析、QTL(quantitative trait locus)定位以及系統演化分析等研究中得到廣泛應用[16-22]。迄今尚未有利用2b-RAD技術對甜瓜種質開展遺傳多樣性研究的相關報道。
厚皮甜瓜是區別于薄皮甜瓜的一個類型,包括厚薄皮雜交類型甜瓜、網紋厚皮甜瓜和哈密瓜品種,具有耐低溫弱光、耐濕以及抗病的特點[13]。因其經濟效益比較可觀,在上海具有較廣的種植面積。本研究以從國內外搜集和自主選育的在果實性狀方面(包括果皮顏色、果面網紋和果肉顏色)差異比較大的28份高代穩定自交系厚皮甜瓜為材料,利用2b-RAD技術對其在分子水平進行遺傳多樣性及親緣關系分析,以期為今后甜瓜SNP分子標記的開發和分子標記輔助選擇育種提供科學依據。
1 材料和方法
1.1 供試材料
28份供試甜瓜種質材料均由上海市農業科學院園藝研究所提供,其編號信息和果實主要性狀詳見表1。

表1 28份甜瓜種質及其果實主要性狀
1.2 方 法
1.2.1 基因組DNA提取和2b-RAD文庫構建采集苗期甜瓜葉片,10株混合取樣。液氮速凍后,置于-80 ℃冰箱保存備用。基因組DNA采用CTAB法進行提取[23],通過瓊脂糖檢測和紫外可見分光光度計對抽提樣本基因組DNA的質量和濃度進行檢測。樣本DNA抽提質檢合格后,利用2b-RAD五標簽串聯技術進行測序文庫構建,采用標準型5′-NNN-3′接頭與BsaXI酶切標簽連接。文庫質控合格后在Illumina nova平臺進行Paired-end測序。2b-RAD測序由上海歐易生物醫學科技有限公司完成。
1.2.2 測序質量分析利用Illumina nova測序平臺得到原始測序序列后,使用Pear軟件(Version 0.9.6)對其進行拼接。為保證后續數據分析結果的準確性,將拼接好的序列按照以下條件進行過濾:過濾刪除含有N堿基比例大于8%的序列;過濾刪除低質量序列 (質量值低于Q30的堿基數超過整條序列堿基數15%)。依據建庫時各個樣本所在的位置,提取出各樣本對應的序列,過濾刪除不含酶切識別位點的序列后得到各樣本的高質量測序序列,即含酶切位點的序列。
1.2.3 全基因組范圍SNP篩查和分型分析對于有參照基因組的標記分型,依據 RAD-typing的分型策略[24],標記分型如下:(1)從甜瓜參考基因組序列(ftp://cucurbitgenomics.org/pub/cucurbit/ genome/ melon/v3.6.1/CM3.6.1_pseudomol.fa.gz)中提取包含BsaX I 酶切位點的標簽作為參考序列;(2)利用SOAP軟件[25]將各樣本的高質量序列比對到參考序列上(參數為-r 0-M 4 -v 2);(3)利用最大似然法(maximum likelihood, ML)進行SNP標記分型。為保證后續分析的準確性,分型完成后通過以下指標對分型結果進一步過濾:剔除只含有1種等位基因的位點;剔除基因組堿基為N的位點;剔除一個標簽內多于2個SNP的標簽;剔除同一位置2種分型的位點;剔除所有樣本中低于80%個體可以分型的位點;剔除最小等位基因頻率低于0.01的位點;剔除等位基因大于2的位點。
1.3 數據處理
分別利用SnpEff軟件(V4.1g)、Genepop軟件(V1.0.5)、Vcftools軟件(V0.1.14)、Admixture軟件(V1.3.0)和Treebest軟件(Version: 1.9.2)進行SNP注釋,遺傳分化系數(Fst)統計分析,SNP位點的觀測雜合度(Ho)、期望雜合度(He)、觀測等位基因數(Na)、有效等位基因數(Ne)、多態信息含量(PIC)和核苷酸多樣性(Pi)分析,群體結構分析以及進化樹分析。
2 結果與分析
2.1 2b-RAD測序結果分析
利用2b-RAD測序技術構建28份甜瓜種質的標簽測序文庫(1份種質即為1個樣本),結果(表2)顯示,測序共獲得391 918 500條原始序列,平均每個樣本的測序序列數為13 997 089。參照Wang等[15]的方法對原始序列進行質量控制,在28個樣本的原始序列中篩選出含有BsaX Ⅰ酶切位點的高質量序列比例均在82%以上(表2),說明利用此方法構建的甜瓜樣本測序文庫質量較好。

表2 2b-RAD測序數據基本分析
2.2 SNP分子標記在甜瓜中的分布
參照RADtyping分型策略[24],對比甜瓜參考基因組,根據比對結果統計樣本獲得的標簽數和測序深度信息。結果顯示,28個甜瓜樣本中比對到參考基因組上的標簽數在67 732~70 919之間,平均標簽數為69 684,測序深度值從21.8~128.36不等,平均測序深度為49×(表3)。從28個樣本的共同測序標簽中共篩選出10 318個高質量的SNP位點,包括9 988個定位于染色體上的SNP位點和330個非染色體SNP位點。從SNP位點在每條染色體上的數量可以看出,6號染色體上存在的SNP位點最多,為1 233個,其次是1號染色體,為1 162個(圖1,A)。這些SNP位點主要分布于基因間區以及基因的上下游序列(圖1,B)。

表3 28個甜瓜樣本的標簽數和測序深度
2.3 SNP 突變類型分析
SNP分型結果中各位點的突變類型具體如下:G/A轉換類型占所有堿基突變類型的33.94%,T/C轉換類型占33.35%;T/A和T/G顛換類型分別占所有堿基突變類型的 9.17%和8.80%,G/C和C/A顛換類型分別占5.96%和 8.78%。SNPs中發生轉換與發生顛換的比值(Ts/Tv)為 2.15。
2.4 28個甜瓜樣本的SNP遺傳多樣性分析
本次分型只統計了二等位基因型,因此每個SNP位點觀測等位基因數Na值均為2。SNP遺傳多樣性分析結果顯示,每個二等位SNP位點觀測雜合度Ho數值在0~1之間,平均值為0.068;期望雜合度He數值在0.035~0.500之間,平均值為0.291;有效等位基因數Ne數值在1.036~2.000之間,平均值為1.484;多態信息含量PIC數值在0.035~0.375之間,平均值為0.237;核苷酸多樣性Pi數值在0.036~0.511之間,平均值為0.297;最小等位基因頻率數值在0.018~0.500之間,平均值為0.214;位點的分型率數值在82.14%~100%之間,平均值為96.54%。

A. 各染色體SNPs的數量分布;B. SNPs在基因組中的分布圖1 SNPs 在甜瓜基因組中的分布情況A. Distribution of the number of SNPs in each chromosome; B. Distribution of SNPs in genomeFig.1 Distribution of SNPs in melon genome
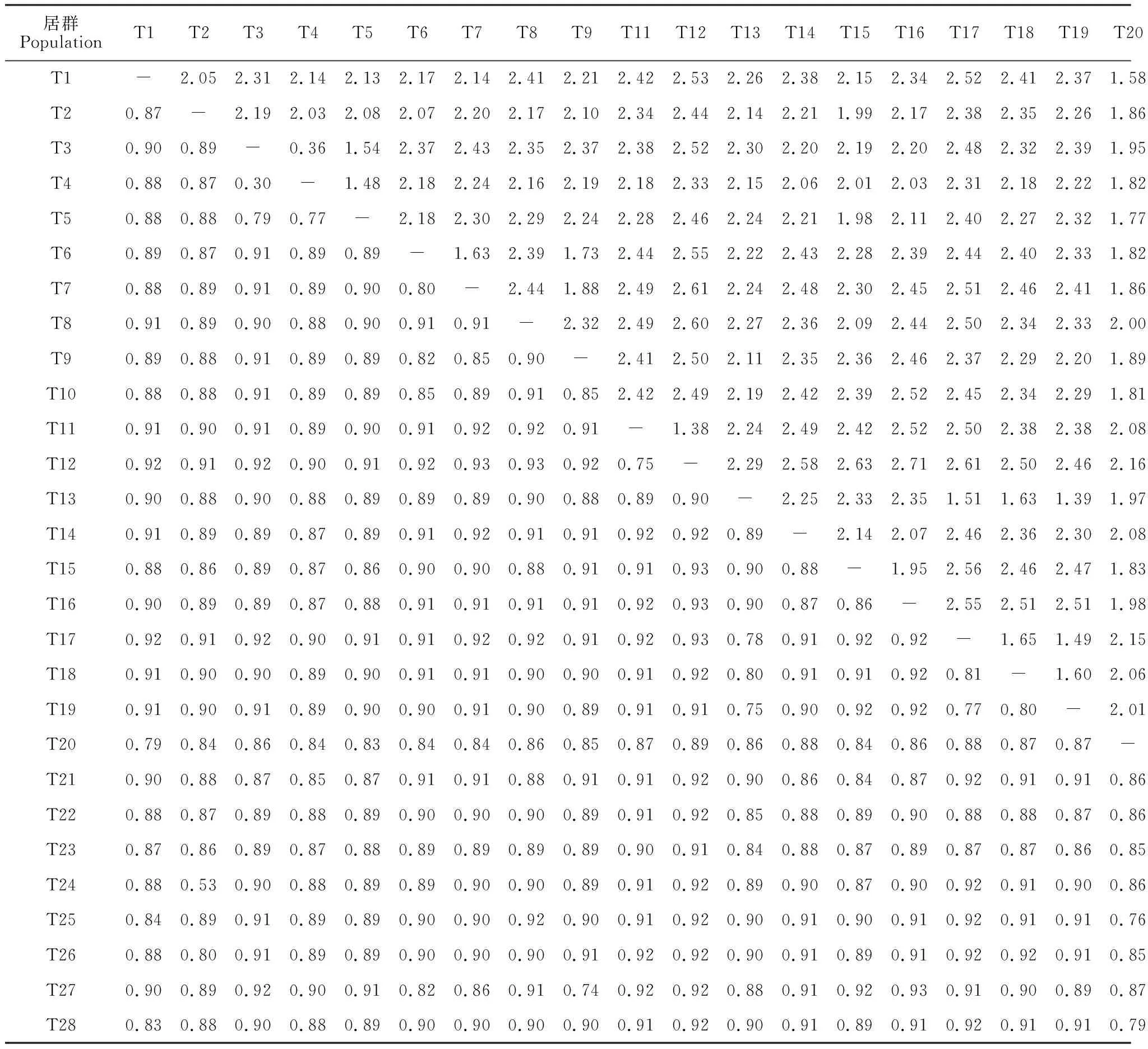
表4 28個甜瓜樣本遺傳分化系數(Fst, 下三角)和遺傳距離(DR, 上三角)
2.5 甜瓜樣本間遺傳變異及分化分析
遺傳分析結果(表4)顯示,28個甜瓜樣本兩兩之間的遺傳分化系數(Fst)在 0.188~0.933之間,平均值為0.885,遺傳距離(DR)范圍在0.208~2.706之間,平均值為2.218,表明各樣本之間均存在高度的遺傳分化(一般認為,若群體Fst值在0~0.05之間,表明其各亞群間不存在分化;若Fst值在0.05~0.15之間,為中度分化;若Fst值在0.15~0.25之間,則為高度分化);其中,T22和T23兩個樣本之間的遺傳分化系數和遺傳距離最小,T12和T16兩個樣本之間的遺傳分化系數和遺傳距離最大(表4)。
依據果皮顏色分類,28份甜瓜種質可分為4個群體,分別為白皮群體(包括T1、T6、T14、T22、T23和T28)、黃皮群體(包括T3、T4、T5、T11、T12、T16和T20)、青皮群體(包括T2、T7、T8、T9、T15、T21、T24和T26)和綠皮群體(包括T10、T13、T17、T18、T19、T25和T27)。遺傳分析結果(表5)顯示,這4個群體兩兩之間的Fst值從0.052~0.188不等,群體間遺傳距離DR值從0.054~0.208不等,其中白皮群體和青皮群體間的Fst值最小,黃皮群體和綠皮群體間的Fst值最大(表5)。各群體之間的Fst值均大于0.05,表明其均存在中度及以上程度的遺傳分化。
依據果面網紋性狀分類,28份甜瓜種質可分為3個群體,分別為光皮群體(包括T12、T14、T16、T21、T22和T23)、稀網群體(包括T1、T3、T4、T8、T11、T15和T26)和密網群體(包括T2、T5、T6、T7、T9、T10、T13、T17、T18、T19、T20、T24、T25、T27和T28)。遺傳分析結果(表6)顯示,這3個群體兩兩之間的Fst值分別為0.004、0.096和0.029,DR值分別為0.004、0.101和0.029,表明光皮群體和密網群體之間存在中度遺傳分化,光皮群體和稀網群體之間以及稀網群體和密網群體之間無顯著分化。
依據果肉顏色分類,28份甜瓜種質可分為3個群體,分別為白肉群體(包括T4、T14、T16、T22和T23)、桔肉群體(包括T1、T2、T5、T6、T7、T9、T10、T11、T12、T17、T18、T20、T27和T28)以及綠肉群體(包括T3、T8、T13、T15、T19、T21、T24、T25和T26)。遺傳分析結果顯示,這3個群體兩兩之間的Fst值分別為0.126、0.033和0.027,DR值分別為0.134、0.033和0.027,表明白肉群體和桔肉群體之間存在中度遺傳分化,白肉群體和綠肉群體之間以及桔肉群體和綠肉群體之間無明顯分化(表7)。
綜合以上結果表明,28份甜瓜種質依據果皮顏色分類的各群體之間的遺傳分化系數總體上是最大的,即遺傳分化程度最高,并且各群體之間均存在中度及以上程度的遺傳分化。
2.6 群體遺傳結構分析
利用10 318個SNPs標記28份甜瓜種質,根據其果實性狀,再結合K值最大似然值分類,將這些種質劃分為3個類群(圖2)。3個類群分別用紅色、綠色和藍色代表,即類群1、類群2和類群3。由圖3可以看出,大部分條帶(每個條帶代表1份種質)都由2~3種顏色組成,表明3個類群大部分種質之間存在不同程度的基因交流。種質所屬類群,可根據顏色占條帶的比例推斷。

表5 基于果皮顏色分類的28個甜瓜樣本群體間的遺傳分化系數(Fst)和遺傳距離(DR)

表6 基于果面網紋性狀分類的28個甜瓜樣本群體間的遺傳分化系數(Fst)和遺傳距離(DR)

紅色代表類群1;綠色代表類群2;藍色代表類群3圖2 28份甜瓜種質SNPs分析群體結構(K=3)Red color blocks represent group 1; Green color blocks represent group 2; Blue color blocks represent group 3Fig.2 Analysis of population structure using identified SNPs for 28 melon germplasms (K=3)

表7 基于果肉顏色分類的28個甜瓜樣本群體間的遺傳分化系數(Fst)和遺傳距離(DR)
2.7 系統進化樹構建及聚類分析
從聚類樹形圖(圖3)可以看出,28份甜瓜種質被分為 3類,第一類包括T2、T3、T4、T5、T8、T14、T15、T16、T21、T24和T26共11份種質,第二類包括T1、T6、T7、T9、T10、T20、T25、T27和T28共9份種質,第三類包括T11、T12、T13、T17、T18、T19、T22和T23共8份種質。其中,第一類主要為自主選育出的高代自交系甜瓜種質,第二類主要為從新疆引進或從新疆品種中選育出來的高代自交系種質,第三類主要為從日本引進或從日本品種中選育出來的高代自交系種質。
3 討 論
豐富多樣的種質資源是選育高產、優質和高抗新品種以及研究物種起源、進化和分類的基礎。甜瓜作為葫蘆科植物中表型變異最為豐富的作物之一,育種工作者利用不同方法針對不同方向對其進行了廣泛研究[6-9, 26-30]。胡建斌等[7]研究發現甜瓜果實性狀的變異是甜瓜表型變異的主要來源。雖然已有研究人員利用不同的甜瓜種質資源對甜瓜果實表型性狀進行了遺傳多樣性分析[8-9, 31-32],但目前仍未見利用2b-RAD技術對其在分子水平上進行研究的相關報道。

居群編號同表1圖3 28份甜瓜種質基于SNP標記的聚類Population numbers are the same as those in Table 1Fig.3 Phylogenetic tree of 28 melon germplasms revealed by SNP markers
2b-RAD 技術利用IIB型限制性內切酶對基因組進行酶切后產生等長的33~36 bp的酶切標簽,經富集后用于下游的高通量測序反應,結合生物信息學分析在全基因組范圍內對SNP位點進行高通量篩選和基因分型分析,獲得的批量的 SNP 位點可為群體遺傳多樣性分析奠定良好的基礎[24, 33]。研究表明,該技術生物學重復的準確性高達99% 以上[34]。本研究經2b-RAD測序后,每個樣本得到的原始序列數從912 682~30 740 808不等,質量過濾后,平均每個樣本中獲得含有BsaXI 酶切位點的高質量序列數占測序原始序列數的82%以上,每個個體的測序深度值在21.8~128.36之間,平均測序深度為49×,達到了高準確度下的分型標準[35],表明本次構建的文庫質量較高,測序結果準確可靠。
SNP堿基替換類型分為轉換(Ts)與顛換(Tv)兩類,轉換發生在A與G或C與T之間,顛換發生在A 與C、A與T、G與C以及G與T之間。研究表明,不同物種間轉換與顛換的比值存在很大差異。砂梨(Pyruspyrifolia)轉錄組測序中轉換與顛換的比值為1.71[36];桑葚不同時期轉錄組測序中轉換與顛換的比值約為1.6[37];澳洲堅果(Macadamiaintegrifolia)重測序結果顯示SNPs轉換和顛換的比率為0.77[38];中國明對蝦(Fenneropenaeuschinensis)2b-RAD測序結果中轉換與顛換的比值為1.402[19];馬氏珠母貝(Pinctadafucata)血細胞轉錄組測序中轉換與顛換的比值為0.5[39]。本研究2b-RAD測序結果顯示SNPs轉換與顛換的比值(Ts/Tv)為2.15。產生這種差異的原因可能與不同物種在進化過程中承受的選擇壓力有關[40]。
多態信息含量(PIC)和雜合度(Ho)是2個較好的衡量基因多態性的指標[19]。當PIC< 0.25時,表示該位點為低度多態位點;PIC介于0.25~0.50之間時,表示該位點為中度多態性位點;PIC> 0.50時,表示該位點為高度多態性位點[41]。本研究中28個甜瓜樣本SNPs位點的PIC值在0.035~0.375之間,表明各SNP位點呈低度多態性或中度多態性。28個樣本SNPs位點的Ho值從0~1不等,表明各位點的雜合度存在較大差異。
種群的遺傳分化指數(Fst)是衡量群體間遺傳分化程度的重要參數。本研究中依據果皮顏色、果面網紋和果肉顏色3個性狀進行分類的各群體之間的Fst值分別從0.052~0.188、0.004~0.096以及0.027~0.126不等,表明依據果皮顏色分類的各群體之間遺傳分化程度最高,群體間的遺傳結構差異最為顯著,而依據果面網紋形成情況進行分類的各群體之間整體遺傳分化程度最低。
本研究以果皮顏色、果面網紋和果肉顏色3個性狀差異比較大的28份甜瓜種質為材料進行遺傳多樣性分析研究,共鑒定到10 318個SNP位點。后續可從以上位點中篩選可重復、易分辯、強專一的 SNP位點集合,用于構建甜瓜種質指紋圖譜、果實性狀相關的QTL定位分析以及種子純度鑒定研究,同時也可將其應用于甜瓜分子標記輔助選擇育種工作中以加快新品種的選育進程。
目前,Fergany M.等[42]通過形態學和SSR標記研究認為甜瓜親緣關系與其生態地理分布具有高度相關性;程振家等[10]通過AFLP標記研究認為厚皮甜瓜聚類結果除了與果面網紋性狀有關外,與生態地理來源也具有一定的相關性;徐志紅等[43]通過形態學分析認為厚皮甜瓜的聚類結果與成熟期有一定的相關性。本研究通過系統進化樹分析將28份厚皮甜瓜種質劃分為3類,其中第二類大部分為從新疆引進或從新疆品種中選育出來的純合種質,第三類大部分為從日本引進或從日本品種中選育出來的純合種質,說明本研究中甜瓜聚類結果與地理來源具有一定關系,這與程振家等[10]和Fergany M.等[42]的研究結果十分相似。
本研究中依據分子水平的聚類分析結果與育種者對育種材料的分類不完全一致。育種者主要是根據甜瓜果實的果皮顏色、果肉顏色和有無網紋等性狀對其進行分類,而這只是一種表型分類。由于參試的28份材料有些是由各種類型的甜瓜雜交后選育出的純合種質,這些種質可能含有多個品種的血緣。因此基于表型的分類不能真實地反映各材料間親緣關系的遠近。程振家等[10]的研究結果也表明基于分子水平的分類方法能夠更加準確地揭示材料之間的內在遺傳聯系。本研究結果也提示我們,在今后的甜瓜育種工作中要綜合材料的表型性狀和分子水平的分析結果,選擇不同地理來源、表型差異顯著、親緣關系遠的材料進行雜交,這樣才更容易培育出優質、高產和抗病的甜瓜新品種。
作者貢獻:張永平、曹燕燕、刁倩楠和陳幼源進行試驗設計;曹燕燕進行數據分析和論文寫作;張永平進行最后的論文修改。全體作者都閱讀并同意最終的文本。
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
