長鏈非編碼RNA在肝纖維化中的作用研究進(jìn)展
2021-03-31 03:53:22張春艷顏羽昕梁潔孟根斯立木馬月宏
解放軍醫(yī)學(xué)雜志 2021年2期
關(guān)鍵詞:小鼠
張春艷,顏羽昕,梁潔,孟根斯立木,馬月宏
內(nèi)蒙古醫(yī)科大學(xué)基礎(chǔ)醫(yī)學(xué)院,呼和浩特 010059
肝纖維化是肝臟結(jié)締組織異常增生的慢性病理過程,可導(dǎo)致細(xì)胞外基質(zhì)(extracellular matrix,ECM)大量累積,包括α-平滑肌肌動蛋白(α-smooth muscle actin,α-SMA)和Ⅰ型膠原α(type Ⅰ collagen α1 chain,Col1α1),并釋放促炎因子和促纖維化因子。多種慢性肝病如病毒性肝炎、酒精中毒、藥物濫用、代謝綜合征、遺傳代謝性疾病及自身免疫性肝炎等均會導(dǎo)致肝纖維化[1-2]。目前肝纖維化的調(diào)節(jié)機(jī)制尚不明確,激活肝星狀細(xì)胞(hepatic fibrosis,HSC)是肝纖維化發(fā)展的重要步驟。正常的HSC處于靜止?fàn)顟B(tài),由于某種病因損傷后HSC被激活,并被分化為成肌纖維母細(xì)胞樣的細(xì)胞,從而促進(jìn)纖維化的形成與發(fā)展。隨著對長鏈非編碼RNA(long non-coding RNA,lncRNA)生理作用及其在肝纖維化發(fā)病中作用機(jī)制的深入研究,通過調(diào)節(jié)肝纖維化患者體內(nèi)lncRNA活性治療肝纖維化有望成為現(xiàn)實(shí)。本文就lncRNA在肝纖維化中的作用研究進(jìn)展進(jìn)行綜述。
1 LncRNA概述
1.1定義 LncRNA是長度>200 nt的競爭性內(nèi)源RNA(competing endogenous RNA,ceRNA),也是RNA聚合酶Ⅱ轉(zhuǎn)錄的副產(chǎn)物,廣泛存在于動物、植物、酵母甚至病毒中,無編碼蛋白質(zhì)的功能。LncRNA與蛋白質(zhì)編碼基因相似,相比miRNA,其在細(xì)胞內(nèi)的轉(zhuǎn)錄比例更高,可在多個水平調(diào)控基因的表達(dá),在胚胎發(fā)育、細(xì)胞分化、疾病及腫瘤的發(fā)生過程中具有重要作用[3]。
1.2分類及作用 L n c R N A 可分為5類:同義lncRNA、反義lncRNA、雙向lncRNA、內(nèi)含子lncRNA及基因間lncRNA[4]。LncRNA具有4種作用:(1)支架作用。與兩種或兩種以上的蛋白質(zhì)相結(jié)合而發(fā)揮調(diào)控作用;(2)誘餌作用。誘導(dǎo)并結(jié)合一系列調(diào)控因子并阻礙其與相應(yīng)的功能位點(diǎn)結(jié)合而發(fā)揮調(diào)控作用;(3)引導(dǎo)作用。招募特定的蛋白質(zhì)并與之結(jié)合形成復(fù)合物而發(fā)揮調(diào)控作用;(4)信號作用。LncRNA可在空間和時間上反映轉(zhuǎn)錄因子或信號通路對基因的調(diào)節(jié)[4]。
2 促肝纖維化lncRNA
2.1肺腺癌相關(guān)轉(zhuǎn)錄本1(metastasis-associated lung adenocarcinoma transcript 1,MALAT1) MALAT1位于人類染色體11q13.1(小鼠染色體19qA),長約8 kb,在腫瘤細(xì)胞的增殖、遷移、侵襲中起到關(guān)鍵作用[5]。Yu等[6]發(fā)現(xiàn),在四氯化碳(CCl4)誘導(dǎo)的小鼠肝纖維化模型中,MALAT1的表達(dá)在活化的HSC中明顯上調(diào),且與miR-101b的表達(dá)呈負(fù)相關(guān)。MALAT1和RAS相關(guān)C3型肉毒桿菌底物1(ras-related C3 botulinum toxin substrate 1,Rac1)是miR101b的靶標(biāo),前者可作為ceRNA增強(qiáng)Rac1的表達(dá),促進(jìn)HSC的增殖和活化。Dai等[7]發(fā)現(xiàn),從亞砷酸鹽處理的人肝細(xì)胞中提取的MALAT1可通過與miRNA-26b結(jié)合促進(jìn)HSC細(xì)胞系LX-2的活化。
2.2漿細(xì)胞瘤變型異位子1(plasmacytoma variant translocation gene 1,PVT1) PVT1位于染色體8q24.21,在細(xì)胞核和細(xì)胞質(zhì)中均有分布。Yu等[8]的研究發(fā)現(xiàn),PV T1通過競爭性結(jié)合miR-152抑制PTCH1(patched 1),從而激活刺猬信號通路(Hedgehog signaling pathway)和上皮-間質(zhì)轉(zhuǎn)化(epithelial-mesenchymal transition,EMT)過程。自噬相關(guān)基因14(autophagy related gene 14,ATG14)是miR-152的直接靶標(biāo),通過PVT1-miR-152-ATG14信號通路誘導(dǎo)自噬有助于缺氧條件下HSC的激活,可促進(jìn)肝纖維化的發(fā)展。
2.3HOX轉(zhuǎn)錄反義基因間RNA(HOX transcript antisense intergenic RNA,Hotair) Hotair位于12號染色體,具體位于HOXC基因座內(nèi)HOXC11和HOXC12之間,包含6232個堿基對。Hotair在乳腺癌、胰腺癌、結(jié)腸癌及非小細(xì)胞肺癌等腫瘤中呈高表達(dá)[9]。Fu等[10]的研究發(fā)現(xiàn),Hotair可通過與miR-148b競爭性結(jié)合而上調(diào)DNA甲基轉(zhuǎn)移酶1(DNA methyltransferase 1,DNMT1),進(jìn)而誘導(dǎo)HSC的活化。此外,Hotair還可通過抑制母體表達(dá)基因3(maternally ex-pressed gene 3,MEG3)的表達(dá)而促進(jìn)HSC的活化。Yu等[11]的研究發(fā)現(xiàn),在CCl4誘導(dǎo)的肝纖維化小鼠中Hotair表達(dá)增加。Hotair通過下調(diào)miR-29b的表達(dá)減弱其介導(dǎo)的表觀遺傳機(jī)制,從而導(dǎo)致DNA甲基轉(zhuǎn)移酶3b(DNA methyltransferase 3b,DNMT3b)、第10號染色體缺失的磷酸酶及張力蛋白同源基因(phosphatase and tensin homolog deleted on chromosome ten,PTEN)的甲基化增強(qiáng),部分促進(jìn)HSC活化。
2.4肝纖維化相關(guān)lncRNA1(liver-enriched fibrosisassociated lncRNA1,LFAR1) LFAR1位于小鼠基因組的一個區(qū)域,該區(qū)域與人類染色體4q25同源,并與人CYP2U1和HADH基因相鄰。Zhang等[12]的研究發(fā)現(xiàn),LFAR1可促進(jìn)Smad2/3與轉(zhuǎn)化生長因子-β受體1(transforming growth factor beta receptor 1,TGFβR1)的結(jié)合及其在細(xì)胞質(zhì)中的磷酸化,還可直接與Smad2/3結(jié)合促進(jìn)Smad2、Smad3、Notch2、Notch3的轉(zhuǎn)錄。因此,LFAR1可通過激活轉(zhuǎn)化生長因子-β(transforming growth factor-β,TGF-β)和Notch通路,促進(jìn)肝纖維化。
2.5核富含轉(zhuǎn)錄本1(nuclear-enriched abundant transcript 1,NEAT1) NEAT1是長約3.7 kb的lncRNA,主要富集于細(xì)胞核中,是參與構(gòu)成細(xì)胞核亞結(jié)構(gòu)的重要RNA分子[13]。Yu等[14]發(fā)現(xiàn),NEAT1和Kruppel樣因子6(Kruppel-like factor 6,KLF6)是miR-122的靶標(biāo)。NEAT1可通過競爭性結(jié)合miR-122來調(diào)節(jié)肝纖維化中KLF6的表達(dá)。同時,NEAT1/miR-122/KLF6在HSC激活過程中起關(guān)鍵作用。Kong等[15]發(fā)現(xiàn),胰島素樣生長結(jié)合蛋白相關(guān)蛋白1(insulin-like growth factor binding proteinrelated protein 1,IGFBPrP1)可增高小鼠肝臟組織中NEAT1、自噬相關(guān)蛋白9a(autophagy associated proteins 9a,Atg9a)的表達(dá)及自噬水平,降低miR-29b的表達(dá)水平。其中Atg9a參與了IGFBPrP1誘導(dǎo)的HSC自噬和激活。因此,NEAT1/miR-29b/Atg9a調(diào)控軸參與了IGFBPrP1誘導(dǎo)的小鼠HSC的自噬和 激活。
2.6被TGF-β活化的長鏈非編碼RNA(long noncoding RNA activated by TGF-β,lncRNA ATB) LncRNA ATB位于14號染色體上,是長度超過8 kb的lncRNA。在丙型肝炎病毒(hepatitis C virus,HCV)相關(guān)肝纖維化患者的肝臟組織、血清及HSC中l(wèi)ncRNA ATB表達(dá)均上調(diào)[16-17]。LncRNA ATB通過競爭性結(jié)合miRNA-425-5p而促進(jìn)Smad2的表達(dá)以及HSC的增殖和活化[16]。Fu等[17]發(fā)現(xiàn),在HCV相關(guān)肝纖維化患者的肝臟組織、血清及HSC中miR-200a表達(dá)下降,β-連環(huán)蛋白(β-catenin)表達(dá)上調(diào)。β-catenin是miR-200a的靶標(biāo),lncRNA ATB作為ceRNA可與miR-200a結(jié)合,促進(jìn)β-catenin的表 達(dá)[17]。因此,lncRNA ATB/miR-200a/β-catenin調(diào)節(jié)軸可能促進(jìn)了肝纖維化的發(fā)展。
2.7長鏈非編碼RNA H19(long non-coding RNA H19,lncRNA H19) LncRNA H19位于人類同源區(qū)域染色體11p15.5,為長約2.3 kb的lncRNA。Song等[18]發(fā)現(xiàn),上皮細(xì)胞黏附分子(epithelial cells adhere to molecules,EpCAM)是電子盒同源盒1(electronic box homologous box 1,ZEB1)的下游靶基因,lncRNA H19和EpCAM的向上調(diào)節(jié)與ZEB1的向下調(diào)節(jié)呈正相關(guān)。LncRNA H19通過激活ZEB1/EpCAM信號通路而促進(jìn)小鼠的肝纖維化。Zhu等[19]的研究發(fā)現(xiàn),lncRNA H19作為ceRNA可與miR-148a相結(jié)合,并隨后維持泛素特異性蛋白酶4(ubiquitin-specific protease 4,USP4)的水平,USP4是miR-148a的一個已確定的靶標(biāo)。該研究結(jié)果揭示了一個新的lncRNA H19/miR-148a/USP4軸,該軸通過HSC的激活和肝細(xì)胞中TGF-β途徑來促進(jìn)肝纖維化。Wang等[20]發(fā)現(xiàn),lncRNA H19通過上調(diào)酒精脫氫酶Ⅲ(alcohol dehydrogenase Ⅲ,ADH3)介導(dǎo)的維甲酸信號從而誘導(dǎo)HSC的激活;Huang等[21]發(fā)現(xiàn),在轉(zhuǎn)染IGFBPrP1的JS-1細(xì)胞中,細(xì)胞自噬活躍程度與lncRNA H19表達(dá)水平呈正相關(guān)。因此,lncRNA H19可通過磷脂酰肌醇-3激酶(phosphatidylinosital 3-kinase,PI3K)/蛋白激酶B(protein kinase B,Akt)/哺乳動物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)信號通路來促進(jìn)IGFBPrP1誘發(fā)HSC自噬。
2.8小核RNA宿主基因7(small nucleic RNA host gene 7,SNHG7) SNHG7位于9號染色體,長度為2157 nt,在人類肝纖維化組織和活化的小鼠原代HSC中表達(dá)上調(diào)。Yu等[22]發(fā)現(xiàn),lncRNA SNHG7可競爭性結(jié)合miR-378-3p,致使細(xì)胞內(nèi)游離的紊亂片段極性蛋白2(disorder fragment polarity protein 2,DVL2)含量增加,β-catenin在細(xì)胞內(nèi)大量積累,促進(jìn)了HSC的激活,誘導(dǎo)了肝纖維化的發(fā)展。
2.9INK4位點(diǎn)的反義非編碼RNA(antisense noncoding RNA in the INK4 locus,ANRIL) ANRIL位于7號染色體,首先發(fā)現(xiàn)于遺傳性黑色素瘤和神經(jīng)系統(tǒng)腫瘤患者。Yang等[23]發(fā)現(xiàn),在活化的HSC和肝纖維化組織中ANRIL的表達(dá)顯著降低,DNA甲基轉(zhuǎn)移酶3A(DNA methyltransferase 3A,DNMT3A)的表達(dá)顯著增加。下調(diào)DNMT3A可增加活化的HSC中ANRIL的表達(dá),而過表達(dá)ANRIL可抑制HSC的激活和腺苷酸活化蛋白激酶(AMP-activated protein kinase,AMPK)信號通路。因此,lncRNA ANRIL缺失可以激活A(yù)MPK途徑,從而促進(jìn)肝纖維化和HSC的活化。
2.10軟骨發(fā)生刺激因子1(stimulator of chondrogenesis 1,SCRG1) SCRG1是一種轉(zhuǎn)錄長度為3118 bp的lncRNA,在人肝硬化組織中其表達(dá)上調(diào)13.62倍。Wu等[24]發(fā)現(xiàn),SCRG1在人體組織和轉(zhuǎn)化生長因子-β1(transforming growth factor-1,TGF-β1)誘導(dǎo)的活化LX-2細(xì)胞中的表達(dá),可隨著肝纖維化的發(fā)展而增加。SCRG1與鋅指蛋白36(tristetraprolin,TTP)有結(jié)合位點(diǎn),并可特異性結(jié)合TTP蛋白。過表達(dá)SCRG1會導(dǎo)致TTP mRNA不穩(wěn)定,蛋白質(zhì)表達(dá)減少。SCRG1靶向下調(diào)TTP可導(dǎo)致腫瘤壞死因子α (tumor necrosis factor alpha,TNF-α)和基質(zhì)金屬蛋白酶2(matrix metalloproteinase,MMP2)的激活,促進(jìn)肝纖維化的發(fā)展。
2.11X染色體失活特異轉(zhuǎn)錄物(X-inactive specific transcript,XIST) XIST位于X染色體無活性中心區(qū)域,長約16 500 nt,作為染色體Xq13.2的轉(zhuǎn)錄產(chǎn)物,能夠影響X染色體相關(guān)基因的激活。XIST與miR-29b結(jié)合可促進(jìn)高遷移率族蛋白盒-1(highmobility group box-1,HMGB1)的表達(dá),從而增強(qiáng)乙醇誘導(dǎo)的LX-2細(xì)胞自噬和激活[25-26]。因此,lncRNA XIST通過miR-29b/HMGB1途徑增強(qiáng)乙醇誘導(dǎo)的LX-2細(xì)胞的自噬和激活,從而促進(jìn)肝纖維化的 發(fā)展。
2.12小Cajal體特異性RNA 10(small Cajal bodyspecific RNA 10,SCARNA10) SCARNA10位于12號染色體,是一種核內(nèi)滯留的lncRNA,也是一種潛在的生物標(biāo)志物。SCARNA10過表達(dá)可加重CCl4誘導(dǎo)的小鼠肝纖維化,且SCARNA10在小鼠肝纖維化組織中表達(dá)顯著上調(diào)[27]。SCARNA10在體內(nèi)、體外均可通過誘導(dǎo)肝細(xì)胞凋亡和HSC活化而促進(jìn)肝纖維化的發(fā)展。Zhang等[28]發(fā)現(xiàn),SCARNA10可與多梳抑制復(fù)合物2(polycomb repressive complex 2,PRC2)結(jié)合而增加Smad2、Smad3、TGF-β1和KLF6的表達(dá),從而促進(jìn)肝纖維化的發(fā)展。
2.13長鏈基因間非編碼RNA01093(long intergenic noncoding RNA 01093,linc01093) Linc01093位于4號染色體。目前主要通過芯片、高通量測序技術(shù)篩選腫瘤組織中差異表達(dá)的linc01093。Tang等[29]發(fā)現(xiàn),在CCl4誘導(dǎo)的肝組織和TGF-β1刺激的肝細(xì)胞中l(wèi)inc01093表達(dá)下調(diào)。Linc01093的表達(dá)下調(diào)促進(jìn)了肝細(xì)胞的凋亡,抑制了肝細(xì)胞的存活。在TGF-β1的刺激下,下調(diào)的linc01093通過促進(jìn)沉默信息調(diào)節(jié)因子1 (silent information regulator 1,SIRT1)的降解和泛素化來促進(jìn)肝細(xì)胞凋亡。
2.14Alu介導(dǎo)的p21轉(zhuǎn)錄調(diào)節(jié)因子(alu-mediated p21 transcriptional regulator,APTR) APTR位于染色體7q21,長度為2303 bp,對細(xì)胞周期和細(xì)胞增殖具有抑制作用。Yu等[30]發(fā)現(xiàn),APTR在肝纖維化組織中表達(dá)上調(diào),其沉默可誘導(dǎo)p21的轉(zhuǎn)錄,從而抑制U87細(xì)胞的生長。APTR與p21在小鼠肝纖維化組織中的表達(dá)呈負(fù)相關(guān),可通過負(fù)調(diào)控p21加速細(xì)胞周期,對HSC具有促增殖的作用。
2.15核糖核酸HOXA轉(zhuǎn)錄本(ribonucleic acid HOXA transcript at the distal tip,HOTTIP) HOTTIP位于染色體7p152,在多種人類腫瘤(包括胃癌、肝細(xì)胞癌和結(jié)直腸癌)中起關(guān)鍵作用[31-32]。Li等[33]發(fā)現(xiàn),HOTTIP對miR-148a具有負(fù)調(diào)控作用,作為miR-148a、TGF-βR1和TGF-βR2的新靶標(biāo),可增強(qiáng)TGF-β/Smad信號傳導(dǎo)。HOTTIP可通過下調(diào)miR-148a,增強(qiáng)TGF-βR1和TGF-βR2的表達(dá),從而促進(jìn)肝纖維化的發(fā)生和發(fā)展。Zheng等[34]發(fā)現(xiàn),HOTTIP作為miR-150的ceRNA,可增加血清反應(yīng)因子(serum response factor,SRF)的表達(dá),誘導(dǎo)小鼠HSC的活化。
2.16牛磺酸調(diào)節(jié)基因1(taurine up-regulated gene 1, TUg1) TUg1是大小為7.1 kb的lncRNA,首次發(fā)現(xiàn)于牛磺酸處理的小鼠視網(wǎng)膜細(xì)胞上調(diào)表達(dá)基因中。Han等[35]發(fā)現(xiàn),TUg1在CCl4及膽管結(jié)扎(BDL)誘導(dǎo)的肝纖維化小鼠模型的肝臟組織以及肝硬化患者中表達(dá)上調(diào)。TUg1可促進(jìn)α-SMA、Col1α1、MMP2/MMP9/MMP10和基質(zhì)金屬肽酶組織抑制因子1(tissue inhibitor of metalloproteinases 1,TIMP1)的表達(dá)。miR-29b是TUg1的靶標(biāo),TUg1可負(fù)向調(diào)節(jié)miR-29b的表達(dá)。因此,lncRNA TUg1作為ceRNA可下調(diào)miR-29b的表達(dá)而促進(jìn)HSC的活化,從而促進(jìn)肝纖維化的發(fā)展。
LncRNA促進(jìn)肝纖維化的作用匯總見表1。
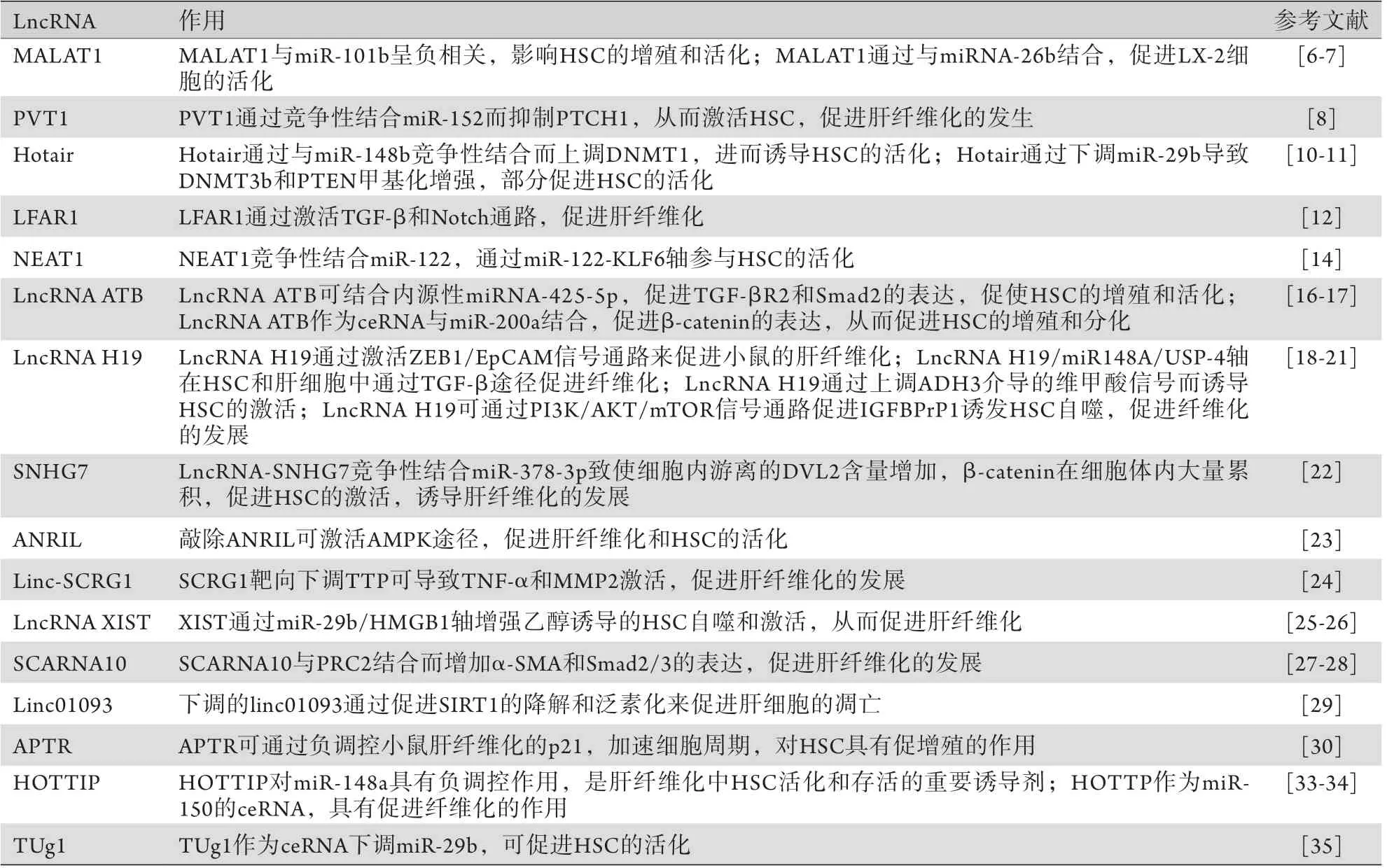
表1 LncRNA促進(jìn)肝纖維化的作用Tab.1 The promotive effects of lncRNA on liver fibrosis
3 抑制肝纖維化lncRNA
3.1母系表達(dá)基因8(maternally expressed gene 8,MEG8) MEG8位于染色體14q32.3上的一簇印跡基因中,主要分布在LX-2細(xì)胞和AML12細(xì)胞的細(xì)胞核內(nèi)。MGE8可促進(jìn)肺癌和胰腺癌細(xì)胞EMT的表觀遺傳學(xué)進(jìn)程,抑制血管平滑肌細(xì)胞(vascular smooth muscle cell,VSMC)和滋養(yǎng)細(xì)胞的增殖和遷移[36-37]。研究發(fā)現(xiàn),MEG8在活化的HSC、損傷的肝細(xì)胞和肝纖維化組織中過表達(dá)[38]。MEG8可通過抑制Notch通路而抑制肝細(xì)胞的HSC活化和EMT過程[38]。MEG8作為Notch信號的新型調(diào)節(jié)劑,為肝纖維化的分子機(jī)制研究提供了新的理論依據(jù)。
3.2預(yù)測基因5091(predicted gene 5091,Gm5091) Gm5091是一種長度為1179 bp的基因間lncRNA,位于Chr17。乙醇處理可使Gm5091在HSC中的表達(dá)下調(diào),并對乙醇誘導(dǎo)的HSC激活和炎癥產(chǎn)生負(fù)調(diào)控作用。Zhou等[39]發(fā)現(xiàn),Gm5091序列包含miR-27b、miR-23b、miR-24的結(jié)合位點(diǎn),且通過RNA下拉測定法證實(shí)miR-27b/23b/24均可與Gm5091結(jié)合。全長Gm5091可降低miR-27b/23b/24的水平,但截短的Gm5091不能刪除結(jié)合位點(diǎn)。Gm5091/miR-27b/23b/24可以減輕酒精性肝纖維化(alcoholic hepatic fibrosis,AHF)。
3.3長鏈基因間非編碼RNA-p21(long intergenic noncoding RNA-p21,lincRNA-p21) LincRNA-p21位于編碼關(guān)鍵細(xì)胞周期調(diào)節(jié)因子Cdkn1a(也稱為p21)的基因上游約15 kb,包含兩個外顯子,長約3.1 kb。LincRNA-p21可作為TGF-β的效應(yīng)器與miR-30相互作用,從而促進(jìn)小鼠的肝纖維化[40];還可作為ceRNA與miR-181b競爭性結(jié)合,減弱miR-181b對PTEN表達(dá)的抑制作用,最終抑制HSC的活化[41]。Yu等[42]發(fā)現(xiàn),lincRNA-p21作為ceRNA可與miR-17-5p相結(jié)合,上調(diào)糖原合成激酶3β(glycogen synthesis kinase 3β,GSK-3β)的表達(dá),抑制Wnt/β-catenin信號通路。因此,通過lincRNA-p21/miR-17-5p/β-catenin信號通路軸,lincRNA-p21能夠抑制HSC的活化。
3.4長鏈非編碼RNA-Hser(long non-coding RNAHser,lnc-Hser) Lnc-Hser主要位于初級肝細(xì)胞和AML12細(xì)胞的細(xì)胞核中。Zhang等[43]發(fā)現(xiàn),敲除lnc-Hser可通過誘導(dǎo)EMT和肝細(xì)胞凋亡在體內(nèi)和體外加重肝纖維化。同時,lnc-Hser可通過補(bǔ)體C5a受體1(complement C5a receptor 1,C5AR1)-河馬(Hippo)-YES相關(guān)蛋白(YES-associated protein,YAP)途徑抑制HSC的凋亡,并通過Notch途徑抑制肝細(xì)胞內(nèi)EMT的累積。總之,lnc-Hser是受損肝細(xì)胞的新型生物標(biāo)志物,也是抗纖維化治療的潛在靶標(biāo)。
3.5生長停滯特異性轉(zhuǎn)錄本5(growth arrest-specific transcript 5,GAS5) GAS5最初是因在生長抑制的鼠纖維原細(xì)胞中呈高表達(dá)而被發(fā)現(xiàn)的,位于人染色體1q251的小開放閱讀框,全長630個核苷酸。 Yu等[44]和Dong等[45]發(fā)現(xiàn),CCl4誘導(dǎo)的大鼠肝纖維化模型中,miR-23a可通過PTEN/PI3K/Akt/mTOR/Snail信號通路促進(jìn)纖維化的發(fā)展。GAS5可與miR-23a競爭性結(jié)合,降低miR-23a的表達(dá),從而抑制肝纖維化。GAS5/miR-23a/PTEN/PI3K/Akt/mTOR/Snail通路在肝纖維化中的作用為肝纖維化的治療提供了潛在的分子靶點(diǎn)。
3.6母系表達(dá)基因3(maternal expressed gene 3,MEG3) MEG3位于人類染色體14q32.3的DLK1-MEG3位點(diǎn),長度為35 kb,具有抑癌作用。Yu等[46]的研究發(fā)現(xiàn),MEG3是一種腫瘤抑制基因,在EMT過程中發(fā)揮重要作用。MEG3在體內(nèi)和體外肝纖維化過程中表達(dá)均降低,其表達(dá)恢復(fù)后,肝纖維化得到抑制,α-SMA和Col1α1表達(dá)降低。值得注意的是,MEG3過表達(dá)抑制了通過EMT激活的HSC,這與上皮標(biāo)志物的增加和間質(zhì)標(biāo)志物的減少有關(guān)。進(jìn)一步研究發(fā)現(xiàn),刺猬信號通路介導(dǎo)的EMT過程在肝纖維化的發(fā)生發(fā)展中具有重要作用,平滑蛋白(smoothened,SMO)是Hh途徑的一員[46]。此外,MEG3被確認(rèn)為是miR-212的靶標(biāo)。miR-212部分參與了MEG3對EMT過程的影響。總之,MEG3通過SMO蛋白和miR-212抑制了刺猬信號通路介導(dǎo)的肝纖維化EMT過程。
LncRNA抑制肝纖維化的作用匯總見表2。
4 總結(jié)與展望
LncRNA是近年肝病領(lǐng)域研究的熱點(diǎn),在肝癌(liver cancer,HCC)、肝纖維化、非酒精性脂肪肝(non-alcoholic fatty liver disease,NAFLD)中的研究取得了突破性進(jìn)展。但多數(shù)lncRNA在肝纖維化中的作用機(jī)制尚未完全闡明。缺失上調(diào)的lncRNA或過度表達(dá)下調(diào)的lncRNA是進(jìn)行l(wèi)ncRNA基礎(chǔ)治療的主要機(jī)制。未來研究應(yīng)致力于尋找準(zhǔn)確地將lncRNA導(dǎo)入體內(nèi)的方法并深入研究其機(jī)制;準(zhǔn)確鑒定lncRNA的靶標(biāo),使導(dǎo)入體內(nèi)的lncRNA作用于特定的與肝纖維化發(fā)展相關(guān)的靶標(biāo)。同時,希望臨床研究人員進(jìn)一步提高對lncRNA在肝纖維化調(diào)控中作用的認(rèn)識,從而有效地篩選lncRNA用于肝纖維化的診斷和治療,為臨床試驗(yàn)提供理論依據(jù)。

表2 LncRNA抑制肝纖維化的作用Tab.2 The inhibitory effects of lncRNA on liver fibrosis
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學(xué)大眾(2021年6期)2021-07-20 07:42:44
科學(xué)(2020年3期)2020-11-26 08:18:30
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學(xué)生學(xué)習(xí)指導(dǎo)(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學(xué)學(xué)報(自然科學(xué)版)(2015年2期)2016-01-10 08:41:55
云南中醫(yī)學(xué)院學(xué)報(2014年3期)2014-07-31 18:57:34
