6個匍匐剪股穎種質對干旱脅迫的生理響應與耐旱性評價
2021-03-15 06:06:41劉南清林紹艷周興元
江蘇農業科學 2021年1期
劉南清 林紹艷 周興元

摘要:采用盆栽模擬干旱脅迫試驗的方法,研究5個野生匍匐剪股穎種質(AgrostisstoloniferaL.)生態型與1個進口品種在干旱脅迫期間的生理響應,測定與分析葉色(Color)、相對含水量(RWC)、最大光化學效率(Fv/Fm)、滲透調節(OA)、蒸散率(ER)、相對電導率(EL)、丙二醛(MDA)含量等指標的變化,并用隸屬函數法進行耐旱性綜合評價。結果表明,隨著干旱脅迫時間的增加,匍匐剪股穎的Color、RWC、Fv/Fm及ER持續降低,而葉片的EL、MDA含量則持續升高,這些生理指標在變化上存在顯著的生態型差異。干旱脅迫16d,匍匐剪股穎生態型A-LY和A-LS種質與生態型A-DY、A-HZ和A-HS種質之間生理指標對比發現,其中生態型A-LY和A-LS種質的指標Color、RWC、Fv/Fm和OA顯著低于生態型A-DY、A-HZ和A-HS種質;生態型A-LY和A-LS種質的指標EL和MDA顯著高于生態型A-DY、A-HZ和A-HS種質。采用隸屬函數法對6個匍匐剪股穎種質的耐旱性進行耐旱性綜合評價,耐旱性排序為A-HS>A-HZ>A-DY>Pen-A4>A-LY>A-LS。
關鍵詞:匍匐剪股穎種質;野生生態型;耐旱性;生理響應;隸屬函數綜合評價
中圖分類號:S688.401文獻標志碼:A
文章編號:1002-1302(2021)01-0118-08
作者簡介:劉南清(1979—),男,廣西平南人,博士,副教授,主要從事草坪生產與管理及草坪草育種研究。E-mail:Liunanqing@jsafc.edu.cn。
通信作者:周興元,博士,教授,主要從事林草植物逆境生理研究。E-mail:Zhouxingyuan@jsafc.edu.cn。
干旱是最為普遍的自然災害,我國地處東亞地區,是典型的季風氣候,季風年際的不穩定性導致我國干旱災害頻發,其發生頻次約占總氣候災害頻次的1/3,為各項災害之首[1]。干旱脅迫是植物生長過程中遭遇的最主要的環境抑制因子之一[2],草坪草尤其是冷季型草坪草因其耐旱性不強,因而容易受到干旱脅迫的不利影響[3]。
匍匐剪股穎(AgrostisstoloniferaL.)作為世界上最重要的冷季型草坪草之一,其原產于歐亞大陸。匍匐剪股穎具有耐寒、耐陰、耐低修剪等優點,并兼具柔軟細膩、色澤鮮綠、成坪品質高等優質的坪用性狀,是當今世界高爾夫球場果嶺草坪的最優秀的建坪草種[4]。然而,匍匐剪股穎的耐旱性較差,為獲得高質量的匍匐剪股穎草坪,在草坪養護管理過程中須要頻繁灌溉,消耗大量的淡水資源,養護成本高。
我國華東地區經濟發達,對草坪的質量標準要求高,匍匐剪股穎被廣泛應用于園林綠化及高爾夫球場的建造中。當前,我國匍匐剪股穎草種基本從國外進口,耐旱性與耐熱性較差,在華東地區因夏秋季干旱脅迫而萎蔫干枯嚴重,極大降低了草坪的使用價值與使用效果。我國華東地區有著豐富的野生匍匐剪股穎種質資源,對其進行耐旱性研究,明確其耐旱機制,并進行耐旱性綜合評價,對挖掘具有強耐旱性的野生匍匐剪股穎種質資源,選育與推廣耐旱型匍匐剪股穎新品種,降低草坪養護管理中水資源與能源的消耗,降低養護成本,提倡生態節約型草坪建植與養護理念,都具有重要的意義。
本試驗以5個采集于華東地區各省(市)的野生匍匐剪股穎生態型,以及國外引進的品種Pen-A4為材料,采用干旱處理,通過測定葉色(Color)、相對含水量(RWC)、最大光化學效率(Fv/Fm)、滲透調節(OA)、蒸散率(ER)、相對電導率(EL)和丙二醛含量(MDA)等指標,研究干旱脅迫下匍匐剪股穎的生理響應,運用模糊數學隸屬函數法綜合評價6個匍匐剪股穎的耐旱性,旨在明確匍匐剪股穎的耐旱機制,挖掘耐旱性強的種質資源,為選育耐旱型匍匐剪股穎鄉土草種提供依據。
1材料與方法
1.1試驗區概況
試驗地位于江蘇省句容市邊城鎮江蘇農博園草坪種質資源圃(32°01′34.76″N,119°14′25.85″E),海拔為50m。年均氣溫為15.5℃,全年無霜期為220~240d,土壤為黃棕壤,pH值為7.6,試驗地土壤肥力中等,地力均勻。
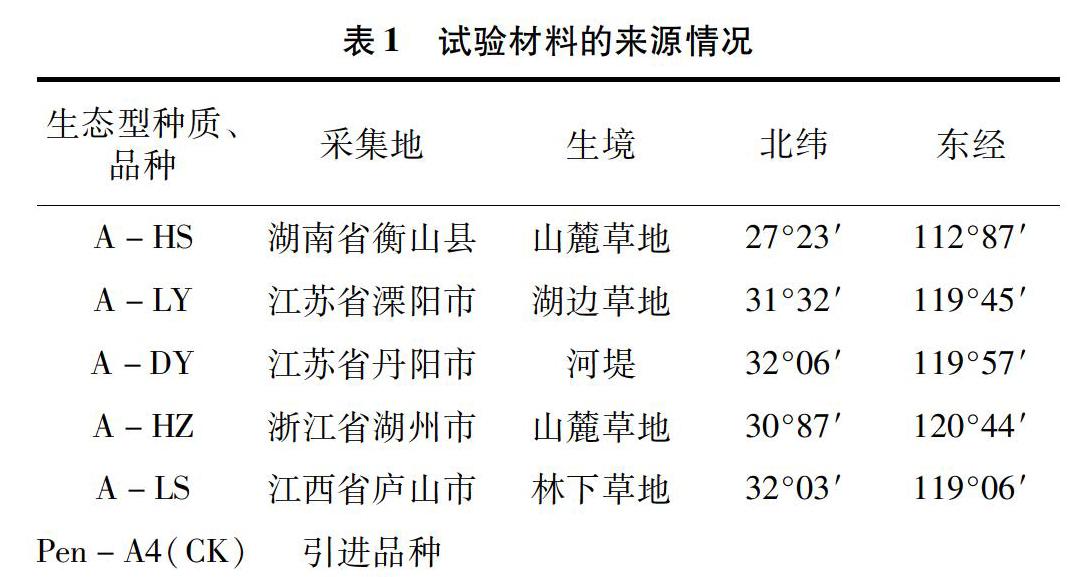
1.2試驗材料
供試材料共6份,2014年采集于華東各省的野生匍匐剪股穎種質資源5份,引進品種1份(表1)。供試材料采集或引進后,于2014年11月種植于種質資源圃,正常水肥管理,所有供試材料均完全成活并生長良好。
1.3試驗方法
2017年3月,從種質資源圃挖取6個匍匐剪股穎草塊,用水清洗干凈根部土壤,隨后立刻移栽到充滿純凈細河沙的聚氯乙烯(PVC)種植管中(深40cm,管徑為12cm),放進江蘇農博園的日光溫室中栽培,每份供試材料設置澆水對照組與干旱處理組,3次重復。3—5月期間,供試材料放置于溫室條件下(10~12h/d的自然光照,晝夜平均溫度分別為21、13℃)栽培3個月,期間正常水肥管理,每隔3d修剪1次,修剪高度為6cm。3個月后,把栽培好的試驗材料轉移進人工氣候模擬箱中進行適應性栽培。2017年8月6日,在人工氣候模擬箱中開始對植物進行干旱脅迫處理。生長室保持20/15℃的晝夜溫度,70%的相對濕度,光照時長為12h/d,以及650mmol/(m2·s)的草坪草冠際高度光照度。
干旱脅迫開始之前,每周給草坪草澆水3次,澆水時,當發現水分從種植管底部流出時即停止澆水,以確保足夠的土壤濕度。每周使用50%的霍格蘭氏全營養液進行施肥1次,每隔3d用剪刀對草坪進行修剪1次,保持6cm的修剪高度。試驗設計2個處理,分別是干旱處理組和對照組。干旱處理組采用自然干旱(即不澆水)的方法,分別于干旱處理0、4、8、12、16d進行各項指標測定,并剪取長勢一致的完全展開葉進行指標的分析測定。對照組(即正常澆水)保持每周3次的澆水頻率,確保根系土壤水分充足。
1.4觀測項目與生理指標的測定
1.4.1葉色
采用目測法測定匍匐剪股穎的葉片色澤,評分標準采用草坪草葉片色澤9級評分法。其中,1級代表分值最低,表示草坪草完全枯黃;9級代表分值最高,表示草坪草葉片含水量充足,色澤墨綠[5]。3次重復。
1.4.2相對含水量[JP3]
RWC計算公式為RWC=100×[(FW-DW)/(TW-DW)][6],FW為葉片的鮮質量;DW指樣品葉片在80℃的烘箱內烘72h后葉片的干質量;TW是把葉片完全浸泡在去離子水中,放置于4℃冰箱中,24h后吸飽水后的質量。3次生物學重復。
1.4.3相對電導率
采用李合生的相對電導率測定方法[7]。取樣后將葉片用去離子水沖洗3次,并用潔凈濾紙吸干表面水分。葉片剪成0.5cm的小片段,準確稱取0.5g,裝入具塞試管。加30mL去離子水,真空抽氣10min,振蕩,加塞,在室溫下靜置1h。3次生物學重復。用電導率儀測得電導率(E1),沸水浴10min,冷卻后測得電導率(E2)。計算公式:EL=E1/E2×100%。
1.4.4滲透調節
通過完全復水法來測定OA,把失水的葉片完全浸泡于水中,使葉片完全吸飽水復水,用壓汁器壓出細胞液,測定葉片飽和滲透壓[8]。把吸飽水后的葉片樣品放置于液氮中低溫急凍處理,然后儲存于-20℃的低溫冰箱中。測定葉片飽和滲透壓時,從低溫冰箱中取出葉片樣品,用壓汁器(FredS.Carver,Wabash,IN)壓出葉片細胞液,用與滲透壓儀配套的專用濾紙吸足細胞液,把吸足了細胞液的濾紙放在滲透壓測定儀(Vapromodel5520;Wescor,Logan,UT)上測定葉片細胞的滲透壓(C,單位為mmol/kg)。用公式1MPa=-C·2.58×10-3把單位mmol/kg換算成MPa。測定的澆水對照的葉片滲透壓與干旱脅迫處理的葉片的滲透壓之差,即為滲透調節OA。3次重復。
1.4.5最大光化學效率
采用葉綠素熒光儀(Fim1500型,美國德克薩斯州休斯敦Dynamax公司生產)測定法測定光化學效率。測定時,用葉綠素熒光測定儀的葉片夾分別夾住各處理的匍匐剪股穎的葉片,關閉葉片夾的透光孔,避光30min處理,讀取Fv/Fm測定值[9],3次重復。
1.4.6蒸散率
蒸散率的測定采用質量平衡法進行測定,每隔2d稱量1次不同處理的總質量,即不同生態型的匍匐剪股穎的總生物量與其種植柱的質量,計算出2次稱量的質量差值,用此質量差值作為蒸散量,蒸散率為單位面積及單位時間內的蒸散量[10]。3次重復。
1.4.7丙二醛含量
MDA含量采用硫代巴比妥酸法測定[11]。剪取葉片鮮樣0.5~1.0g,在-80℃的液氮中冷凍。解凍后,加7mL濃度為50mmol/L的磷酸鹽緩沖溶液(pH值為7.0),并立即在冰浴中研磨。以上所得提取液放置于4℃于20000g高速離心25min。待所得的上清液與三氯乙酸和硫代巴比妥酸充分反應后,用分光光度計(SpectronicInstruments,NewYork)在532nm與600nm波長下分別測定其吸光度。使用Dhindsa的計算公式[12]計算出MDA的含量。3次重復。
1.5數據處理
不同處理間的方差分析采用SAS軟件(9.0版)進行。用Fisher檢驗法確定處理間差異的顯著性,P<0.05表示差異顯著。
采用公式X(μ)=(X-Xmin)/(Xmax-Xmin)計算模糊數學隸屬函數,公式中的X(μ)為匍匐剪股穎某一耐旱性相關指標的隸屬函數值;X為該指標的平均值;Xmax和Xmin分別為該指標的最大值、最小值。最后將匍匐剪股穎各生態型所有性狀的具體隸屬值加和,加和平均值即代表各生態型各自的耐旱性綜合值[13]。
2結果與分析
2.1葉色與葉片相對含水量的變化
2.1.1葉色的變化
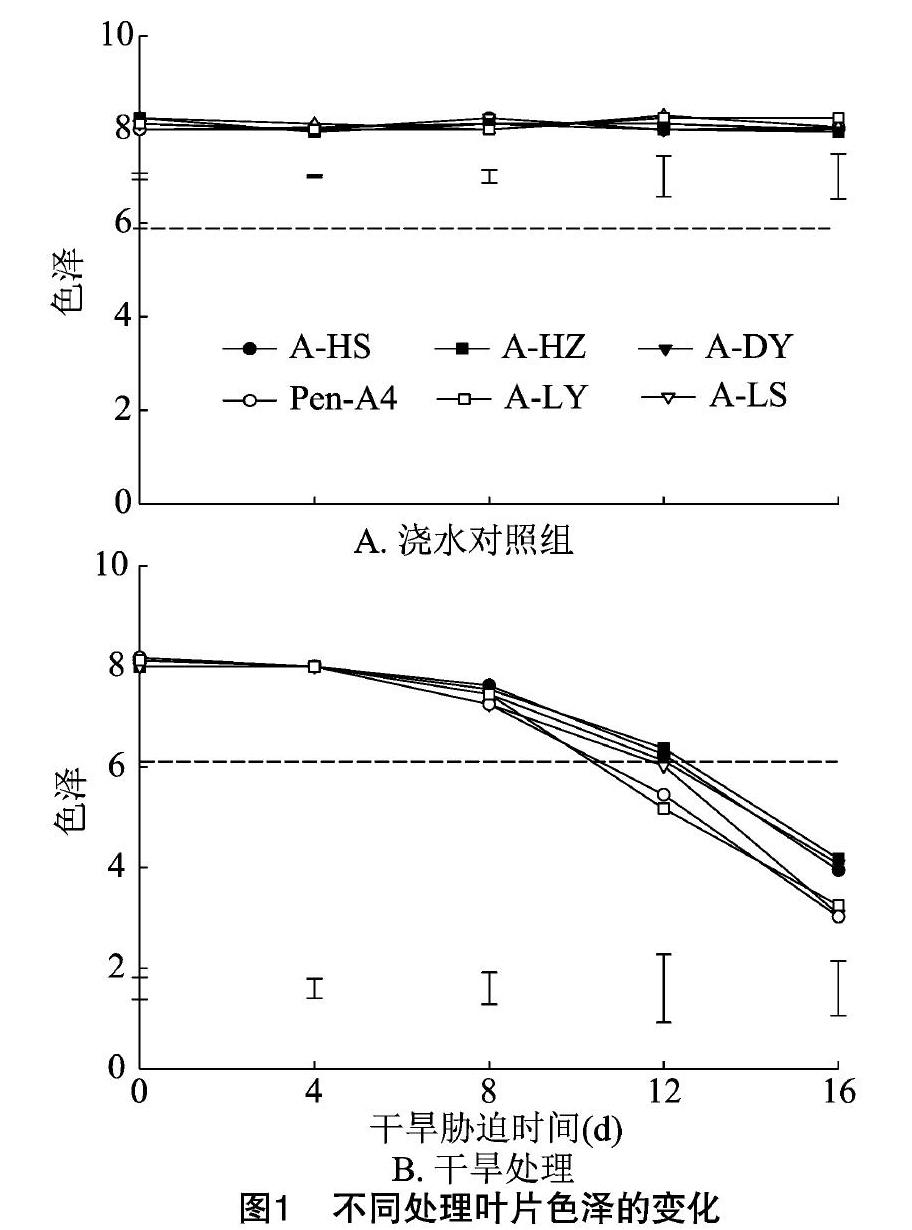
隨干旱處理時間的延長,澆水對照組匍匐剪股穎的葉色都保持在7.95~8.30的較高水平,不同生態型種質間無明顯差異(圖1-a)。
在干旱處理的0~4d,各生態型匍匐剪股穎葉色降低不明顯;干旱脅迫至8d時,匍匐剪股穎的葉色開始出現明顯的下降,各生態型間的下降幅度差異明顯;干旱脅迫12d時,6個匍匐剪股穎葉色都急劇下降,葉色的降低程度在不同生態型間差異明顯,野生生態型A-LY與Pen-A4種質的葉色明顯低于其他生態型,降低到6.0以下;干旱脅迫16d,A-HS、A-HZ、A-DY、Pen-A4、A-LY和A-LS種質的葉色分別下降47.81%、51.38%、49.85%、61.56%、63.00%、60.00%;在干旱脅迫16d后,A-LY與A-LS種質的葉色一直明顯低于A-HS、A-HZ、A-DY種質(圖1-b)。
2.1.2葉片相對含水量的變化
澆水對照組在整個試驗期間葉片的相對含水量維持在85%~94%之間,不同生態型種質和品種間無明顯差異(圖2-a)。
干旱脅迫開始的0~4d,匍匐剪股穎葉片的相對含水量下降不明顯;干旱脅迫4d至試驗結束,匍匐剪股穎各生態型與品種的葉片相對含水量持續下降,在下降速度與下降幅度上,不同生態型種質與品種間差異明顯;干旱脅迫16d時,A-HS、A-HZ、A-DY、Pen-A4、A-LY和A-LS種質相對含水量分別下降了46.95%、49.87%、50.75%、55.38%、60.70%、62.39%;野生生態型種質A-HS、A-HZ和A-DY的葉片相對含水量明顯高于A-LY、A-LS與Pen-A4種質(圖2-b)。
2.2葉片相對電導率與丙二醛含量的變化
2.2.1葉片相對電導率的變化
干旱脅迫期間,澆水對照組匍匐剪股穎的葉片相對電導率維持在較低的水平,不同生態型種質間和品種間差異不明顯(圖3-a)。
隨著干旱脅迫加重,匍匐剪股穎各生態型和品種的葉片相對電導率持續上升,但是在升高速度與程度上各生態型和品種間差異明顯;干旱脅迫16d時,A-HS、A-HZ、A-DY、Pen-A4、A-LY和A-LS種質的葉片相對電導率分別為0d的3.21、2.98、3.24、3.22、3.85、4.83倍;干旱8d至試驗結束,匍匐剪股穎野生生態型種質A-LY、A-LS與Pen-A4的葉片相對電導率都明顯高于A-DY、A-HZ與A-HS種質(圖3-b)。
2.2.2葉片丙二醛含量的變化
澆水對照組匍匐剪股穎葉片的MDA含量較低,不同生態型種質與品種間差異不明顯(圖4-a)。
干旱脅迫期間,匍匐剪股穎葉片的MDA含量持續增加,但是不同生態
型和品種在MDA含量的增加幅度上差異明顯;干旱脅迫16d時,A-HS、A-HZ、A-DY、Pen-A4、A-LY和A-LS種質葉片的MDA含量分別是初始的3.76、4.39、5.21、4.83、4.41、5.22倍;匍匐剪股穎野生生態型種質A-LY與A-LS的葉片MDA含量明顯高于Pen-A4、A-DY、A-HZ與A-HS種質(圖4-b)。
2.3葉片滲透壓的變化
干旱脅迫期間,澆水對照組所有匍匐剪股穎葉片的OA穩定保持在0.150~0.242MPa之間,不同生態型種質與品種間差異不明顯(圖5-a)。
干旱脅迫發生時,6個匍匐剪股穎葉片內的滲透調節都表現出先輕微下降,后快速升高,再快速下降的趨勢,在12~16d的干旱脅迫期間,野生生態型種質A-HS、A-HZ與A-DY的OA都明顯大于品種Pen-A4與A-LY、A-LS種質(圖5-b)。
2.4蒸散率的變化
在澆水對照組中,匍匐剪股穎的蒸散率存在明顯的生態型和品種間差異。野生生態型種質A-LY、A-LS、A-DY與品種Pen-A4的蒸散率較高,而A-HS與A-HZ種質的蒸散率則相對較低(圖6-a)。
干旱脅迫過程中,6個匍匐剪股穎的蒸散率持續降低,在10~12、14~16d,品種Pen-A4的蒸散率明顯高于其他5個匍匐剪股穎野生生態型種質之外;干旱脅迫18~20d,A-HS、A-HZ、A-DY、
Pen-A4、A-LY和A-LS種質蒸散率分別下降至2~4d蒸散率的8.56%、9.13%、8.72%、7.98%、6.22%、6.75%;在其他時期內,匍匐剪股穎的蒸散率并未表現出生態型和品種間的差異(圖6-b)。
2.5最大光化學效率的變化
干旱脅迫期間,澆水對照組6個匍匐剪股穎種質的最大光化學效率保持在0.76~0.80的高水平,不同生態型種質和品種間無明顯差異(圖7-a)。
干旱脅迫下,匍匐剪股穎的最大光化學效率持續下降。在0~8d下降速度緩慢,6個匍匐剪股穎種質間無明顯差異。在干旱脅迫的8~16d,最大光化學效率急劇下降,試驗結束時,最大光化學效率最低的生態型種質A-LS降到0.35;干旱脅迫16d后,A-HS、A-HZ、A-DY、Pen-A4、A-LY和A-LS種質的最大光化學效率分別下降了38.43%、38.92%、41.31%、41.66%、54.89%、56.87%;匍匐剪股穎各生態型種質和品種間最大光化學效率差異明顯,A-HS、A-HZ、A-DY種質和對照品種Pen-A4的最大光化學效率均顯著高于A-LY與A-LS種質(圖7-b)。
2.6干旱脅迫期間耐旱生理指標的隸屬函數法分析
對5個匍匐剪股穎野生生態型種質與品種Pen-A4的葉色、最大光化學效率、相對含水量、滲透調節、相對電導率與丙二醛含量等指標進行隸屬函數法計算綜合耐旱性,結果表明,5個匍匐剪股穎種質野生生態型與品種Pen-A4間的綜合耐旱性差異明顯,綜合耐旱性排序為A-HS>A-HZ>A-DY>Pen-A4>A-LY>A-LS(表2)。
2.7干旱脅迫期間生理指標相關性分析
相關性分析結果(表3)表明,干旱脅迫下,綜合耐旱性X(μ)與Color、Fv/Fm、RWC及OA之間存在顯著或極顯著正相關,相關系數分別為0.70、0.96、0.95、0.82;X(μ)與EL及MDA之間存在極顯著的負相關,相關系數分別為-0.87、-0.94;X(μ)與ER之間呈負相關性,但是相關性不顯著。
3討論與結論
在逆境脅迫發生時,植物細胞膜系統的通透性與膜脂過氧化程度會增加,植物受到逆境脅迫的傷害。在干旱脅迫下,草坪草細胞膜系統的通透性與膜脂過氧化程度升高,膜系統加速解體,草坪草遭受干旱脅迫傷害[14-16]。當前,相對電導率與丙二醛含量作為衡量細胞膜通透性與膜脂過氧化程度的重要指標而被廣泛接受[17]。干旱脅迫下,草地早熟禾(PoapratensisL.)[18-20]、高羊茅(FestucaarundinaceaSchreb.)[21-24]、黑麥草(LoliumperenneL.)[24-25]與匍匐剪股穎[26]等草坪草的相對電導率與丙二醛含量都持續升高。光合作用在調節植物耐旱性方面起著至關重要的作用。相關研究表明,當受到干旱脅迫時,草坪草的葉綠素含量快速降低,最大光化學效率與光合作用也因而大幅度降低,草坪草在干旱脅迫下的存活率降低[27-29]。
保持適宜的葉片水分狀態對維持葉片的正常生理生化功能尤為重要。當干旱脅迫發生時,能在較長時間內維持葉片適宜的相對含水量的植物,保持其正常生理代謝的可能性更大,存活率也更高,耐旱性更強。滲透調節被認為是諸多植物的耐旱生理機制[30],在草坪草中同樣如此[31-32]。在缺水狀態下,提高滲透調節有利于維持細胞膨壓[33-35]。相關對匍匐剪股穎的耐旱研究發現,干旱脅迫下,一方面,匍匐剪股穎因缺水加劇,相對含水量下降,另一方面,細胞內的多糖、脯氨酸、甜菜堿、多胺等滲透調節物質在干旱脅迫時合成增加,從而維持細胞的滲透調節,降低干旱脅迫傷害[15-16]。研究發現,草坪草植物的葉片蒸散率與草坪草植物的水分利用率具有密切相關性,蒸散率因物種或品種的不同而存在明顯的差異[36-37]。相關研究認為,低蒸散率是草坪草節水、耐旱的一個重要性狀[38]。然而,相關研究發現,匍匐剪股穎不同品種間的蒸散率僅在水分充足時存在明顯差異,而在干旱脅迫下,則品種間的蒸散率差異不明顯[39-40]。
本試驗中,干旱脅迫發生時,6個匍匐剪股穎種質的葉色、相對含水量、最大光化學效率與蒸散率都持續降低,相對電導率與丙二醛含量則出現持續升高,表明6個匍匐剪股穎種質的外部形態與內部細胞都受到了干旱脅迫的傷害。這可能由于干旱脅迫使得匍匐剪股穎無法從缺水的根系土壤獲取足夠的水分,植物體內的細胞含水量下降,植株相對含水量降低,細胞膜的流動性降低,膜透性和膜脂過氧化程度均持續上升,因而導致細胞膜系統與細胞解體的加速,伴隨著膜系統與細胞的解體,匍匐剪股穎葉片細胞內的葉綠體開始大量解體,葉色快速失綠,葉片最大光化學效率急劇降低。本研究發現,滲透調節在干旱脅迫期間出現了先升后降的變化趨勢,在干旱脅迫發生8d后,6個匍匐剪股穎種質的滲透調節同時升高,野生生態型A-HS、A-HZ與A-DY的滲透調節持續升高至干旱脅迫發生12d時。這可能是匍匐剪股穎適應干旱脅迫的一種機制,通過提高滲透調節,阻止水分由細胞內快速流失,保持一定的水合能力,降低干旱傷害。
本研究發現,在干旱脅迫下,匍匐剪股穎的葉色、相對含水量、最大光化學效率、滲透調節、蒸散率、相對電導率、丙二醛含量等7個生理指標的變化存在明顯的生態型或品種間的差異。通過隸屬函數法對6個匍匐剪股穎種質進行耐旱性綜合評價發現,6個匍匐剪股穎種質的耐旱性存在明顯的生態型與品種間的差異,綜合耐旱性與相關生理指標的相關性分析結果表明,匍匐剪股穎的綜合耐旱性與葉色、最大光化學效率、相對含水量、滲透調節等存在顯著或極顯著的正相關關系,而與相對電導率及丙二醛含量存在極顯著的負相關關系,但與蒸散率的相關性不明顯[13],這些指標可以作為衡量匍匐剪股穎耐旱性的生理指標。
6個匍匐剪股穎種質的綜合耐旱性存在差異,排序為A-HS>A-HZ>A-DY>Pen-A4>A-LY>A-LS。說明在野生匍匐剪股穎種質中存在著比引進品種耐旱性更高的生態型,匍匐剪股穎種質生態型A-HS、A-HZ與A-DY對選育強耐旱型匍匐剪股穎具有較高的價值,可以作為匍匐剪股穎耐旱種質育種的材料。
參考文獻:
[1]黃會平.1949—2007年全國干旱災害特征、成因及減災對策[J].干旱區資源與環境,2010,24(11):94-98.
[2]曲濤,南志標.作物和牧草對干旱脅迫的響應及機理研究進展[J].草業學報,2008,17(2):126-135.
[3]馬祎,王彩云.幾種引進冷季型草坪草的生長及抗旱生理指標[J].草業科學,2001,18(2):57-61.
[4]TurgeonAJ.TurfgrassManagemen[M].UpperSddleRiver,NJ:PrenticeHall,2008:84-86.
[5]TurgeonAJ.TurfgrassManagemen[M].4thed.Englewood:PrenticeHall,1996:207-211.
[6]BarrsHD,WeatherleyPE.Are-examinationoftherelativeturgiditytechniqueforestimatingwaterdeficitsinleaves[J].AustralianJournalofBiologicalSciences,1962,15(3):413-428.
[7]李合生.植物生理生化實驗原理和技術[M].北京:高等教育出版社,2000.
[8]BüssisD,HeinekeD.Acclimationofpotatoplantstopolyethyleneglycol-inducedwaterdeficit:Ⅱ.Contentsandsubcellulardistributionoforganicsolutes[J].JournalofExperimentalBotany,1998,49(325):1361-1370.
[9]MerewitzEB,GianfagnaT,HuangB.Proteinaccumulationinleavesandrootsassociatedwithimproveddroughttoleranceincreepingbentgrassexpressinganiptgeneforcytokininsynthesis[J].JournalofExperimentalBotany,2011,62(15):5311-5333.
[10]MccannSE,HuangB.Evaluationofdroughttoleranceandavoidancetraitsforsixcreepingbentgrasscultivars[J].HorticulturalScience,2008,43(2):519-524.
[11]劉祖祺,張石城.植物抗性生理學[M].北京:中國農業出版社,1994:372.
[12]DhindsaRS.Inhibitionofproteinsynthesisbyproductsoflipidperoxidation[J].Phytochemistry,1982,21(2):309-313.
[13]李俊慶,芮文利,齊敏忠,等.水分脅迫對不同抗旱型花生生長發育及生理特性的影響[J].中國農業氣象,1996(1):11-13.
[14]AbrahamEM,HuangBR,BonosSA,etal.EvaluationofdroughtresistanceforTexasbluegrass,Kentuckybluegrass,andtheirhybrids[J].CropScience,2004,44(5):1746-1753.
[15]LiuNQ,ShenYX,HuangBR.Osmoregulantsinvolvedinosmoticadjustmentfordifferentialdroughttoleranceindifferentbentgrassgenotypes[J].JournaloftheAmericanSocietyforHorticulturalScience,2015,140(6):605-613.
[16]LiuNQ,LinSY,HuangBR.Differentialeffectsofglycinebetaineandspermidineonosmoticadjustmentandantioxidantdefensecontributingtoimproveddroughttoleranceincreepingbentgrass[J].JournaloftheAmericanSocietyforHorticulturalScience,2017,142(1):20-26.
[17]RachmilevitchS,DacostaM,HuangBR.Physiologicalandbiochemicalindicatorsforstresstolerance[M].BocaRaton:CrcPress,2006.
[18]李壽田,韓建國,毛培勝.26個草地早熟禾品種苗期抗旱性綜合評價[J].草業科學,2012,29(7):1114-1119.
[19]白利國,俞玲,馬暉玲.野生草地早熟禾對干旱脅迫的生理響應[J].草原與草坪,2014,34(2):86-91.
[20]王昕慧,孫曉陽,李勛,等.鉀素增強干旱脅迫下草地早熟禾生理耐性的機制[J].中國草地學報,2018,40(4):23-29.
[21]HuangBR,FryJD.Rootanatomical,physiological,andmorphologicalresponsestodroughtstressfortallfescuecultivars[J].CropScience,1998,38(4):1017-1022.
[22]DuHM,WangZL,HuangBR.Differentialresponsesoftallfescueandzoysiagrasstoheatanddroughtstress[J].ActaHorticulturae,2008,783:207-213.
[23]謝曉蓉,李唯,劉金榮,等.21個高羊茅品種抗旱抗熱性的生理評價[J].草地學報,2009,17(2):202-205.
[24]楊有俊,鄭明珠,秦偉志,等.干旱及復水處理下坪用黑麥草和高羊茅抗旱特性比較[J].草業科學,2012,29(3):370-376.
[25]OraeeA,TehranifarA,NezamiA,etal.Effectsofdroughtstressoncoldhardinessofnon-acclimatedviola(Viola×wittrockiana‘IonaGoldwithBlotch)incontrolledconditions[J].ScientiaHorticulturae,2018,238:98-106.
[26]MaXQ,ZhangJ,BurgessP,etal.Interactiveeffectsofmelatoninandcytokininonalleviatingdrought-inducedleafsenescenceincreepingbentgrass(Agrostisstolonifera)[J].EnvironmentalandExperimentalBotany,2018,145:1-11.
[27]劉美君,隋曉青,安沙舟.干旱脫水對狗牙根葉片光合機構的影響[J].草地學報,2018,26(2):441-446.
[28]常紅軍,秦毓茜.干旱和鹽脅迫對草地早熟禾草坪質量及其葉綠素熒光參數的影響[J].西北植物學報,2008,28(9):1850-1855.
[29]杜建雄,師尚禮,劉金榮,等.干旱脅迫和復水對草地早熟禾3個品種生理特性的影響[J].草地學報,2010,18(1):73-77.
[30]BohnertHJ,JensenRG.Strategiesforengineeringwater-stresstoleranceinplants[J].TrendsinBiotechnology,1996,14(3):89-97.
[31]QianYL,FryJD,UphamWS.Rootinganddroughtavoidanceofwarm-seasonturfgrassesandtallfescueinKansas[J].CropScience,1997,37(3):905-910.
[32]DacostaM,HuangBR.Changesincarbonpartitioningandaccumulationpatternsduringdroughtandrecoveryforcolonialbentgrass,creepingbentgrass,andvelvetbentgrass[J].JournaloftheAmericanSocietyforHorticulturalScience,2006,131(4):789-802.
[33]HuangBR,FryJD.Osmoticpotential,sucroselevel,andactivityofsucrosemetabolicenzymesintallfescueinresponsetodeficitirrigation[J].JournaloftheAmericanSocietyforHorticulturalScience,2010,135(6):505-510.
[34]BurgessP,HuangBR.Effectsofsequentialapplicationofplantgrowthregulatorsandosmoregulantsondroughttoleranceofcreepingbentgrass[J].CropScience,2013,54(2):837-844.
[35]黃鑫,楊健,羅新寧.調虧灌溉對棉花生長及滲透調節物質的影響[J].江蘇農業科學,2020,48(7):96-101.
[36]郭君,向佐湘.五種草坪草的蒸散量研究[J].作物研究,2009,23(3):197-200.
[37]潘全山,韓建國,王培.五個草地早熟禾品種蒸散量及節水性[J].草地學報,2001,9(3):207-212.
[38]AlvesAAC,SetterTL.Responseofcassavatowaterdeficit:leafareagrowthandabscisicacid[J].CropScience,2000,40(1):131-137.
[39]McCannSE,HuangBR.Evaluationofdroughttoleranceandavoidancetraitsforsixcreepingbentgrasscultivars[J].HortScience,2008,43(2):519-524.
[40]MccannSE,HuangBR.Effectsoftrinexapac-ethylfoliarapplicationoncreepingbentgrassresponsestocombineddroughtandheatstress[J].CropScience,2007,47(5):2121-2128.
