基于生物信息學(xué)方法構(gòu)建類風(fēng)濕性關(guān)節(jié)炎miRNA-mRNA調(diào)控網(wǎng)絡(luò)*
2021-03-17 00:58:00曹維維袁成良
國(guó)際檢驗(yàn)醫(yī)學(xué)雜志 2021年5期
曹維維,袁成良
德陽(yáng)市人民醫(yī)院檢驗(yàn)科,四川德陽(yáng) 618000
類風(fēng)濕性關(guān)節(jié)炎是一種累積多個(gè)系統(tǒng)的常見(jiàn)自身免疫性疾病,其主要臨床表現(xiàn)為慢性進(jìn)行性的滑膜炎癥,對(duì)關(guān)節(jié)軟骨和骨質(zhì)造成破壞最終導(dǎo)致關(guān)節(jié)功能障礙。全球患病率為0.5%~1.0%[1]。類風(fēng)濕性關(guān)節(jié)炎的發(fā)病機(jī)制涉及多種細(xì)胞類型,其中位于滑膜關(guān)節(jié)的成纖維樣滑膜細(xì)胞在炎癥、骨質(zhì)破壞及血管翳的形成等病理過(guò)程中發(fā)揮重要作用[2]。研究發(fā)現(xiàn),類風(fēng)濕性關(guān)節(jié)炎外周血及炎癥組織中的微小RNA(miRNA)存在異常表達(dá),并與關(guān)節(jié)炎癥的發(fā)生和發(fā)展、滑膜組織的增生以及滑膜細(xì)胞對(duì)凋亡作用的耐受性等病理過(guò)程密切相關(guān)[3]。本研究基于生物信息學(xué)方法構(gòu)建miRNA-mRNA調(diào)控網(wǎng)絡(luò),為明確miRNA在類風(fēng)濕性關(guān)節(jié)炎中的分子調(diào)控機(jī)制提供新的思路。
1 材料與方法
1.1材料 本研究采用美國(guó)國(guó)立生物技術(shù)信息中心(NCBI)的基因表達(dá)綜合數(shù)據(jù)庫(kù)(GEO數(shù)據(jù)庫(kù))獲得類風(fēng)濕性關(guān)節(jié)炎的基因表達(dá)譜數(shù)據(jù)。其中GSE72564包含4例類風(fēng)濕性關(guān)節(jié)炎患者和4例骨關(guān)節(jié)炎滑膜樣本的miRNA表達(dá)數(shù)據(jù)。GSE55235包含10例類風(fēng)濕性關(guān)節(jié)炎患者和10例骨關(guān)節(jié)炎滑膜樣本的mRNA表達(dá)數(shù)據(jù)。
1.2差異miRNA和差異基因的篩選 本研究將樣本分為類風(fēng)濕性關(guān)節(jié)炎組和骨關(guān)節(jié)炎組,應(yīng)用GEO數(shù)據(jù)庫(kù)中的GEO2R在線分析工具,利用t檢驗(yàn)進(jìn)行多重比較,Benjamini & Hochberg 矯正P值,設(shè)定篩選條件為|logFC|>1且P<0.05,其中FC為差異倍數(shù)(FC)。篩選得到差異表達(dá)的miRNA和差異表達(dá)基因。
1.3靶基因預(yù)測(cè) miRNet(https://www.mirnet.ca/miRNet/upload/MirUploadView.xhtml)是一個(gè)同時(shí)收錄miRTarBase v8.0,TarBase v8.0和miRecords及miRanda等多個(gè)數(shù)據(jù)庫(kù)信息,具有綜合分析功能的在線生物信息預(yù)測(cè)數(shù)據(jù)庫(kù)。本研究采用miRNet在線數(shù)據(jù)庫(kù)進(jìn)行miRNA下游靶基因預(yù)測(cè)分析。
1.4功能和通路富集分析 miRNA預(yù)測(cè)的靶基因與差異表達(dá)基因取得交集,使用R語(yǔ)言中的clusterProfiler包對(duì)預(yù)測(cè)得到的靶基因與差異表達(dá)基因交集中的基因進(jìn)行基因本體論(GO)功能富集分析及京都基因與基因組百科全書(shū)(KEGG)通路富集分析,閾值P<0.05。
1.5miRNA-mRNA調(diào)控網(wǎng)絡(luò)構(gòu)建和cytohubba分析 通過(guò)綜合分析得到miRNA-mRNA關(guān)系對(duì)。利用可視化軟件Cytoscape構(gòu)建miRNA-mRNA調(diào)控網(wǎng)絡(luò)圖,并使用cytohubba插件進(jìn)行核心基因及miRNA的篩選。
2 結(jié) 果
2.1差異表達(dá)miRNA和差異表達(dá)基因的篩選 使用GEO2R在線分析工具對(duì)miRNA和mRNA基因表達(dá)譜芯片GSE72564和GSE55235進(jìn)行分析,共篩選出差異miRNA23個(gè),其中上調(diào)miRNA6個(gè),下調(diào)miRNA17個(gè)。共篩選到差異基因1 038個(gè),其中643個(gè)上調(diào)基因,395個(gè)下調(diào)基因。使用ggplot包繪制火山圖(圖1):差異表達(dá)miRNA火山圖(圖1A)和差異表達(dá)基因火山圖(圖1B)。
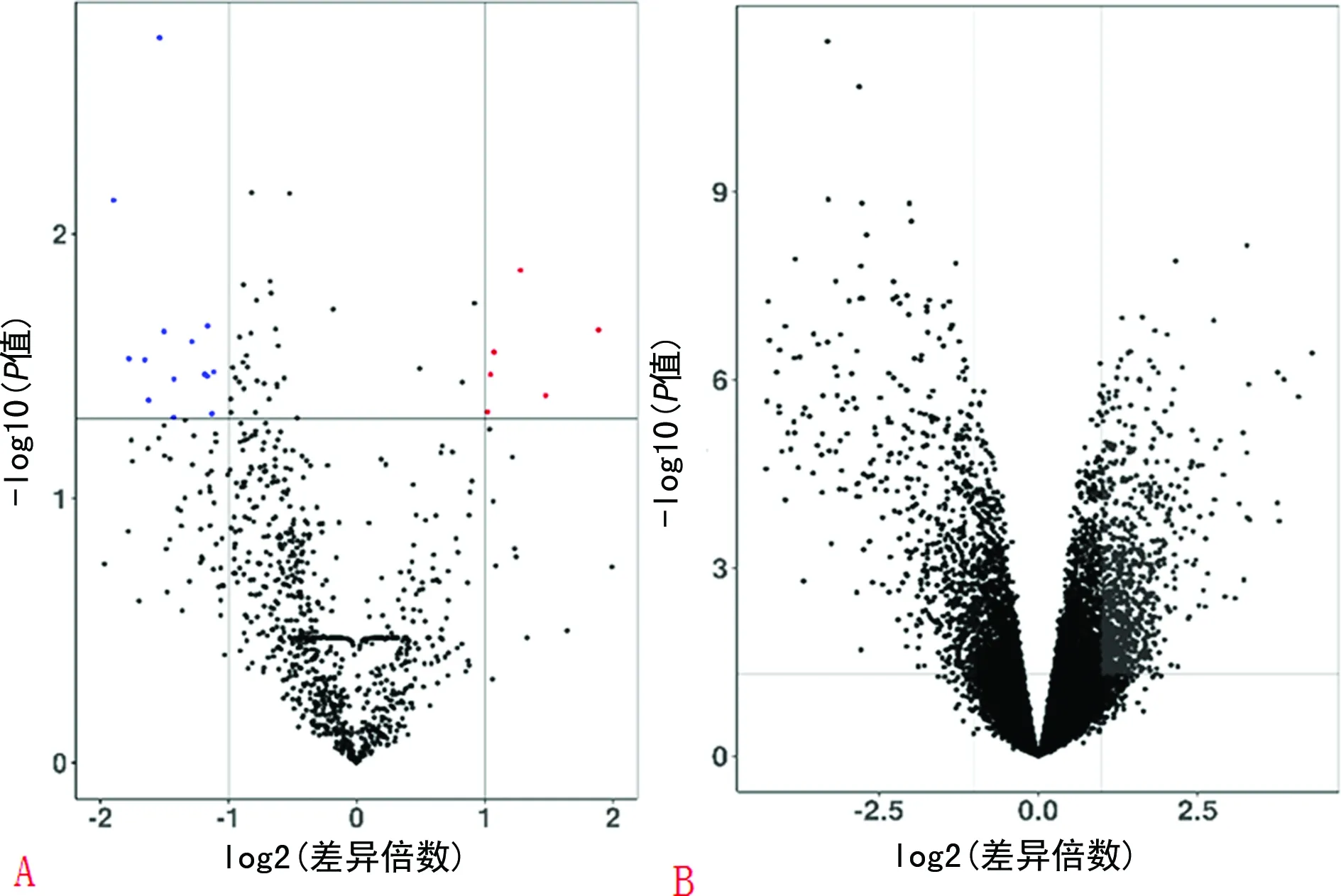
注:A為差異表達(dá)miRNA篩選;B為差異表達(dá)基因篩選。
2.2靶基因預(yù)測(cè) 分別選取下調(diào)倍數(shù)最顯著的前6個(gè)miRNA:hsa-miR-30c-2-3p、hsa-miR-1305、hsa-miR-2116-5p、hsa-miR-26a-1-3p、hsa-miR-708-5p、hsa-miR-218-5p和上調(diào)的6個(gè)miRNA:hsa-miR-2276-3p、hsa-miR-1193、hsa-miR-653-5p、hsa-miR-496、hsa-miR-4263、hsa-miR-346,使用miRNet在線預(yù)測(cè)工具,對(duì)篩選出的差異miRNA進(jìn)行可能的靶基因預(yù)測(cè)分析,得到2 044個(gè)預(yù)測(cè)靶基因。2 044個(gè)預(yù)測(cè)靶基因與1 038個(gè)差異基因取交集得到142個(gè)靶基因。
2.3KEGG和GO分析 GO分析:在BP(生物過(guò)程)方面,主要富集與細(xì)胞對(duì)氧含量降低的反應(yīng)、缺氧反應(yīng)、肌細(xì)胞分化、細(xì)胞生長(zhǎng)調(diào)節(jié)、上皮細(xì)胞增殖、白細(xì)胞分化等生物過(guò)程。在細(xì)胞組成方面主要富集與染色外基質(zhì)、染色質(zhì)、轉(zhuǎn)錄因子復(fù)合體、細(xì)胞質(zhì)膜及其蛋白復(fù)合物、受體復(fù)合物等細(xì)胞成分中。而分子功能方面主要富集與轉(zhuǎn)錄輔助因子活性、氨基多糖結(jié)合、染色質(zhì)結(jié)合、細(xì)胞黏附分子結(jié)合、蛋白異源二聚化活性等分子功能。KEGG分析:主要富集與磷脂酰肌醇3-激酶/蛋白激酶B(PI3K/AKT)信號(hào)通路、人類T細(xì)胞白血病病毒Ⅰ型感染、癌癥中的轉(zhuǎn)錄失調(diào)、EB病毒感染、乙型肝炎、庫(kù)欣綜合征、Th17細(xì)胞分化、Th1/Th2細(xì)胞分化等信號(hào)通路。
2.4miRNA-mRNA調(diào)控網(wǎng)絡(luò)的構(gòu)建和cytohubba分析 使用cytoscape(v3.8.0)可視化軟件繪制miRNA-mRNA的調(diào)控網(wǎng)絡(luò)圖,見(jiàn)圖2。三角形節(jié)點(diǎn)表示miRNA,矩形節(jié)點(diǎn)代表mRNA。圖2中可見(jiàn)一個(gè)靶基因可受多個(gè)miRNA調(diào)控,而1個(gè)miRNA則可調(diào)控多個(gè)靶基因。其中hsa-miR-218-5p調(diào)控的靶基因最多且以上調(diào)為主,其次為hsa-miR-30c-2-3p,同樣以上調(diào)為主。使用cytohubba插件,通過(guò)最大團(tuán)中心性MCC算法篩選出前十位的hub基因:PPP2R1B、MGAT4C、TUBB2A、ZNF124、CAPZB、HMGA1、HNRNPA1、HSPG2、KLHL21、TOP2A、WARS、CRISPLD2。
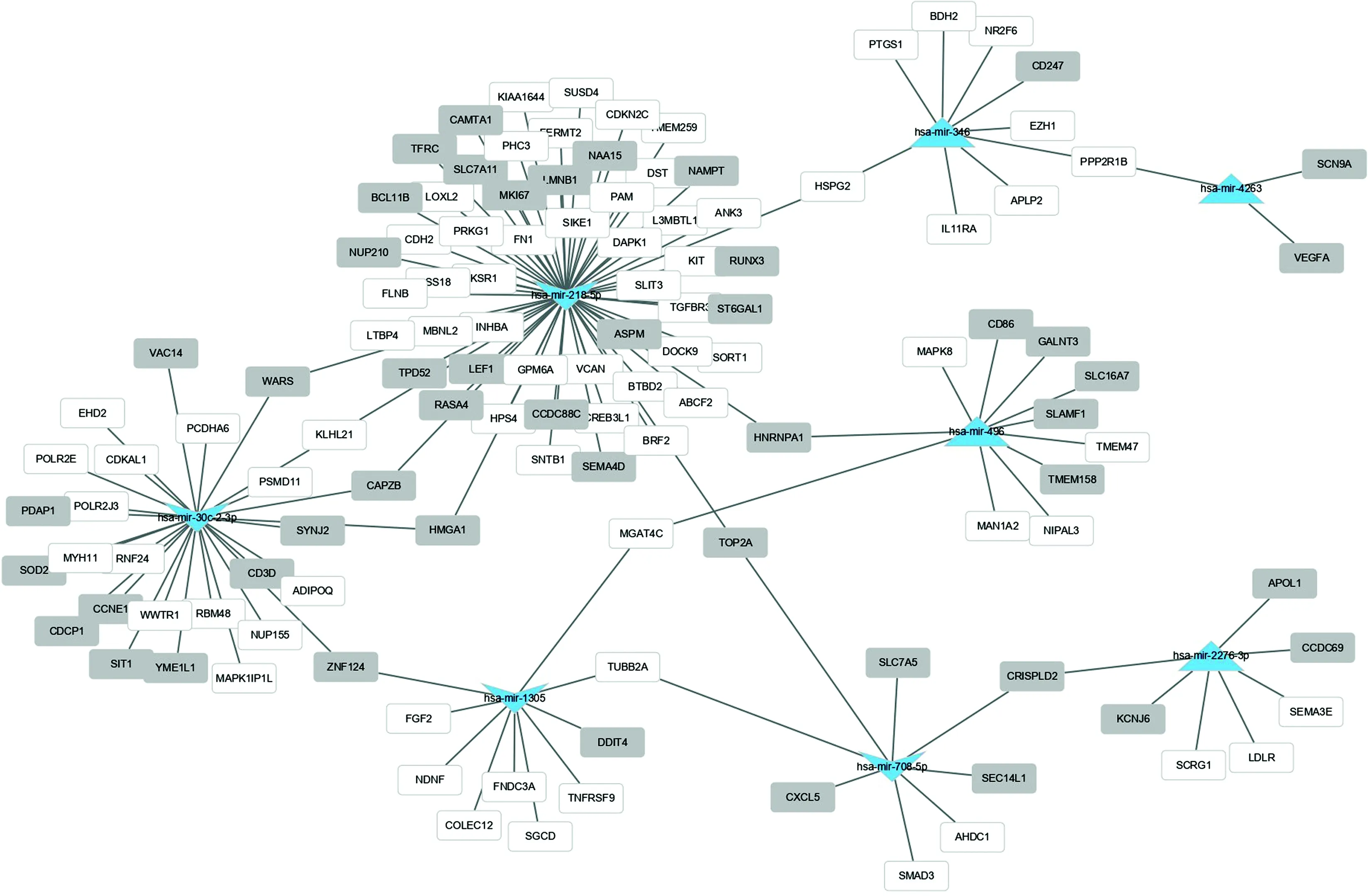
注:■為下調(diào)的基因;□為上調(diào)的基因;▲為上調(diào)的miRNA;▼為下調(diào)的miRNA。
3 討 論
類風(fēng)濕性關(guān)節(jié)炎作為一種累積多個(gè)系統(tǒng)的常見(jiàn)自身免疫性疾病。研究發(fā)現(xiàn)遺傳易感性、病毒感染、環(huán)境及生活方式等是類風(fēng)濕性關(guān)節(jié)炎目前已知的危險(xiǎn)致病因素[4-5]。miRNA在類風(fēng)濕性關(guān)節(jié)炎患者的血液、關(guān)節(jié)、滑膜和不同細(xì)胞類型中均存在差異表達(dá),而這些差異表達(dá)的miRNA可通過(guò)增強(qiáng)或抑制不同細(xì)胞的增殖分化、遷移和侵襲及血管新生等病理過(guò)程參與介導(dǎo)疾病的發(fā)生和發(fā)展。本研究以miRNA的作用機(jī)制為基礎(chǔ),通過(guò)生物信息分析方法成功構(gòu)建miRNA-mRNA調(diào)控網(wǎng)絡(luò),進(jìn)一步探討miRNA及相關(guān)信號(hào)通路在類風(fēng)濕性關(guān)節(jié)炎發(fā)生和發(fā)展中的分子調(diào)控機(jī)制,為類風(fēng)濕性關(guān)節(jié)炎的靶向診斷和治療提供可靠的理論依據(jù)。
利用GEO數(shù)據(jù)庫(kù)中的GSE72564和GSE55235,篩選與類風(fēng)濕性關(guān)節(jié)炎相關(guān)的差異表達(dá)的miRNA和mRNA,共篩選出差異miRNA23個(gè)(上調(diào)miRNA6個(gè)和下調(diào)miRNA17個(gè))及差異表達(dá)基因1 038個(gè)(643個(gè)上調(diào)基因和395個(gè)下調(diào)基因)。通過(guò)對(duì)預(yù)測(cè)靶基因與差異表達(dá)基因進(jìn)行整合,篩選出142個(gè)候選基因。進(jìn)行KEGG通路富集和GO功能富集分析,發(fā)現(xiàn)其主要富集于缺氧反應(yīng)及細(xì)胞黏附分子結(jié)合等生物過(guò)程和分子功能,參與調(diào)控PI3K/AKT信號(hào)通路、Th17細(xì)胞分化、Th1/Th2細(xì)胞分化等信號(hào)通路。
類風(fēng)濕性關(guān)節(jié)炎的滑膜組織炎癥及成纖維樣滑膜細(xì)胞(FLS)的大量增殖和分化使耗氧量增加,導(dǎo)致局部缺氧。缺氧誘導(dǎo)因子1α(HIF-1α)作為缺氧誘導(dǎo)因子HIF家族成員之一,對(duì)氧水平敏感并在缺氧條件下顯著增加。低氧環(huán)境可以使HIF-1α的積累,驅(qū)動(dòng)血管內(nèi)皮生長(zhǎng)因子(VEGF)及多種促血管生成介質(zhì)如CXCL8、CCR20的表達(dá)[6-7]。還可以增加促炎因子如白細(xì)胞介素(IL)-6、金屬基質(zhì)蛋白酶的分泌及增加滑膜細(xì)胞的侵襲性[8]。同時(shí)研究證實(shí),多種細(xì)胞黏附分子與類風(fēng)濕性關(guān)節(jié)炎的發(fā)生和發(fā)展密切相關(guān)[9-10]。MELINTE等[11]發(fā)現(xiàn),類風(fēng)濕性關(guān)節(jié)炎患者的滑膜軟骨組織中含有大量的血小板內(nèi)皮細(xì)胞黏附分子1(PECAM-1/CD31)。CD31作為免疫球蛋白超家族的成員,位于內(nèi)皮細(xì)胞的細(xì)胞間邊界上,在炎癥、細(xì)胞凋亡及白細(xì)胞的跨內(nèi)皮遷移中起到關(guān)鍵作用[12]。此外,研究者通過(guò)類風(fēng)濕性關(guān)節(jié)炎患者的FLS的共培養(yǎng)實(shí)驗(yàn)結(jié)果顯示,鈣黏蛋白11在FLS細(xì)胞間的接觸點(diǎn)上表達(dá),與α-連環(huán)蛋白和β-連環(huán)蛋白共定位形成黏附連接[13],進(jìn)而證明了鈣黏蛋白11在FLS細(xì)胞間的黏附作用。
PI3K/AKT信號(hào)通路作為一種重要的細(xì)胞內(nèi)信號(hào)轉(zhuǎn)導(dǎo)通路,已被證實(shí)與類風(fēng)濕性關(guān)節(jié)炎的發(fā)生發(fā)展有關(guān)[14]。有研究表明,類風(fēng)濕性關(guān)節(jié)炎患者FLS中的PI3K/AKT信號(hào)通路可以通過(guò)刺激細(xì)胞因子如IL-17、IL-22等引起FLS細(xì)胞的異常增殖并加重滑膜炎性反應(yīng)[15]。此外,PI3K/AKT信號(hào)通路可通過(guò)刺激Th17細(xì)胞分泌IL-17使FLS表達(dá)過(guò)量的核因子κB受體活化因子配體(RANKL),促進(jìn)破骨細(xì)胞的分化和形成從而導(dǎo)致骨質(zhì)破壞。值得一提的是,Th17細(xì)胞及幼稚CD4+T細(xì)胞的分化一定程度上也與PI3K/AKT信號(hào)通路的調(diào)節(jié)有關(guān)[16]。
Th17作為CD4+T細(xì)胞的亞群,可分泌IL-17A、IL-17F和IL-21等多種細(xì)胞因子,在類風(fēng)濕性關(guān)節(jié)炎患者外周血、關(guān)節(jié)滑液和滑膜組織中廣泛表達(dá)并參與促進(jìn)炎性反應(yīng)和軟骨破壞等多種病理過(guò)程。研究表明類風(fēng)濕性關(guān)節(jié)炎患者存在Th1/Th2失衡,Th1亞群占優(yōu)勢(shì),Th1通過(guò)分泌IL-2和γ-干擾素等細(xì)胞因子參與細(xì)胞免疫,Th2通過(guò)分泌IL-4/IL-10等細(xì)胞因子參與體液免疫[17]。Th1的異常增加導(dǎo)致促炎細(xì)胞因子的產(chǎn)生,加重炎癥和組織損傷的發(fā)生[18-20]。
本研究利用人類基因表達(dá)芯片,采用差異基因分析和靶基因預(yù)測(cè)等生物信息分析方法,構(gòu)建了miRNA-mRNA網(wǎng)絡(luò)。hsa-miR-218-5p、has-miR-30c-3p、hsa-miR-1305、hsa-miR-708-5p表達(dá)下調(diào),而hsa-miR-496、hsa-miR-346、hsa-miR-2276-3p、hsa-miR-4263表達(dá)為上調(diào)。研究發(fā)現(xiàn),hsa-miR-218-5p是成骨分化的重要誘導(dǎo)因子,其可通過(guò)靶向結(jié)合分泌型糖蛋白Slit及其跨膜受體Robo蛋白(Slit-Robo)途徑的跨膜受體蛋白1(Robo1)并抑制Dickkopf-1(DKK-1)的分泌從而促進(jìn)類風(fēng)濕性關(guān)節(jié)炎的FLS成骨分化[21]。長(zhǎng)鏈非編碼RNA DANCR在滑膜間充質(zhì)干細(xì)胞軟骨形成過(guò)程中起關(guān)鍵作用,hsa-miR-1305作為的DANCR的下游靶點(diǎn),其過(guò)度表達(dá)可以通過(guò)降低轉(zhuǎn)化生長(zhǎng)因子-β通路成員Smad4的表達(dá)進(jìn)而抑制hsa-miR-1305的表達(dá)而誘導(dǎo)軟骨細(xì)胞的增殖和分化作用[22]。hsa-miR-708-5p可通過(guò)抑制Wnt3a蛋白/細(xì)胞內(nèi)β-連環(huán)蛋白(Wnt3a/β-catenin)通路在蛋白和轉(zhuǎn)錄水平的活性促進(jìn)細(xì)胞凋亡并抑制細(xì)胞增殖從而參與類風(fēng)濕性關(guān)節(jié)炎的疾病調(diào)節(jié)機(jī)制[23]。SEMAAN等[24]發(fā)現(xiàn),hsa-miR-346可利用RNA結(jié)合蛋白穩(wěn)定作用控制類風(fēng)濕性關(guān)節(jié)炎中腫瘤壞死因子-α蛋白的釋放及其mRNA的穩(wěn)定性。
在Hub基因中,KLHL21編碼一組高度保守的蛋白,涉及多種細(xì)胞和分子并廣泛參與到炎性反應(yīng)、氧化應(yīng)激反應(yīng)等病理過(guò)程。研究證實(shí),類風(fēng)濕性關(guān)節(jié)炎患者滑膜巨噬細(xì)胞中的KLHL21表達(dá)上調(diào)[25]。MEI等[26]發(fā)現(xiàn),KLHL21可靶向作用于核轉(zhuǎn)錄因子-κB(NF-κB)抑制蛋白激酶β(IKKβ),對(duì)腫瘤壞死因子-α(TNF-α)激活的NF-κB信號(hào)通路產(chǎn)生負(fù)性調(diào)節(jié)作用,其過(guò)表達(dá)可以誘導(dǎo)細(xì)胞因子IL-8、IL-1β等多種促炎因子的釋放致使炎癥加重。硫酸肝素蛋白多糖2可以編碼一種大的多結(jié)構(gòu)域蛋白Perlecan,作為血管外基質(zhì)的重要組成部分,它有助于維持內(nèi)皮屏障功能,也是一種有效的平滑肌細(xì)胞增殖抑制劑,有助于維持血管穩(wěn)態(tài)。Perlecan可以和多種細(xì)胞表面受體,如整合素、細(xì)胞外基質(zhì)分子和生長(zhǎng)因子如成纖維細(xì)胞生長(zhǎng)因子、內(nèi)皮生長(zhǎng)因子、血小板衍生生長(zhǎng)因子等相互作用,從而在介導(dǎo)細(xì)胞的遷移、增殖分化等過(guò)程中發(fā)揮重要作用[27-28]。hsa-miR-218-5p作為高頻下調(diào)表達(dá)的miRNA,可同時(shí)靶向作用于KLHL21和 HSPG2的表達(dá)并下調(diào)二者水平。因此該miRNA-mRNA調(diào)控網(wǎng)絡(luò)與類風(fēng)濕性關(guān)節(jié)炎的發(fā)病機(jī)制間重要關(guān)系可作為新的研究方向進(jìn)行深入探究。
綜上,本研究通過(guò)差異基因篩選及靶基因預(yù)測(cè)等生物信息學(xué)分析方法,成功構(gòu)建了miRNA-mRNA調(diào)控網(wǎng)絡(luò)并進(jìn)行信號(hào)通路及功能富集分析和揭示了與類風(fēng)濕性關(guān)節(jié)炎相關(guān)的關(guān)鍵基因。這些關(guān)鍵基因在類風(fēng)濕性關(guān)節(jié)炎的發(fā)生發(fā)展過(guò)程中起到重要作用,為后續(xù)深入類風(fēng)濕性關(guān)節(jié)炎的診斷和治療提供了新的診斷靶點(diǎn)和治療思路。
猜你喜歡
音樂(lè)探索(2022年2期)2022-05-30 21:01:37
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國(guó)特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
電子制作(2018年18期)2018-11-14 01:48:24
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
鄭州大學(xué)學(xué)報(bào)(醫(yī)學(xué)版)(2015年2期)2015-02-27 14:50:46
中國(guó)中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
山東女子學(xué)院學(xué)報(bào)(2014年6期)2014-03-01 02:24:55
