蒙古櫟次生林林木競爭對不同撫育間伐方式的響應
2021-03-19 02:14:44胡雪凡段光爽張會儒張曉紅
林業科學研究 2021年1期
胡雪凡,段光爽,張會儒,4*,盧 軍,張曉紅
(1.中國林業科學研究院資源信息研究所,北京 100091;2.國家林業和草原局森林經營與生長模擬重點實驗室,北京 100091;3.信陽師范學院數學與統計學院,河南 信陽 464000;4.國家林業和草原局森林經營工程技術研究中心,北京 100091)
競爭是生物間相互作用的重要方面之一[1],可分為種內和種間競爭,對林木個體的生長發育、森林群落的結構和群落演替進程都有影響。林木競爭狀況多通過林木競爭指數來反映,競爭指數對林木之間的競爭進行了定量描述,在形式上反映了林木個體生長與生存空間的關系,實質上反映了樹木對環境資源需求及其在獲取環境資源時要承受的競爭壓力[2]。競爭隨著林齡的增加以及經營措施和環境的改變而發生改變[3],目前國內學者對種內和種間競爭的研究主要體現在優勢樹種種內和種間競爭現狀的分析[1,4-6],有少量關于群落種內和種間競爭隨時間變化的研究[7],但對林分內的競爭關系尤其是種內和種間競爭對經營措施的響應研究較少。研究林分的競爭關系及其對經營措施的響應對維護森林生態系統的穩定,更好實現森林的各項功能,科學的經營和管理森林至關重要[8]。
國內目標樹經營相關的研究多集中在對林分生長、結構、更新、生物多樣性等的影響[9-12],對林木競爭的影響研究較少。蒙古櫟(Quercus mongolicaFisch.ex Ledeb)次生林在我國東北地區分布范圍很廣,多為闊葉紅松(Pinus koraiensisSieb.et Zucc.)林過伐后形成的處于次生演替階段的森林群落,林地生產力較低且生態功能退化,如何通過經營來改善其林分狀況,提高其經濟價值和生態效益有很重要的意義[13]。
本研究以蒙古次生林為對象,分析了林木競爭關系以及不同撫育間伐方式對林分內競爭狀態的影響差異。為了解蒙古櫟次生林內部競爭關系及撫育間伐方式對競爭關系的影響效應和機制提供依據。
1 研究區概況
研究區位于吉林省汪清林業局塔子溝林場,地理坐標為129°97′~130°22′ E,43°32′~43°49′ N。林區地貌以低山丘陵為主,土壤以暗棕壤為主。氣候為溫帶大陸性季風氣候,年均溫3.9 ℃左右,1 月份平均氣溫最低,約為?32 ℃;7 月份平均氣溫最高,約為22 ℃,年均降水量為600~700 mm,多集中在夏季[12]。
研究區主要喬木樹種有蒙古櫟、紅松、白樺(Betula platyphyllaSuk.)、大青楊(Populus ussuriensisKom.)、糠椴(Tilia mandshuricaRup et Maxim.)、春榆(Ulmus davidianaPlanch.var.japonica(Rehd.)Nakai)、色木槭(Acer pictumThunb.ex Murray)、長白落葉松(Larix olgensisHenry)、黑樺(Betula dahuricaPall.)、水曲柳(Fraxinus mandshuricaRupr.)、臭冷杉(Abies nephrolepis(Trautv.)Maxim.)、魚鱗云杉(Picea jezoensisCarr.var.microsperma(Lindl.) Cheng et L.K.Fu)、黃檗(Phellodendron amurenseRupr.)等[12]。
2 材料與方法
2.1 數據來源
2013 年在塔子溝林場選擇具有代表性的蒙古櫟次生林林分(林齡40~70 a),設置12 塊面積為1 hm2(100 m×100 m)的固定監測樣地。調查樣地內胸徑5 cm 及以上的林木,測量并記錄坐標、樹種、胸徑、樹高等信息。對12 塊樣地采取完全隨機區組設計,各組林木胸徑、蓄積及斷面積無顯著差異。試驗設置4 種處理,3 次重復。4 種處理包括:(1)傳統經營,即T1 處理(1、7、9 號樣地),遵循《森林撫育規程》(GB/T15781—2009)的規定進行撫育間伐設計,采伐強度控制在25%以下,采伐對象主要是生長不良的林木,伐后郁閉度在0.6 以上;(2)目標樹經營密度1,即T2 處理(2、4、11 號樣地),按照目標樹單株木集約經營理念制定采伐設計,用材目標樹選取密度約為70 株·hm?2,目標樹的樹種選擇上優先考慮鄉土的、價值比較高的以及此類群落演替的頂級樹種。另外考慮了林木生命力、質量和距離3 種因素,將影響目標樹生長的林木確定為采伐木;(3)目標樹經營密度2,即T3 處理(3、8、10 號樣地),用材目標樹選取密度為100 株·hm?2,其余措施與T2 一致。(4)對照樣地,即T0 處理(5,6,12 號樣地)。為便于對比試驗,T0 和T1 按照100 株·hm?2選取了目標樹。2018 年9 月對所有樣地進行了復測,撫育經營前樣地信息見表1。
樣地邊緣林木的競爭木可能處于邊界之外,因此本研究將樣地邊界5 m 內區域視為緩沖區,其中林木只作競爭木來參與計算,核心區(90 m×90 m)的林木既作為競爭木又作為對象木參與計算。
2.2 研究方法
2.2.1 優勢樹種確定 采用優勢度分析法來確定優勢樹種[4,14]。具體方法是計算蒙古櫟次生林喬木樹種的相對胸高斷面積,作為優勢度,然后通過公式(1)確定優勢樹種個數。

式中:d表示確定優勢樹種個數的閾值,N為總樹種數,T為優勢度排位在前的樹種,即上位種,U為優勢度排位在后的樹種,及剩余種xi為上位種(T)的優勢度,x為優勢樹種所占的理想百分比,xj為剩余種的優勢度。如果喬木層只有一個優勢樹種,那么理想百分比為100%;如果有兩個優勢樹種,則理想百分比為50%;如果有3 個優勢樹種,理想百分比為33.3%,以此類推來分別計算d值。認為當d取值最小時的上位種數為優勢樹種個數M1,最后將優勢度從大到小排序,選擇前M1個樹種作為該樣地的優勢樹種。
主要樹種確定標準是:12 塊蒙古櫟次生林樣地中,樹種均出現且斷面積占比達到1%。這樣能更充分反映林分內的競爭關系,而種內和種間競爭關系僅著重分析了優勢樹種。
2.2.2 競爭指數 選擇應用最廣泛的Hegyi競爭指數[15],計算方法如下:
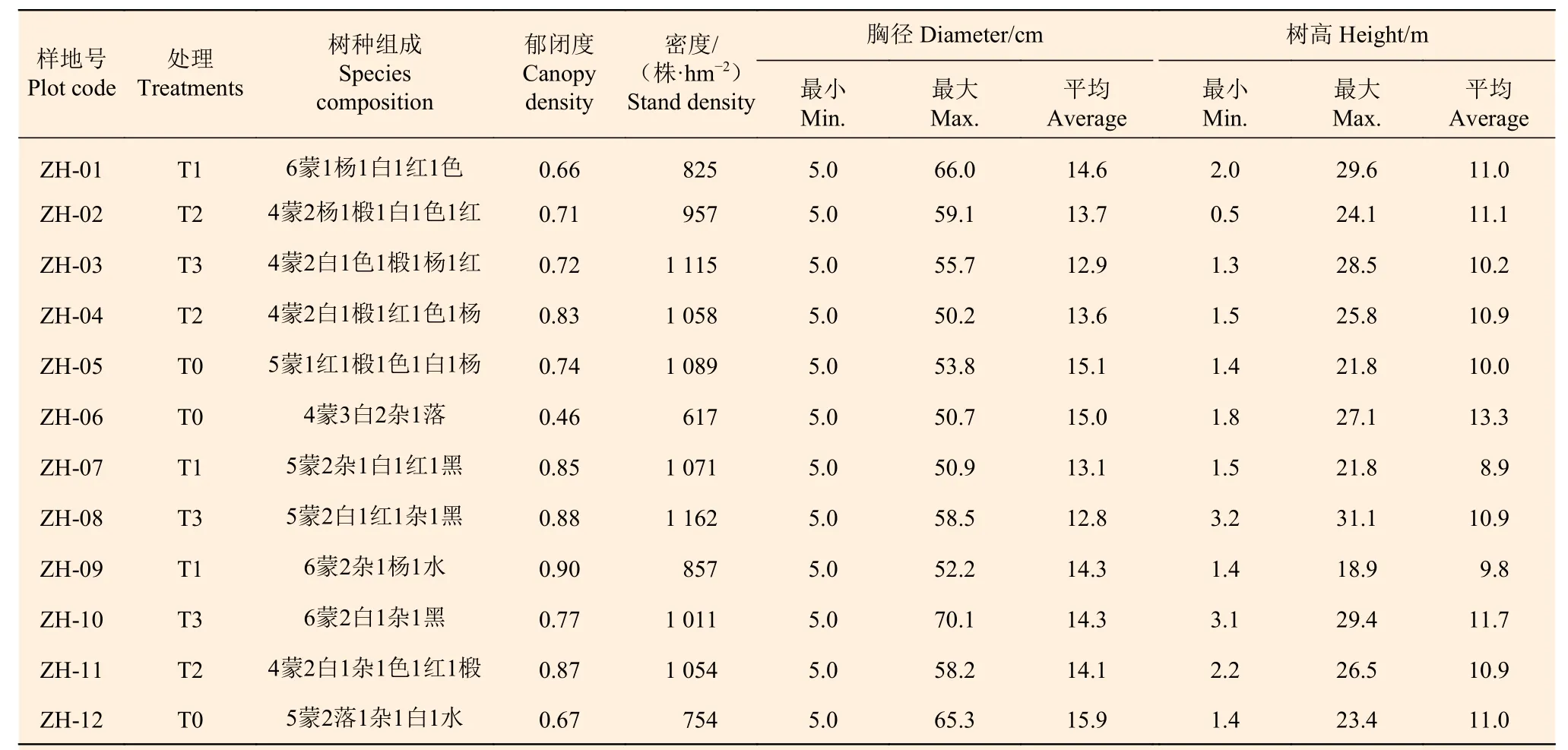
表1 樣地基本概況Table 1 General description of sample plots
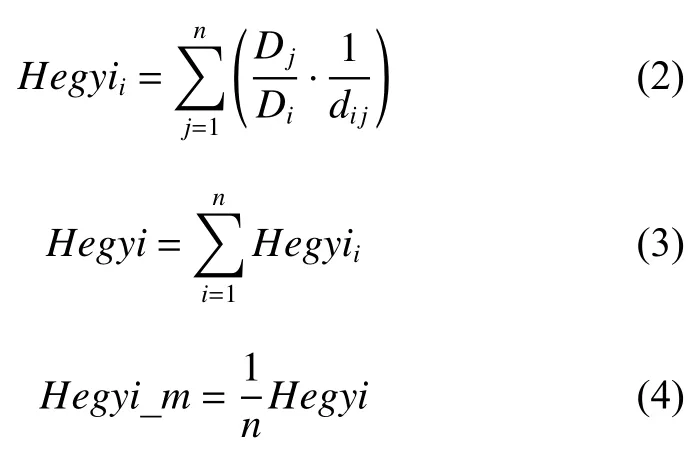
其中:Hegyii表示第i株對象木的競爭指數,Dj表示第j個競爭木的胸徑,Di表示第i個對象木的胸徑,dij表示第i個對象木與第j個競爭木之間的水平距離,n表示對象木的競爭木個數,Hegyi表示總競爭指數,Hegyi_m表示平均競爭指數。種內競爭指數(Hegyi_intra)和種間競爭指數(Hegyi_inter),定義為將競爭木中與對象木樹種相同的和不同的分別作為競爭木計算得出的競爭指數。平均種內競爭指數(Hegyi_intra_m)和平均種間競爭指數(Hegyi_inter_m)分別是Hegyi_intra和Hegyi_inter的平均值。這些競爭指數值越大代表競爭越激烈。
2.2.3 競爭木的選取 目前競爭木選擇的方法主要有胸高斷面積法[16]、固定半徑法[15]、影響域法等。一些學者[17]指出競爭木個數選擇應為4,而另一些研究有不同見解[18-21]。本研究采用有序樣本聚類分析的方法[22]來確定競爭木個數,即依次選取距離對象木最近的n株相鄰木作為競爭木,分別計算樣地的Hegyi、Hegyi_intra、Hegyi_inter、Hegyi_m、Hegyi_intra_m和Hegyi_inter_m。以這些競爭指數作為分類樣本,采用有序樣本聚類分析法分成2 類,根據分類結果的一致性來確定競爭木個數。有序樣本聚類算法采用最優分割法(或Fisher 法)進行分類,其遞推公式為:

式中φ表示分類的損失函數,p0表示某一種分類方案,n表示競爭木個數,這里n=3,4,5,···,14,D表示某類樣本離差平方和。
3 研究結果
3.1 優勢樹種和主要樹種
根據撫育經營前樣地調查的數據,利用公式(1)計算得出優勢樹種為蒙古櫟、紅松、白樺和雜木,這些樹種胸高斷面積和株數分別占樣地總量的75.41%和70.26%(表2)。雜木主要是青楷槭(Acer tegmentosumMaxim.)等小喬木樹種,多為叢狀分布,因此確定樣地的優勢樹種為蒙古櫟、紅松和白樺;主要樹種為蒙古櫟、白樺、紅松、色木、糠椴、大青楊、水曲柳、落葉松和黑樺(表2)。

表2 蒙古櫟樣地樹種株數及胸高斷面積Table 2 The number and basal area of tree species in Mongolian oak plots
3.2 競爭木確定
利用撫育經營前樣地調查數據,樣地內主要樹種的Hegyi和Hegyi_m隨競爭木株數n增加的計算結果如圖1 所示。
隨著競爭木增多,Hegyi和Hegyi_m呈上升趨勢,且上升趨勢逐步減緩。競爭強度較大樹種的競爭排位并不隨著選擇競爭木數量的增多而改變。而競爭強度較低的樹種的競爭排序因競爭木選擇的數量不同而發生變化。各競爭指數Hegyi、Hegyi_intra、Hegyi_inter、Hegyi_m、Hegyi_intra_m和Hegyi_inter_m的分類結果一致:[3,4,5,6,7]和[8,9,10,11,12,13,14],這種劃分下,組內殘差平方和最小,也即7 和8 為最優分割點。考慮到競爭木株數越多,計算值越穩定,本研究選取8 個競爭木來計算Hegyi,從而分析撫育經營前林分的競爭狀態及競爭對間伐的響應。

圖1 基于不同競爭木數量的主要樹種總競爭指數(a)和平均競爭指數(b)Fig.1 The total (a) and average (b) competition index of the main species based on the different number of competing neighbors
3.3 撫育經營前林木競爭狀態分析
單因子方差分析顯示Hegyi,Hegyi_intra和Hegyi_inter在T0,T1,T2 和T3 下無顯著差異。樣地內主要樹種的Hegyi排序為:蒙古櫟>紅松>色木槭>糠椴>白樺>大青楊>落葉松>水曲柳>黑樺,樹種的競爭排序和株數有明顯的正相關關系(圖2a)。樣地主要樹種的Hegyi_m排序為:水曲柳>色木槭>糠椴>紅松>白樺>蒙古櫟>落葉松>大青楊>黑樺(圖2b)。種內競爭大于種間競爭的樹種為:蒙古櫟、白樺、水曲柳,種間競爭大于種內競爭的為:紅松、色木槭、糠椴、大青楊、落葉松和黑樺,樣地內種內競爭(30132)略低于種間競爭(31937)(圖2a)。
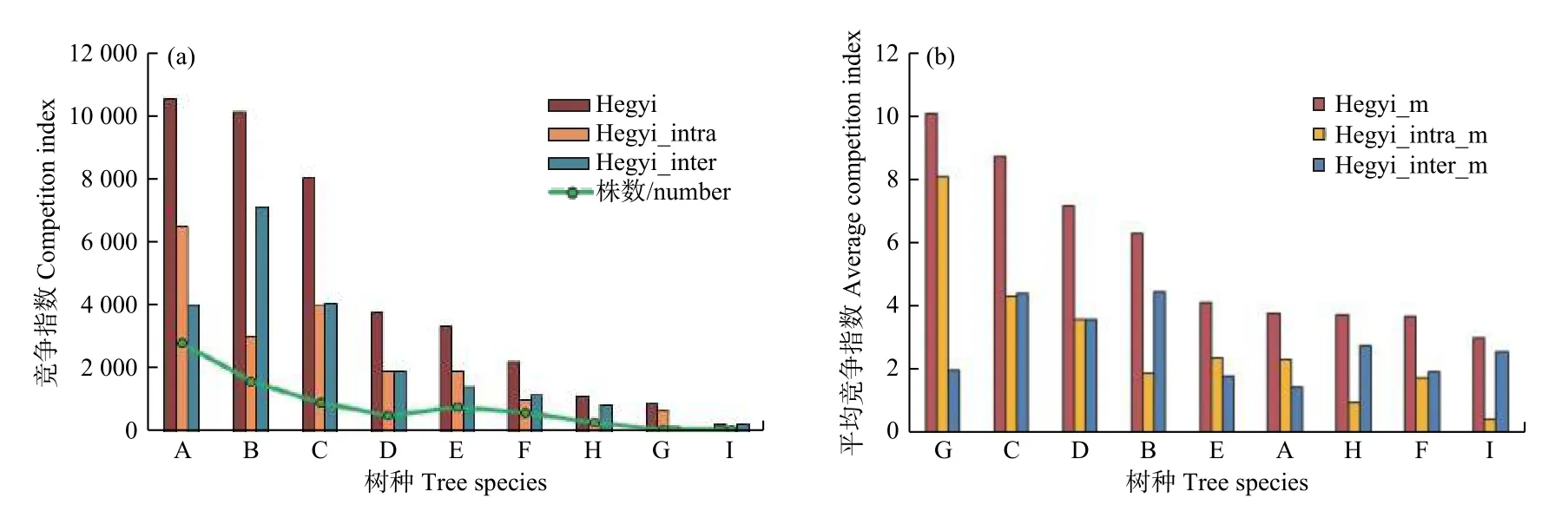
圖2 主要樹種的總競爭指數(a)和平均競爭指數(b)Fig.2 Total(a) and average(b) competition index of main tree species
3.4 林分內競爭對不同撫育間伐方式的響應
3.4.1 不同林層競爭強度對不同撫育間伐方式的響應 按照國際林聯(IUFRO)的標準劃分林層[23]。根據兩期調查數據(2013 年和2018 年)分別對各個林層的Hegyi和Hegyi_m進行統計(表3)。
2013 年各林層的Hegyi排序為中林層>下林層>上林層,2018 年T0,T2 和T3 各林層的競爭表現與2013 年類似,只有T1 的總競爭強度表現為中林層>上林層>下林層。2013 年和2018 年的Hegyi_m均呈現為下林層>中林層>上林層。Hegyi變化方面:T0 下林層降低53%,而中、上林層分別提高12%和14%,T1 下林層大幅降低(86%),但中上林層升高,T2,T3 上林層分別下降了19%和38%,下林層也呈降低趨勢,中林層均有所提高。就Hegyi_m的變化來說,T0 下林層和上林層分別下降了9%和8%,中林層提高14%,T1 下林層和中林層有所上升,上林層略有降低。T2 和T3 中林層和上林層下降,下林層上升。
3.4.2 優勢樹種競爭對不同撫育間伐方式的響應2018 年優勢樹種的競爭較2013 年發生了明顯變化(表4)。4 種方式下白樺和蒙古櫟的Hegyi和Hegyi_m均有呈下降趨勢,T2 紅松的Hegyi_m和T1,T3 紅松的Hegyi 和Hegyi_m 均呈下降趨勢。T2,T3 白樺的Hegyi和Hegyi_m下降幅度最大,T0 下降幅度最小,T3 紅松的Hegyi和Hegyi_m下降幅度較大,T1 對蒙古櫟的競爭影響最小。
3.4.3 種內種間競爭對不同撫育間伐方式的響應 4 種方式下Hegyi_intra_m均呈下降趨勢(圖3),分別下降18%,21%,13%和15%,Hegyi_inter_m基本呈上升趨勢(圖3),但上升幅度較小,其中T1 上升最明顯(8%)。

表3 不同撫育間伐方式下各林層的Hegyi 和Hegyi_mTable 3 Hegyiand Hegyi_m of different forest story in different treatment
由圖4 可知,蒙古櫟的種間競爭樹種主要是紅松和白樺。T0 方式下,兩期(2013,2018)蒙古櫟種間競爭樹種的競爭強度及排序變化不大。T1 方式下,蒙古櫟競爭樹種排序未發生變化,來自紅松的競爭有所提升,來自白樺和大青楊的競爭有所減少,其他樹種變化不大。T2 方式下,來自紅松和糠椴的競爭提高,來自白樺和大青楊的競爭降低。T3 方式下,來自白樺的競爭減少,而來自紅松的競爭提高。

表4 不同撫育間伐方式下優勢樹種Hegyi,Hegeyi_m 的變化Table 4 Changes of Hegyi and Hegeyi_m under different tending and thinning measures

圖3 Hegyi_intra_m 和Hegyi_inter_m 對不同撫育間伐方式的響應Fig.3 Response of Hegyi_intra_mand Hegyi_inter_mto different thinning treatment
紅松主要的競爭樹種是蒙古櫟,T0 紅松的競爭樹種強度排序變化不大,但來自落葉松和水曲柳的競爭強度增大,來自蒙古櫟、糠椴和大青楊的競爭強度有所下降。T1 主要競爭樹種的強度排序發生部分變化,但強度變化不大。T2 紅松的競爭樹種強度排序無變化,但來自白樺的競爭有所下降,來自色木槭和糠椴的競爭強度提高。T3 主要競爭樹種的排序變化不大,來自糠椴的競爭提高,排序靠前。對于白樺來說,主要的競爭樹種為蒙古櫟和紅松,其次為色木槭和大青楊。T0 白樺競爭樹種強度和競爭排序無明顯變化。T1 方式下,來自色木槭的競爭強度和排序提升。T2 方式下,來自大青楊的競爭強度下降,而來自色木槭的競爭強度升高。T3 競爭排序不變,但來自色木槭和糠椴的競爭強度提高,來自大青楊的競爭強度明顯下降。
綜上所述,T1,T0 方式下優勢樹種的競爭樹種組成和強度變化不大且無明顯趨勢,而T2 和T3 方式下,各優勢樹種來自糠椴、色木槭的競爭強度增大,來自大青楊的競爭強度減小。

圖4 優勢樹種的競爭樹種組成Fig.4 Species composition of competitive tree of dominant species
3.4.4 目標樹競爭對不同撫育間伐方式的響應 4 種方式下,目標樹的Hegyi_m均呈下降趨勢,T2 和T3 方式下目標樹的Hegyi_m和Hegyi_intra_m的下降幅度(14.0%,15.3%)明顯高于T0 和T1(4.4%,7.8%),4 種方式下目標樹的Hegyi_inter_m下降程度接近(表5)。目標樹的Hegyi變化趨勢與Hegyi_m變化趨勢和幅度一致。

表5 不同撫育間伐方式下目標樹和非目標樹的平均競爭指數Table 5 Average competition index of crop trees and other trees to different thinning treatment
目標樹的Hegyi_m均明顯小于非目標樹,T2和T3 方式下目標樹的Hegyi_m下降幅度分別為14%和15%,而非目標樹僅降低了3.8%和5.8%。T0 和T1 中非目標樹的種內競爭下降幅度要明顯大于目標樹的種內競爭下降幅度,而T2 和T3 呈現出相反的趨勢。目標樹和非目標樹的種間競爭變化不大(表5)。
4 討論
蒙古櫟次生林主要樹種的競爭強度排序隨競爭木數量增多的變化規律顯示:總競爭強度和平均競爭強度較大的樹種的競爭排序不隨競爭木數量的增多而改變,而競爭強度較小的樹種的排序變化較大,這與湯孟平等的研究結果類似[4]。蒙古櫟次生林主要樹種的種內競爭強度略小于種間競爭強度,這與多數研究[1,4,24-26]不一致。這可能與蒙古櫟次生林主要樹種的空間分布格局和混交程度有關系,林分的平均混交度[27]為0.602 3,表明對象木的競爭木為不同樹種的概率較高,種間競爭強度較大,導致種間競爭高于種內競爭。
植物物種之間的競爭能力取決于該物種的生態幅度和生態習性以及在生態系統的生態位[28]。不同種之間的生態位越接近,表明對自然資源的爭奪越激烈,競爭也就越激烈。可以看出平均競爭強度排序靠前的幾個樹種,水曲柳、色木槭、糠椴、紅松多是耐陰樹種,或說是在幼齡階段是耐陰樹種,因此在較強的競爭環境下也能生存,甚至可能自更新開始就處于較強的競爭環境下,而競爭較小的樹種多是喜光樹種,如楊樺、蒙古櫟和落葉松,這些樹種更新所需的光照強度較大,意味著生長空間就較大,因此受到的競爭較小。
對比蒙古櫟次生林不同林層的競爭得出與許多研究[29-30]一致的結果:高層林木平均競爭壓力要比中下層林木的小。比較不同方式的撫育間伐,發現傳統的撫育采伐有效降低了下林層的總競爭,而基于目標樹經營的撫育間伐有效緩解了上林層的總競爭和平均競爭。這與兩種采伐方式有關,傳統的撫育間伐是采弱留好,采密留稀,因此采伐對象多是處在中、下林層的林木,而基于目標樹經營的撫育間伐的目的是為目標樹釋放空間,目標樹多是林分的優勢木,處于上林層。
種內競爭對撫育間伐的響應要明顯高于種間競爭,針對目標樹和非目標樹的響應研究也呈現這種趨勢。這表明撫育間伐時采伐木與周圍林木是同一樹種的幾率較大,而在不采取任何措施的情況下,種內競爭對林木的生長影響更大。這是由于相同的樹種具有同樣的生態需求,因此種內個體競爭劇烈[1]。
目標樹的平均競爭明顯小于非目標樹,這是由于目標樹多是優勢木,所處的林層多于上林層,因此受到的競爭相對較小。T2 和T3 對目標樹競爭的減緩作用明顯優于T0 和T1,但T0 和T1 對非目標樹的競爭降低作用要強于T2 和T3。這也印證了目標樹經營理念,即只針對目標樹,對林分中其他不影響目標樹生長的林木不做考慮。隨著目標樹選擇株數的增多,采伐木數量提高,隨之應該帶來更大的競爭變化,但本研究中T2,T3 兩種目標樹選擇密度的林分內競爭關系沒有明顯差異,因此關于不同目標樹株數的選擇對林分內競爭關系的影響需要進一步研究。
5 結論
本研究利用12 塊1 hm2蒙古櫟次生林兩次調查數據開展了林木競爭關系對不同撫育間伐方式(傳統經營(T1)、目標樹經營密度1(T2)和目標樹經營密度2(T3)和對照試驗(T0))的響應關系分析。蒙古櫟次生林樣地的優勢樹種為:蒙古櫟、紅松和白樺,主要樹種的競爭排序為:蒙古櫟>紅松>色木槭>糠椴>白樺>大青楊>落葉松>水曲柳>黑樺,基于目標樹經營的撫育間伐對緩解上林層以及目標樹的競爭有明顯作用,可維持蒙古櫟和紅松在林分中的相對重要性;更注重培養有價值的樹種,有利于培育優勢樹種和頂級樹種,促進林分的正向演替。
