基于功能類群分析呼蘭河口濕地浮游植物群落結(jié)構(gòu)特征
2021-03-31 06:43:50范亞文陸欣鑫
生態(tài)學(xué)報 2021年3期
賈 鵬,范亞文,2,陸欣鑫,2,*
1 哈爾濱師范大學(xué)生命科學(xué)與技術(shù)學(xué)院, 哈爾濱 150025 2 黑龍江省水生生物多樣性研究重點實驗室, 哈爾濱 150025
浮游植物是濕地生態(tài)系統(tǒng)中的重要初級生產(chǎn)者,也是水生生物食物鏈中能量流動、物質(zhì)循環(huán)的基礎(chǔ)環(huán)節(jié)[1- 2],其群落結(jié)構(gòu)、物種組成的變化是反映水環(huán)境狀況的重要指標(biāo)[3- 4]。浮游植物的時空分布格局不僅可以影響濕地的生態(tài)功能和穩(wěn)定性,而且可以反映濕地生態(tài)環(huán)境的變化,也是探索濕地生態(tài)系統(tǒng)功能的重要手段之一[5- 6]。目前,在濕地生態(tài)系統(tǒng)中有關(guān)浮游植物的生態(tài)研究,主要集中在不同環(huán)境梯度和不同生活類型下浮游植物群落結(jié)構(gòu)的差異性分析[7- 9]。這類研究多使用傳統(tǒng)的浮游植物集群方法,不能有效地反映浮游植物在濕地生態(tài)系統(tǒng)中的功能。Reynolds[10]從個體環(huán)境適應(yīng)性、生態(tài)位競爭機(jī)制角度出發(fā),將具有相同環(huán)境適應(yīng)策略的浮游植物種類劃分為同一個類群,形成了具有相同生態(tài)適應(yīng)特征的分組,即“功能類群”。隨著功能類群研究的不斷深入,Salmaso[11]等通過植物功能型和FG功能類群的優(yōu)點提出了MFG劃分法;Kruk[12]等根據(jù)浮游植物的形態(tài)特征,提出了MBFG劃分法,這種劃分方法大大簡化了功能類群的應(yīng)用。目前,浮游植物功能類群方法已廣泛應(yīng)用在湖泊[13]、河流[14]、水庫[15]及濕地[16]等水域生態(tài)系統(tǒng)中。
濕地生態(tài)系統(tǒng)是水陸兩種生態(tài)系統(tǒng)的過渡區(qū)域,與湖泊、河流等水體相比,水體流動性和營養(yǎng)物質(zhì)更穩(wěn)定,具有防控洪水、調(diào)節(jié)徑流等功能[17]。近年來,由于人為活動干擾,導(dǎo)致濕地生態(tài)環(huán)境不斷惡化、生物多樣性降低和生態(tài)功能退化[18]。呼蘭河口濕地保護(hù)區(qū)位于哈爾濱市呼蘭區(qū)南部,松花江北岸,呼蘭河河口,東西長63.5km,南北寬21.3km,總面積為192.6 km2[19]。該濕地具有調(diào)節(jié)周邊小氣候、蓄洪調(diào)水和保護(hù)物種多樣性等生態(tài)功能[20]。隨著對濕地生態(tài)系統(tǒng)保護(hù)研究的不斷深入,目前已對呼蘭河口濕地高等動物[19]、高等植物[20]、水生生物[16]和浮游植物群落結(jié)構(gòu)[21- 22]開展了相關(guān)生態(tài)研究,以往的研究結(jié)果表明該濕地的浮游植物群落結(jié)構(gòu)表現(xiàn)出明顯的空間差異,FG功能類群分布特征對該濕地水環(huán)境空間異質(zhì)性響應(yīng)明顯[22]。
以呼蘭河口濕地保護(hù)區(qū)水域為主體,通過FG、MFG和MBFG三種功能類群對呼蘭河口濕地浮游植物群落進(jìn)行研究。定性、定量的評估浮游植物功能類群的演替模式;基于多元統(tǒng)計分析對驅(qū)動不同功能類群演替的環(huán)境因子進(jìn)行篩選與識別;初步分析不同功能類群對于呼蘭河口濕地水環(huán)境變化的響應(yīng)效果。為溫帶濕地生態(tài)特征研究提供新的分析手段,為我國溫帶河口濕地水生生物多樣性保護(hù)與受損濕地恢復(fù)提供理論依據(jù)。
1 材料與方法
1.1 采樣點設(shè)置
呼蘭河口濕地自然保護(hù)區(qū)位于黑龍江省哈爾濱市,北部與雙井鎮(zhèn)、方臺鎮(zhèn)相連,南與黃土山鄉(xiāng)、巨源鎮(zhèn)隔江相望。屬于北溫帶大陸性氣候,封凍期較長,氣溫較低,冬天漫長而寒冷;夏天暑期短,炎熱、多雨,年平均溫度3.6℃[21]。呼蘭河口濕地自然保護(hù)區(qū)內(nèi)水源地主要為兩處:一是呼蘭河,發(fā)源于小興安嶺西側(cè)、自東向西流動的中型河流;二是松花江,濕地核心區(qū)域則是由呼蘭河進(jìn)入松花江后形成[23]。
本研究于2018年春季(4、5月)、夏季(6、7、8月)和秋季(9、10月)三個季節(jié)對呼蘭河口濕地進(jìn)行樣品的采集,根據(jù)呼蘭河口濕地的生境特點共設(shè)置11個采樣點(圖1),各采樣點的海拔在110—130m之間。根據(jù)其生態(tài)特征,將11個采樣點劃分為四個生態(tài)功能區(qū)域,其中S1、S3、S11為入水口區(qū)域,受人為活動影響較大;S5、S7、S10為出水口區(qū)域,屬于松花江流域;S4、S6、S8、S9為濕地保護(hù)區(qū)域,保護(hù)區(qū)正在治理與恢復(fù);S2為農(nóng)田區(qū)域,受農(nóng)田耕種影響較大。利用GPS全球衛(wèi)星定位系統(tǒng)收集采樣點經(jīng)緯度信息,采樣點地圖的繪制通過ArcMap 10.2完成。

圖1 呼蘭河口濕地采樣點布設(shè)示意圖Fig.1 Map and sampling sites in Hulanhe Wetland樣點S1、S3、S11為入水口區(qū)域;樣點S2為農(nóng)田區(qū)域;樣點S5、S7、S10為出水口區(qū)域;S4、S6、S8、S9為濕地保護(hù)區(qū)域
1.2 樣品采集、處理及鑒定
分別于2018年4—10月末的上午對11個采樣點進(jìn)行定量樣品與定性樣品的采集。定性樣品采用25號浮游生物網(wǎng)在水面表層下0.5m處作“∞”字形緩慢撈取;定量樣品用1L的采水器在水表層以下0.5m處進(jìn)行水樣的采集,現(xiàn)場加入1.5%的魯格氏固定液15mL進(jìn)行固定,帶回實驗室沉淀24h后,濃縮至50mL,用光學(xué)顯微鏡進(jìn)行鏡檢計數(shù)[24]。浮游植物的鑒定、功能類群的劃分、生態(tài)分布信息主要參考胡鴻鈞、魏印心、胡韌等文獻(xiàn)[25- 28]。將至少在一個采樣點相對豐度超過15%的功能類群定義為代表性功能類群。
1.3 理化指標(biāo)的測定
采集水質(zhì)樣品與采集浮游植物樣品同步進(jìn)行,采用Hydrolab多參數(shù)水質(zhì)分析儀現(xiàn)場測定水溫(WT)、pH、電導(dǎo)率(SpCond.)、溶解氧(DO)、氧化還原電位(ORP)、濁度(Tur.)以及葉綠素a(Chl-a)。現(xiàn)場采集的水樣用棕色玻璃瓶避光保存,帶回實驗室24h內(nèi)依據(jù)《水和廢水監(jiān)測方法》(國家環(huán)境保護(hù)總局,2002)進(jìn)行總氮(TN)、總磷(TP)、高錳酸鹽指數(shù)(CODMn)、五日生化需氧量(BOD5)[29]的測量。
1.4 數(shù)據(jù)分析
使用PRIMER 5.0軟件進(jìn)行ANOSIM和SIMPER分析。獨立樣本T檢驗通過SPSS 20.0軟件完成。去趨勢分析(DCA)和冗余分析(RDA)采用軟件CANOCO 4.5進(jìn)行,在進(jìn)行排序分析前,對解釋變量和響應(yīng)變量數(shù)據(jù)進(jìn)行l(wèi)og(x+1)轉(zhuǎn)換,使樣本總體趨于正態(tài)分布。通過蒙特卡洛置換檢驗,判斷解釋變量對響應(yīng)變量生態(tài)分布影響的顯著性,建立二維排序圖。使用Origin 2018與Excel 2018軟件對圖表進(jìn)行繪制。
2 結(jié)果與分析
2.1 水體理化因子變化
獨立樣本t檢驗表明在不同季節(jié)之間水體理化因子變化明顯,除TP和SpCond.外其他理化指標(biāo)在季節(jié)之間均存在一定的變化規(guī)律(P<0.05)(表1)。DO、pH和Tur.都呈現(xiàn)隨春、夏、秋季節(jié)變化逐漸升高的規(guī)律(P<0.05),三季平均值分別為2.66、8.73、41.48mg/L,變化范圍分別在0.83—5.34mg/L、7.28—10.04和21.93—97.60mg/L之間;夏季的WT高于春、秋兩季(P<0.05),三季平均值為19.45℃,變化范圍在9.89—31.23℃之間;CODMn呈現(xiàn)出隨著春、夏、秋季節(jié)變化逐漸降低的規(guī)律(P<0.01),三季平均值為5.49mg/L,變化范圍在0.82—10.50mg/L之間;SpCond.和ORP呈現(xiàn)出春季最高,夏季最低的規(guī)律,三季平均值分別為402.57mS/cm和414.91mV,變化范圍在161.13—1180.00mS/cm和340.00—511.00mV之間。
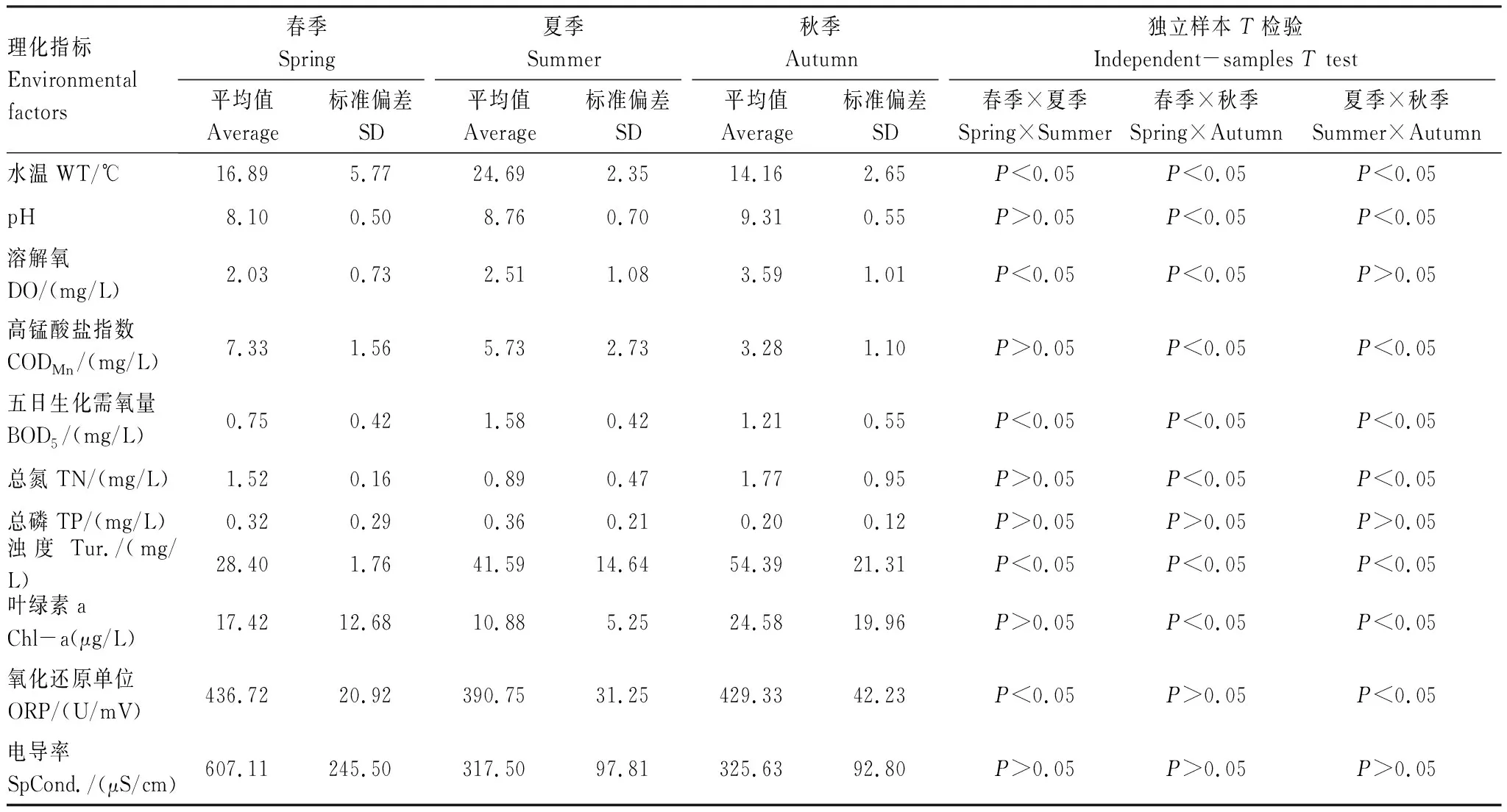
表1 呼蘭河口濕地水體理化因子季節(jié)變化
2.2 呼蘭河口濕地浮游植物種類組成
在研究期間共鑒定浮游植物243個分類單位,包括10變種,隸屬于7門9綱18目32科75屬。綠藻門(Chlorophyta)種類最多,為104種及8變種,占浮游植物種類總數(shù)的46.09%;裸藻門(Euglenophyta)種類46種及1變種,占19.34%;硅藻門(Bacillariophyta)種類44種及1變種,占18.52%;藍(lán)藻門(Cyanophyta)種類27種,占11.11%;甲藻門(Dinophyta)種類7種,占2.88%;隱藻門(Cryptophyta)種類3種,占1.23%;金藻門(Chrysophyta)種類僅2種,占0.82%。在空間尺度上,呈現(xiàn)濕地保護(hù)區(qū)域(188種)>入水口區(qū)域(183種)>出水口區(qū)域(172種)>農(nóng)田區(qū)域(135種)的規(guī)律;在時間尺度上,呈現(xiàn)夏季(198種)>秋季(152種)>春季(143種)的規(guī)律。
2.3 呼蘭河口濕地浮游植物豐度的時空變化
浮游植物豐度在時間尺度上表現(xiàn)為夏季平均值最高,變化范圍在(0.67—26.26)×106個/L之間;其次為秋季,變化范圍在(0.39—20.15)×106個/L之間;春季最低,變化范圍在(0.50—27.78)×106個/L之間。
浮游植物平均豐度呈現(xiàn)出濕地保護(hù)區(qū)域>入水口區(qū)域>農(nóng)田區(qū)域>出水口區(qū)域的規(guī)律。濕地保護(hù)區(qū)域內(nèi)的浮游植物平均豐度最高,變化范圍在(15.92—95.44)×106個/L之間;入水口區(qū)域浮游植物豐度次之,變化范圍在(4.47—25.40)×106個/L之間;農(nóng)田區(qū)域浮游植物豐度變化范圍在(1.28—27.78)×106個/L之間;出水口區(qū)域浮游植物豐度最低,變化范圍在(2.44—11.33)×106個/L之間(圖2)。

圖2 呼蘭河口濕地浮游植物豐度分布特征 Fig.2 Phytoplankton abundance spatial and temporal distribution of Hulanhe Wetland
2.3 FG、MFG和MBFG功能類群演替
根據(jù)Reynold和Padisák等提出的FG功能類群劃分方法,共劃分FG功能類群25個,分別為B/C/D/E/F/G/H1/J/LM/Lo/M/MP/N/P/S1/S2/SN/T/TB/W1/W2/X1/X2/X3/Y。其中代表性功能類群為B/D/F/J/Lo/MP/P/S1/W1/W2/X1/Y(表2,表3)。出現(xiàn)頻率超過50%的功能類群為B/D/F/H1/J/Lo/MP/P/S1/W1/W2/X1/X3/Y,出現(xiàn)頻率在30%—50%的功能類群為E/G/LM/X2,出現(xiàn)頻率小于30%的功能類群為C/M/N/T/TB/S2/SN(圖3)。FG功能類群季節(jié)演替特征為春季B(梅尼小環(huán)藻CyclotellameneghinianaKützing)/D(谷皮菱形藻Nitzschiapalea(Kützing) W.Smith)/Y(嚙蝕隱藻CryptomonaserosaHering)→夏季B/D/S1(湖泊假魚腥藻Pseudanabaenalimnetica(Lemmermann) Komárek)→秋季B/X1(狹形纖維藻AnkistrodesmusangustusBernard)(圖4)。
根據(jù)Salmaso等對功能類群的劃分[11],本次研究中呼蘭河口濕地共劃分MFG功能類群20個,分別為1b/1c/2c/2d/3a/3b/5a/5d/5e/6a/6b/6c/7a/8a/9a/9b/10a/11a/11b/11c。代表性功能類群為1c/2c/2d/5a/6a/6b/7a/9b/10a/11a/11b(表3)。出現(xiàn)頻率大于50%的功能類群為1b/1c/2c/2d/5a/5d/5e/6a/6b/7a/8a/9b/10a/11a/11b,出現(xiàn)頻率大于30%且小于50%的功能類群為11c/3b/6c/9a,功能類群3a出現(xiàn)頻率小于30%(圖3)。MFG功能類群季節(jié)演替特征為春季2d(隱藻門小個體)/7a(中心綱小個體)/6b(羽紋綱大個體)→夏季5a(顫藻目絲狀體)/6b/7a/→秋季7a/10a(綠藻綱絲狀體)(圖4)。
基于Kruk等研究結(jié)果[12],在呼蘭河口濕地共劃分MBFG功能類群6個,分別為Ⅰ/Ⅲ/Ⅳ/Ⅴ/Ⅵ/Ⅶ(表2,表3)。研究期間6種功能類群均出現(xiàn),其中Ⅴ類群在三個季節(jié)中都占據(jù)主要地位,出現(xiàn)頻率100%,Ⅳ和Ⅵ出現(xiàn)頻率超過98%,Ⅲ功能類群出現(xiàn)頻率最低但也超過了50%以上(圖3)。MBFG功能類群季節(jié)演替特征為春季Ⅵ(具有硅質(zhì)外壁且無鞭毛的種類)/Ⅴ(具鞭毛中到大型的單細(xì)胞種類)→Ⅵ/Ⅳ(不具有明顯特征的中等大小種類)→Ⅵ/Ⅳ(圖4)。

表2 呼蘭河濕地浮游植物功能群ANOSIM分析及SIMPER分析

表3 呼蘭河口濕地三種浮游植物功能類群的代表性屬種
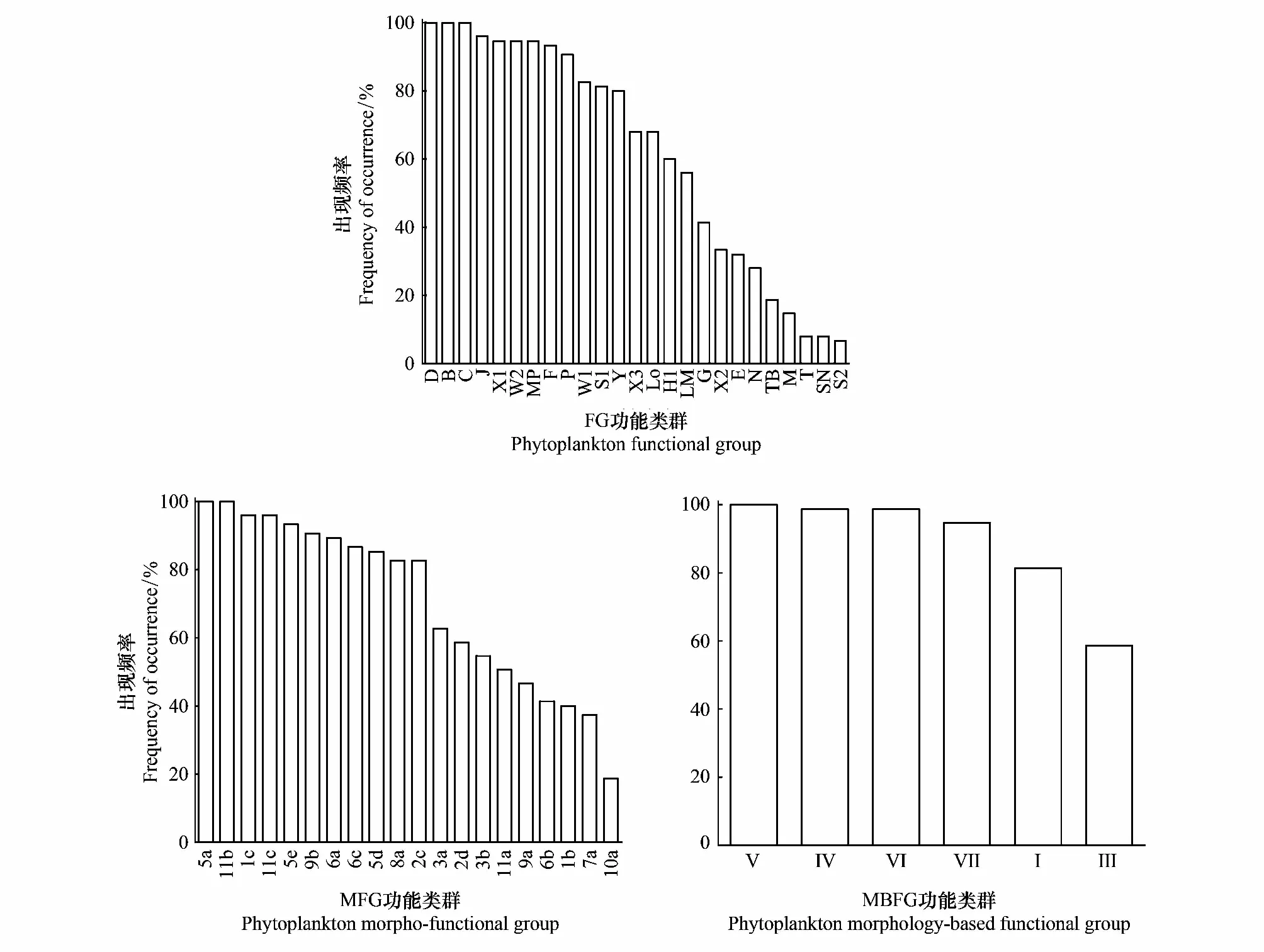
圖3 呼蘭河口濕地浮游植物功能類群出現(xiàn)頻率Fig.3 Frequency distribution of functional groups in Hulanhe Wetland

圖4 呼蘭河口濕地三種功能類群豐度相對百分比Fig.4 Relative percentage of abundance of three functional groups in Hulanhe Wetland
基于FG、MFG和MBFG功能類群群落相似性分析(ANOSIM),表明季節(jié)變化對浮游植物群落演替驅(qū)動作用明顯(P<0.05)。SIMPER分析表明,B/SI/Y/W2/X1/D/Lo、7a/5a/10a/2d/6b/2c/11a和Ⅵ/Ⅳ類群,分別為不同季節(jié)之間FG、MFG和MBFG類群演替的主要貢獻(xiàn)者。
2.4 多元統(tǒng)計分析
對呼蘭河口濕地的11種環(huán)境變量(WT、pH、SpCond.、DO、ORP、Tur.、Chl-a、TP、TN、CODMn與BOD5)與FG、MFG和MBFG功能類群進(jìn)行排序分析,不同功能類群對環(huán)境變化的響應(yīng)模式進(jìn)行識別。三種功能類群DCA分析顯示最長長度梯度分別為3.003和0.920和1.222,因此選擇線性模型RDA對優(yōu)勢功能類群與環(huán)境變量進(jìn)行排序分析。經(jīng)蒙特卡洛置換檢驗表明,呼蘭河口濕地三種功能類群的生態(tài)分布受多種環(huán)境因子共同作用影響,驅(qū)動功能類群生態(tài)分布的環(huán)境因子為:CODMn、WT、TP、SpCond.、pH、BOD5、Tur.、ORP和DO。

圖5 呼蘭河濕地植物功能類群與環(huán)境變量的RDA分析Fig.5 RDA of representative functional groups and environmental variables in Hulanhe wetland
3 討論
3.1 呼蘭河口濕地浮游植物功能類群演替特征
浮游植物功能類群方法從個體生態(tài)學(xué)角度出發(fā),基于物種生態(tài)適應(yīng)性角度分析種群的生態(tài)選擇機(jī)制[30]。不同功能類群內(nèi)的浮游植物種類具有其自身獨特的生態(tài)特征,其演替通常與水體營養(yǎng)狀態(tài)、光照及水溫密切相關(guān)[31],在“上行效應(yīng)”和“下行效應(yīng)”兩種機(jī)制調(diào)控下,當(dāng)棲息地水環(huán)境發(fā)生變化時,浮游植物自身的形態(tài)、生理適應(yīng)特征會主動或被動的適應(yīng)環(huán)境特征的改變,以演替的形式來維持濕地的生態(tài)平衡[32]。
呼蘭河口濕地受溫帶季風(fēng)氣候影響,季節(jié)之間水體理化環(huán)境特征變化明顯[14]。FG功能類群將物種的生態(tài)屬性劃分到功能類群當(dāng)中,其演替模式被認(rèn)為是響應(yīng)水環(huán)境變化的有效指標(biāo)[18]。在本研究中FG功能類群季節(jié)演替特征為,春季B/D/Y類群演替為夏季B/D/S1類群,最后被秋季的B/X1類群所取代。B類群是硅藻門中的小環(huán)藻屬,為α-ms(中營養(yǎng))指示種,其生活環(huán)境適宜在溫度20℃左右[33- 34]、高營養(yǎng)鹽的中小型較淺的水體中。水體中營養(yǎng)鹽含量的升高是導(dǎo)致富營養(yǎng)化的根本原因,其變化會直接影響浮游植物的豐度。在高營養(yǎng)鹽的水體中,浮游植物的豐度通常會受光照的限制,光的可利用性也會影響浮游植物的競爭和群落多樣性[35],因此耐受低光環(huán)境的類群通常會占據(jù)優(yōu)勢地位。呼蘭河口濕地在春季期間,水溫較低,日照時間相對較少,處于營養(yǎng)鹽積累階段,低水溫的條件會限制大部分浮游植物的生長,與此同時B類群具有高捕光效率和高相對表面積比的特點,因此在棲息地的適宜性及較低光照的基礎(chǔ)上,B類群在群落中占據(jù)優(yōu)勢地位[36]。Y類群的代表性物種為卵形隱藻和嚙蝕隱藻,適宜生活在磷濃度較高的水體中,對低光耐受,為廣適應(yīng)性種類[32]。由于春季濕地保護(hù)區(qū)域人為干擾較少,水體流動性差,磷濃度較高,同時Y類群物種具有鞭毛,有較強的運動性,使其能在水體中垂直運動,可以更好地躲避捕食者,進(jìn)而在資源競爭中占據(jù)有利地位[35]。因此,B和Y類群能在春季占據(jù)優(yōu)勢。D類群適應(yīng)于營養(yǎng)鹽含量較高的淺水水體、耐受于低光條件。呼蘭河口濕地夏季水體溫度升高,溶解氧含量和pH(平均值pH=8.76)增加,水體呈堿性。在一定范圍內(nèi),溫度和溶解氧的升高會促進(jìn)B/D類群的生長[37]。同時,水體的弱堿性為D類群提供了良好的生活條件,因此D類群能在夏季占據(jù)優(yōu)勢地位[38]。呼蘭河口濕地夏季降雨充沛,周邊土地中的營 養(yǎng)物質(zhì)通過淋溶作用進(jìn)入水體,入水口區(qū)域、農(nóng)田區(qū)域和濕地保護(hù)區(qū)域具有較多的人為活動干擾,因此導(dǎo)致水體變渾濁(濁度平均值為41.59 mg/L),限制光照攝入,進(jìn)而影響耐受低光環(huán)境的B/D類群生長。營養(yǎng)鹽是水體中重要的生源要素,由于呼蘭河口濕地水體氮磷比小于16∶1,水體受氮限制明顯。低氮的水體限制了大部分浮游植物的生長,而藍(lán)藻的生長與水體的氮磷比密切相關(guān)[39],S1類群中的絲狀藍(lán)藻具有固氮能力,因此營養(yǎng)鹽水平降低可能是S1類群中的絲狀藍(lán)藻(中富營養(yǎng))占據(jù)優(yōu)勢的重要原因。本文SIMPER分析也證明S1類群的變化是影響不同季節(jié)之間FG功能類群差異的主要功能類群之一。從生物因素的角度看,“上行效應(yīng)”和“下行效應(yīng)”對不同營養(yǎng)類型的水體作用是不一樣的,這是由浮游植物的粒徑大小導(dǎo)致的[40]。S1類群中的絲狀藍(lán)藻可抵御浮游動物的捕食,在低牧食壓力的情況下容易占據(jù)優(yōu)勢,因此S1類群代替了春季廣適應(yīng)性的Y類群,在夏季渾濁且溫暖的水體中與B/D類群共同占據(jù)主要優(yōu)勢地位。呼蘭河口濕地夏、秋兩季之間存在總氮含量明顯升高的過程(P<0.05),X1類群的代表性種為狹形纖維藻,為α-ms(富營養(yǎng))指示種,能夠耐受低溫環(huán)境,適宜在含氮量高的水體中生長[35]。因此替代了夏季的藍(lán)藻門S1類群。陸欣鑫等[22]對呼蘭河口濕地的研究也證明了X1類群在秋季占據(jù)優(yōu)勢地位。
MFG功能類群反映了浮游植物運動能力、大小、形狀、營養(yǎng)物質(zhì)喜好、營養(yǎng)鹽和碳的獲取能力等[40]。本次研究中,MFG功能類群季節(jié)性演替特征為由春季的2d/7a/6b類群演替為夏季的5a/7a/6b類群,最后由秋季的10a/7a類群所替代。MFG功能類群的季節(jié)演替特征所反映的水環(huán)境變化過程與FG功能類群差異不大。MBFG功能類群方法是利用浮游植物對水環(huán)境變化特征的響應(yīng)為標(biāo)準(zhǔn),反映某一特定生境模塊的狀況。ANOSIM分析表明,以MBFG功能類群為研究手段分析季節(jié)之間群落結(jié)構(gòu)差異效果不明顯。季節(jié)之間SIMPER分析也表明,在不同季節(jié)之間影響群落結(jié)構(gòu)差異的類群無變化。其原因可能是Ⅴ類群(具鞭毛中到大型的單細(xì)胞種類)和Ⅵ類群(具有硅質(zhì)外壁且無鞭毛的種類)為春季的主要優(yōu)勢功能類群,兩類群主要為隱藻和硅藻。春季低溫且低光照,具有鞭毛的隱藻根據(jù)自身的形態(tài)特點以及對低溫低光的耐受性,使其能在春季具有很強的競爭能力,與Ⅵ類群的硅藻在春季共同占據(jù)優(yōu)勢地位。夏季和秋季主要為Ⅵ類群的硅藻種類和Ⅳ類群中不具有明顯特征的中等大小種類,Ⅳ類群種類繁多,包含大部分除硅藻以外的其他浮游植物種類,此類群具有廣適應(yīng)性的特點,因此在夏、秋兩季占據(jù)優(yōu)勢地位。由于呼蘭河口濕地夏、秋兩季Ⅳ/Ⅵ類群對資源具有很強的競爭力,導(dǎo)致Ⅴ類群在水體中的生長被限制,這可能是Ⅴ類群在夏、秋兩季不占據(jù)優(yōu)勢的原因之一。
對比FG、MFG和MBFG三種功能類群的演替模式不難發(fā)現(xiàn),FG與MFG類群在描述呼蘭河口濕地浮游植物演替特征上有重疊部分,差異較小;MBFG與其他兩種功能類群相比,在表征季節(jié)間水環(huán)境的變化過程中效果并不理想。應(yīng)用功能類群方法對溫帶典型水庫中的研究曾指出,MBFG功能類群分類方法在水生態(tài)研究中雖然具有簡單、高效的優(yōu)勢,但對環(huán)境的解釋度不夠[41]。FG功能類群將浮游植物的定量和定性數(shù)據(jù)與多種水環(huán)境數(shù)據(jù)相結(jié)合,很大程度上提高了浮游植物對環(huán)境的響應(yīng)能力,將包含多種環(huán)境因子的復(fù)雜生態(tài)環(huán)境與浮游植物的耐受性和敏感性相關(guān)聯(lián),進(jìn)而說明浮游植物與環(huán)境因子之間的相關(guān)性,同時預(yù)測研究水體的營養(yǎng)狀態(tài)。雖然MFG弱化了對生態(tài)環(huán)境的考慮,導(dǎo)致對環(huán)境的解釋不如FG功能類群清晰,但仍然能夠揭示呼蘭河口濕地浮游植物群落季節(jié)變化特征。MBFG在響應(yīng)溫帶濕地季節(jié)間環(huán)境變化梯度的分析中,效果沒有FG和MFG功能類群理想。
3.2 影響呼蘭河口濕地浮游植物功能類群演替的環(huán)境因子
大量研究表明,溫帶湖泊、水庫及濕地水體中浮游植物群落的演替受浮游動物、濾食性魚類通過下行效應(yīng)影響,另一方面,水溫、光照、營養(yǎng)鹽及資源可利用率等上行效應(yīng)更是驅(qū)動群落演替的主要原因[39]。通過對中國3個典型湖泊浮游植物研究發(fā)現(xiàn),浮游植物功能類群對不同營養(yǎng)類型的湖泊響應(yīng)明顯[42]。基于MBFG功能類群對亞馬遜河流域浮游植物群落特征研究,得出MBFG類群能有效的指示水體電導(dǎo)率和pH分布特征[43]。呼蘭河口濕地是中型恢復(fù)性濕地,其入水口區(qū)域與農(nóng)田區(qū)域受季節(jié)性的人為活動干擾較大。浮游植物功能類群對濕地棲息環(huán)境的時空異質(zhì)性響應(yīng)存在獨特模式,優(yōu)勢類群的支配地位會隨著空間異質(zhì)性的改變而被其他類群所替代。營養(yǎng)鹽、pH、水溫、濁度和水動力特征等都是影響浮游植物出現(xiàn)或者消失的重要因素,同時影響各種功能類群的組成。RDA分析得出三種功能類群的演替是由多種環(huán)境因子共同作用的結(jié)果。經(jīng)過蒙特卡洛置換檢驗分析表明,pH、Tur.、BOD5和CODMn是影響呼蘭河口濕地浮游植物功能類群演替的最主要因子。
CODMn為指示浮游植物棲息環(huán)境有機(jī)污染的重要指標(biāo)之一,其數(shù)值越高說明水體富營養(yǎng)程度越高[44]。CODMn的變化往往會影響浮游植物的演替[45]。在研究期間,呼蘭河口濕地CODMn值在(0.82—10.50)mg/L之間變化,三種功能類群在三個季節(jié)間CODMn差異性都呈顯著水平(P<0.01)。RDA排序圖顯示,FG功能類群中的Y/D/W2類群、MFG功能類群中的2d/2c類群和MBFG功能類群中的Ⅴ類群都與CODMn值變化密切相關(guān),并都呈現(xiàn)出隨著CODMn值升高各類群豐度上升的趨勢,這些類群主要在春季和夏季中具有較高的權(quán)重。春季研究區(qū)域水體較淺且渾濁,營養(yǎng)物質(zhì)積累,由于春季農(nóng)田區(qū)域與入水口區(qū)域人為活動干擾,造成水體中有機(jī)污染物增多,因此春季的CODMn值最高,平均值為7.33mg/L。呼蘭河口濕地春季較高的CODMn值也伴隨著相對較高的磷含量,Y/D/2d/Ⅴ類群對水體中的磷濃度需求較大,因此在春季水體中高磷濃度的條件下,促進(jìn)了這些類群的生長。同時春季水溫和光照較低,耐受低光和低溫的B/D/7a/6b/Ⅵ類群在浮游植物群落中也占據(jù)優(yōu)勢地位。在一定程度上,CODMn值變化所反映出的水體有機(jī)物質(zhì)含量的改變也影響了FG功能類群的B/D/Y/S1/X1類群的生長與演替,此結(jié)果與同氣候帶的另一濕地(金河灣濕地)的研究結(jié)果近乎一致[16]。BOD5與CODMn共同反映水體中的有機(jī)物含量,RDA排序分析表明,S1/X1/5a/10a/Ⅶ類群的分布與BOD5含量密切相關(guān),隨著BOD5的升高,其種類豐度逐漸升高。BOD5值在夏、秋兩季較高,平均值分別為1.58mg/L和1.21mg/L,水體呈富營養(yǎng)狀態(tài),因此S1/X1/5a/10a類群較適應(yīng)夏、秋兩季的水體環(huán)境。Ⅶ類群體內(nèi)可貯存營養(yǎng)鹽,多為具有外膠被和較小表面積比的綠球藻目、顫藻目的群體,屬于冗余耐受生長策略[26],通過形成較大群體,以抵御外部的捕食壓力。同時BOD5與Chl-a也密切相關(guān),在適當(dāng)?shù)姆秶鷥?nèi),有機(jī)物質(zhì)會促進(jìn)浮游植物的代謝[39],浮游植物通過自身光合作用會產(chǎn)生大量的有機(jī)物釋放到水體中,使水體中的BOD5進(jìn)一步升高,這一結(jié)論在對巢湖相關(guān)的功能類群研究中也得到證明[46]。可見,BOD5所反映的水體中有機(jī)物含量的變化,對濕地中S1/X1/5a/10a/Ⅶ類群的生長、演替和分布都有著明顯的影響。
pH與浮游植物的分布關(guān)系密切,特別是與一些硅藻種類[47]。相關(guān)研究表明,pH變化可以通過影響胞內(nèi)部分酶活性或跨膜質(zhì)子梯度來影響浮游植物的光合作用過程,進(jìn)而影響浮游植物的生長[48]。RDA排序表明,FG功能類群的D/Y類群、MFG功能類群的6b類群和MBFG功能類群的Ⅴ類群均與pH呈負(fù)相關(guān)。浮游植物的光合及呼吸作用日變化研究表明,通常情況下浮游植物夜間進(jìn)行呼吸作用會釋放大量的CO2,造成水體pH降低,本研究樣品采集時間多為清晨至上午,這也是pH與浮游植物豐度呈負(fù)相關(guān)的原因之一[42];MBFG功能類群的Ⅳ/Ⅵ類群與pH變化密切相關(guān),浮游植物通過光合作用利用水中無機(jī)碳(H2CO3),產(chǎn)生氫氧根離子,導(dǎo)致水體pH升高,隨著其上升,導(dǎo)致水體pH與浮游植物豐度呈正相關(guān),因此水體中的pH適度的升高會影響Ⅳ/Ⅵ類群浮游植物的生長[49]。濁度是由有機(jī)與無機(jī)漂浮物、微生物和塵土等微小顆粒懸浮物質(zhì)所共同引起的[40],與光照強度、水體理化性質(zhì)以及氣候條件具有密切的關(guān)系,水體的濁度直接會影響浮游植物的豐度。RDA排序分析表明,Y/6b/Ⅴ類群隨著濁度的升高表現(xiàn)出豐度下降的趨勢。由于夏、秋兩季降雨充沛,營養(yǎng)鹽通過淋溶作用進(jìn)入水體,與此同時水體擾動較大,導(dǎo)致水體的濁度逐漸升高[39],較高的濁度會影響浮游植物的光合作用,使浮游植物產(chǎn)生的有機(jī)物質(zhì)減少,由于Y/6b/Ⅴ類群的個體相對較大,在相對面積的光照射下,這些類群的浮游植物光合作用更弱,從而導(dǎo)致Y/6b/Ⅴ類群在夏、秋兩季高濁度的水體中處于不利地位。B/10a/Ⅵ類群豐度的變化也與濁度密切相關(guān),由于濁度的增加會使水體的透明度下降,進(jìn)而限制了光照的射入,由于B類群的梅尼小環(huán)藻以及10a/Ⅵ類群的部分個體相對較小,在相同光照條件下,這些類群的光合作用更強,因此10a/Ⅵ類群更具有競爭優(yōu)勢。因此B/10a/Ⅵ類群在濁度升高的條件也會占據(jù)優(yōu)勢地位。呼蘭河口濕地是正在恢復(fù)中的濕地,本次研究與近8年前研究相比[21- 22],雖然優(yōu)勢種略有改變,但水生植物整體群落結(jié)構(gòu)較為穩(wěn)定。綜合來看,呼蘭河口濕地浮游植物群落演替受多種環(huán)境因子共同影響,其中pH、Tur.、BOD5和CODMn與三種浮游植物功能類群演替密切關(guān)系。本研究通過分析不同功能類群分布特征及環(huán)境相關(guān)性,以期進(jìn)一步完善濕地水生態(tài)系統(tǒng)的評價體系,這對呼蘭河口濕地水生生物多樣性保護(hù)與生態(tài)恢復(fù)具有一定的理論與指導(dǎo)意義。
4 結(jié)論
(1)呼蘭河口濕地在調(diào)查期間共鑒定浮游植物7門243個分類單位。共劃分FG功能類群25個,MFG功能類群20個,MBFG功能類群6個。
(2)相較于MBFG功能類群,FG和MFG功能類群能更好的響應(yīng)呼蘭河口濕地水環(huán)境的空間異質(zhì)性。
(3)CODMn和BOD5、pH、Tur.是驅(qū)動呼蘭河口濕地浮游植物群落結(jié)構(gòu)空間變化的主要環(huán)境因子。
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學(xué)學(xué)報(自然科學(xué)版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國科技論壇(2017年7期)2017-07-25 08:49:53
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學(xué)生導(dǎo)刊(2016年34期)2016-04-11 00:49:44
電測與儀表(2015年5期)2015-04-09 11:30:52
中國中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年22期)2014-03-01 04:32:55
