未產(chǎn)卵雌性黃鱔的性轉(zhuǎn)變
2021-04-01 07:56:08張利紅張為民
水生生物學(xué)報(bào) 2021年2期
范 淼 楊 威 孫 數(shù) 李 忠 張利紅* 張為民*
(1.中山大學(xué)生命科學(xué)學(xué)院水生經(jīng)濟(jì)動(dòng)物研究所, 廣州 510275; 2.仙桃市忠善黃鱔工廠(chǎng)化繁育基地, 仙桃 433012;3.中國(guó)水產(chǎn)科學(xué)研究院長(zhǎng)江水產(chǎn)研究所, 武漢 430223)
脊椎動(dòng)物性別決定和分化發(fā)育機(jī)制一直是生物學(xué)研究熱點(diǎn)。目前研究表明, 脊椎動(dòng)物性別決定方式包括基因決定性別和環(huán)境因素決定性別, 性別分化方向呈現(xiàn)可塑性[1]。魚(yú)類(lèi)具有雌雄異體和雌雄同體等多種性別表現(xiàn)形式, 性別分化可塑性高, 是研究脊椎動(dòng)物性別決定和分化發(fā)育的良好模型。同時(shí), 很多魚(yú)類(lèi)在生長(zhǎng)和體型等方面呈現(xiàn)性別二態(tài)型, 通過(guò)性控育種實(shí)現(xiàn)單性化養(yǎng)殖將產(chǎn)生重大的經(jīng)濟(jì)效益[2,3], 而魚(yú)類(lèi)性別決定和分化機(jī)制研究是實(shí)現(xiàn)魚(yú)類(lèi)性控育種的基礎(chǔ)。
黃鱔(Monopterus albusZuiew)隸屬合鰓魚(yú)目,合鰓魚(yú)科, 黃鱔屬, 是目前我國(guó)重要的特色淡水養(yǎng)殖魚(yú)類(lèi), 在湖北、安徽和湖南等地廣泛養(yǎng)殖。但是,黃鱔養(yǎng)殖的苗種仍主要是人工捕獲的野生黃鱔。由于過(guò)度捕撈, 近年來(lái)黃鱔自然資源日益減少, 嚴(yán)重影響黃鱔規(guī)模化人工養(yǎng)殖業(yè)的可持續(xù)發(fā)展。早在1944年, 我國(guó)脊椎動(dòng)物性別決定研究先驅(qū)劉建康先生發(fā)現(xiàn)黃鱔是一種雌性先熟的性轉(zhuǎn)變魚(yú)類(lèi)[3]。由于幼體階段性腺分化發(fā)育的單向性(雌性)和成體階段性腺自然可塑性(雌性轉(zhuǎn)變?yōu)樾坌?, 黃鱔已成為研究脊椎動(dòng)物性別分化發(fā)育機(jī)制的重要魚(yú)類(lèi)模型[4]。黃鱔自然性轉(zhuǎn)變規(guī)律和調(diào)控機(jī)制的闡明不僅具有很高的理論價(jià)值, 也將為其規(guī)模化人工繁育技術(shù)研發(fā)提供重要的支撐。
Liu[5]通過(guò)分析在重慶北碚野外采集的黃鱔性腺組織學(xué)結(jié)構(gòu)及其性別比例, 首次提出黃鱔具有自然性轉(zhuǎn)變現(xiàn)象。隨后, Liem[6]分析了印度尼西亞萬(wàn)隆地區(qū)野外捕獲和實(shí)驗(yàn)室養(yǎng)殖的黃鱔性腺組織學(xué)結(jié)構(gòu)和性別比例, Chan和Phillips[7]分析了中國(guó)香港和廣東地區(qū)的黃鱔性腺組織學(xué)結(jié)構(gòu)和性別比例, 這些研究都證實(shí)了黃鱔存在自然性轉(zhuǎn)變。Chan等[8]通過(guò)活體解剖技術(shù)跟蹤同一條黃鱔性腺組織學(xué)變化, 進(jìn)一步直接證實(shí)了黃鱔存在自然性轉(zhuǎn)變。雄性黃鱔都是從雌性黃鱔通過(guò)間性階段轉(zhuǎn)變而來(lái), 沒(méi)有發(fā)現(xiàn)雄魚(yú)再轉(zhuǎn)變?yōu)殚g性或雌性的現(xiàn)象[6,8]。
在黃鱔卵巢向精巢轉(zhuǎn)變時(shí), 最早出現(xiàn)的組織結(jié)構(gòu)變化是生殖褶增厚, 并在生殖褶內(nèi)圍出現(xiàn)零星的雄性生殖細(xì)胞; 隨著雄性生殖細(xì)胞的增殖和雌性生殖細(xì)胞的退化, 最終卵巢被精巢所取代, 雌性黃鱔轉(zhuǎn)變?yōu)樾坌渣S鱔[9]。關(guān)于雌性黃鱔在什么發(fā)育階段開(kāi)始性轉(zhuǎn)變, 有些學(xué)者認(rèn)為是在雌鱔性成熟產(chǎn)卵后發(fā)生[5,7,10—12]。通過(guò)分析年齡、體長(zhǎng)和體重等與黃鱔性轉(zhuǎn)變的相關(guān)性, 發(fā)現(xiàn)黃鱔性別與年齡和體長(zhǎng)關(guān)系較密切[13]。有研究表明, 雌性黃鱔2齡后開(kāi)始性轉(zhuǎn)變[6,13]。由于生存環(huán)境和/或地理位置的不同, 不同地區(qū)的黃鱔發(fā)生性轉(zhuǎn)變時(shí)體長(zhǎng)不同, 印度尼西亞萬(wàn)隆地區(qū)黃鱔在體長(zhǎng)為16 cm左右時(shí)開(kāi)始性轉(zhuǎn)變[6],中國(guó)香港和廣東地區(qū)的黃鱔在體長(zhǎng)為20 cm左右時(shí)開(kāi)始性轉(zhuǎn)變[7], 中國(guó)天津地區(qū)的黃鱔在體長(zhǎng)為40 cm左右時(shí)發(fā)生性轉(zhuǎn)變[14]。
增殖細(xì)胞核抗原(Proliferating cell nuclear antigen, 簡(jiǎn)稱(chēng)PCNA)是真核細(xì)胞DNA合成所必需的一種核蛋白。在細(xì)胞增殖過(guò)程中, PCNA表達(dá)量在G1晚期開(kāi)始上升, S期達(dá)到高峰, G2期下降[15,16]。PCNA作為細(xì)胞增殖的分子標(biāo)記, 常用于分析魚(yú)類(lèi)性腺細(xì)胞的增殖。美洲花鱔(Synbranchus marmoratusBloch)是一種先雌后雄的性轉(zhuǎn)變魚(yú)類(lèi), 其性轉(zhuǎn)變?cè)缙谛韵俳Y(jié)締組織中Leydig細(xì)胞的PCNA免疫活性很高[17]。在孵化180d的黑棘鯛(Acanthopagrus schlegeliiBleeker)早期精巢中, 發(fā)現(xiàn)生殖細(xì)胞中有PCNA免疫陽(yáng)性信號(hào)[18]。本文分析了實(shí)驗(yàn)室內(nèi)從受精卵或幼苗開(kāi)始養(yǎng)殖至不同時(shí)段的黃鱔性腺發(fā)育狀況, 并對(duì)性腺進(jìn)行了PCNA免疫組織化學(xué)分析, 旨在闡明黃鱔性轉(zhuǎn)變的基本規(guī)律, 以及間性性腺中各種細(xì)胞增殖與性轉(zhuǎn)變的關(guān)聯(lián)。
1 材料與方法
1.1 實(shí)驗(yàn)黃鱔及其飼養(yǎng)
5月齡黃鱔為仙桃市忠善黃鱔工廠(chǎng)化繁育基地(湖北仙桃)2019年7月孵出的黃鱔苗, 于2019年9月運(yùn)回本實(shí)驗(yàn)室(廣東廣州)飼養(yǎng); 12月齡和34月齡黃鱔分別為本實(shí)驗(yàn)室于2018年5—6月和2016年5—6月孵化的黃鱔苗養(yǎng)成。在自然光照和溫度條件下, 參照何智等[19]報(bào)道的方法, 實(shí)驗(yàn)室內(nèi)黃鱔飼養(yǎng)于直徑為 80 cm 的塑料盆中, 投喂生物餌料(先期為水蚯蚓, 5月齡后為黃粉蟲(chóng)), 投喂量約為體重的5%。養(yǎng)殖盆中放置1—2條灰色塑料管(直徑6.5 cm,長(zhǎng)度37 cm), 以便黃鱔藏匿, 模仿其生態(tài)習(xí)性。每天換水, 并根據(jù)生長(zhǎng)情況適時(shí)分盆, 控制養(yǎng)殖密度[20]。5月齡和12月齡黃鱔一直飼養(yǎng)在實(shí)驗(yàn)室內(nèi), 34月齡黃鱔在性腺活檢手術(shù)前1個(gè)月轉(zhuǎn)移至室外水泥池(1 m×1 m×1 m)中飼養(yǎng)。在整個(gè)飼養(yǎng)和實(shí)驗(yàn)過(guò)程中,每天上、下午觀(guān)察和記錄黃鱔生存狀態(tài)。
1.2 黃鱔性腺樣品的采集和分析
通過(guò)性腺活檢手術(shù)[8]獲得黃鱔性腺樣品。黃鱔腸道位于腹腔的腹面, 腎臟位于腹腔的背面, 緊貼背壁, 性腺位于腸道與腎臟之間, 呈管狀, 以腸系膜與腸道相連。黃鱔性腺活檢手術(shù)基本操作如下: 使用麻醉劑MS-222(500 mg/L; Sigma, A5040, MO,USA)麻醉實(shí)驗(yàn)黃鱔。將麻醉后的黃鱔置于實(shí)驗(yàn)臺(tái),腹面朝上, 用碳鋼手術(shù)刀片(11號(hào); 上海浦東金環(huán)醫(yī)療用品股份有限公司, 中國(guó)上海)于麻醉黃鱔泄殖孔前0.3—0.5 cm處腹中線(xiàn)偏右上方縱向開(kāi)口0.5—1 cm, 小心把性腺與腸系膜剝離, 剪取適量性腺樣品后, 于腹腔內(nèi)埋植經(jīng)75%酒精消毒的電子標(biāo)簽(廣州市洪騰條碼技術(shù)有限公司, 中國(guó)廣州), 然后用Δ1/2醫(yī)用縫合針(6×14; 巢湖市賓雄醫(yī)療器械有限公司, 中國(guó)安徽)和非吸收型外科縫合線(xiàn)(4-0; 揚(yáng)州源康醫(yī)療器械有限公司, 中國(guó)江蘇)縫合傷口, 一般縫合3—5針即可。在縫合處涂抹紅霉素藥膏(廣東恒建制藥有限公司, 中國(guó)廣東)后, 將術(shù)后黃鱔置于曝氣水中蘇醒。如要對(duì)同一條黃鱔個(gè)體進(jìn)行多次活檢手術(shù), 下一次手術(shù)是在上一次手術(shù)位置前(頭部方向)1—2 mm處實(shí)施。
30條5月齡黃鱔[體重(15.3±2.9) g, 體長(zhǎng)(25.7±1.2) cm]于2019年12月間進(jìn)行性腺活檢手術(shù); 17條12月齡黃鱔[體重(9.1±3.1) g, 體長(zhǎng)(20.3±2.5) cm]和94條34月齡黃鱔[體重(25.5±9.1) g, 體長(zhǎng)(30.5±3.7) cm]于2019年3—5月間進(jìn)行性腺活檢手術(shù)。于2019年3—4月間, 對(duì)94條34月齡黃鱔中的36條雌魚(yú)[體重(20.8±6.7) g, 體長(zhǎng)(29.2±3.4) cm]性腺發(fā)育進(jìn)行了為期1個(gè)月的跟蹤研究。
通過(guò)性腺活檢手術(shù)剪取長(zhǎng)約0.5 cm黃鱔性腺組織, 置于波恩氏液中固定24h, 然后按常規(guī)石蠟組織學(xué)切片流程處理。切片厚度為 4—5 μm, 蘇木精-伊紅(HE)染色, 中性樹(shù)膠封片, 使用Nikon顯微鏡(E800, Nikon, Japan)進(jìn)行觀(guān)察和拍照。
1.3 PCNA免疫組織化學(xué)定位
黃鱔性腺組織石蠟切片經(jīng)脫蠟和水化后, 用3%過(guò)氧化氫(無(wú)水甲醇∶30%過(guò)氧化氫∶ddH2O=8∶1∶1, 現(xiàn)配現(xiàn)用)在室溫下處理15min, 滅活內(nèi)源性過(guò)氧化物酶; 用PBS(137 mmol/L NaCl, 2.7 mmol/L KCl, 10 mmol/L Na2HPO4, 2 mmol/L KH2PO4; pH=7.4)浸洗3次(每次5min)后, 組織切片放入預(yù)熱(95℃)的0.01 mol/L檸檬酸鹽緩沖液(pH= 6.0)處理15min, 進(jìn)行抗原修復(fù); 取出玻片放涼至室溫后, PBS浸洗3次(每次5min), 滴加適量的正常山羊血清, 在室溫濕潤(rùn)的條件下孵育30min, 封閉非特異性結(jié)合位點(diǎn); 在封閉后的切片上滴加PCNA即用型抗體(MAB-0145鼠單抗, 克隆號(hào)PC10, 福州邁新生物科技有限公司, 中國(guó)), 4℃過(guò)夜孵育, 再于室溫下孵育3h。在一抗孵育完后, 用PBS浸洗切片3次(每次5min), 滴加適量免疫組化信號(hào)放大劑(EliVisionTMSuper, KIT-9922,福州邁新生物科技有限公司), 室溫孵育15min, 經(jīng)PBS浸洗3次(每次5min), 再滴加適量高敏型酶標(biāo)抗鼠/兔聚合物(EliVisionTMSuper, KIT-9922, 福州邁新生物科技有限公司), 室溫孵育15min后, PBS浸洗3次(每次5min), DAB(DAB-1031, 福州邁新生物科技有限公司, 中國(guó))顯色。陰性對(duì)照以PBS代替PCNA抗體, 其余操作一致。用Nikon顯微鏡(E800,Nikon, Japan)觀(guān)察切片和拍照。
1.4 數(shù)據(jù)處理
實(shí)驗(yàn)黃鱔體重和體長(zhǎng)等生物學(xué)參數(shù)采用SPSS統(tǒng)計(jì)軟件進(jìn)行分析。數(shù)據(jù)表示為平均值±標(biāo)準(zhǔn)差(Mean±SD)。
2 結(jié)果
2.1 未經(jīng)產(chǎn)卵過(guò)程的間性和雄性黃鱔
實(shí)驗(yàn)所用黃鱔是從幼魚(yú)(指5月齡魚(yú))或受精卵(指12月齡和34月齡魚(yú))開(kāi)始飼養(yǎng), 每天記錄生活狀況。在飼養(yǎng)過(guò)程中, 未發(fā)現(xiàn)有任何雌鱔發(fā)育至腹部膨大呈半透明狀態(tài), 也未觀(guān)察到養(yǎng)殖盆中有黃鱔卵粒或吐泡筑巢行為, 因此判斷實(shí)驗(yàn)黃鱔沒(méi)有經(jīng)歷產(chǎn)卵過(guò)程。
用石蠟組織切片和HE染色技術(shù), 分析30條5月齡、17條12月齡和94條34月齡黃鱔的性腺發(fā)育狀況。結(jié)果表明, 在5月齡黃鱔中, 雌性占比66.7%, 間性占比13.3%, 雄性占比20.0%(圖1和表1); 在12月齡黃鱔中, 雌性占比82.4%, 雄魚(yú)占比17.6%(圖2和表1); 在34月齡黃鱔中, 雌性占比78.7%, 間性占比12.8%, 雄性占比8.5%(圖3和表1)。
不同年齡段黃鱔的性別與其個(gè)體大小關(guān)系分析表明, 在5月齡和34月齡的黃鱔中, 間性和雄性個(gè)體較雌性偏大(圖4A和4C), 但12月齡黃鱔中雄性個(gè)體較雌性偏小(圖4B)。
2.2 三齡雌鱔不經(jīng)產(chǎn)卵過(guò)程發(fā)生性轉(zhuǎn)變現(xiàn)象
對(duì)94條34月齡黃鱔中的36條雌性黃鱔性腺發(fā)育進(jìn)行了跟蹤研究。結(jié)果表明, 在1個(gè)月的跟蹤實(shí)驗(yàn)過(guò)程中, 30條雌鱔依舊為雌性(圖5A2和A2’), 但有6條雌鱔變成為間性(圖5B2和B2’), 性轉(zhuǎn)變發(fā)生比例為16.7%。在黃鱔間性性腺中, 生殖褶增厚, 出現(xiàn)間質(zhì)細(xì)胞, 及早期雄性生殖細(xì)胞包括精原細(xì)胞和精母細(xì)胞(圖5B2’)。
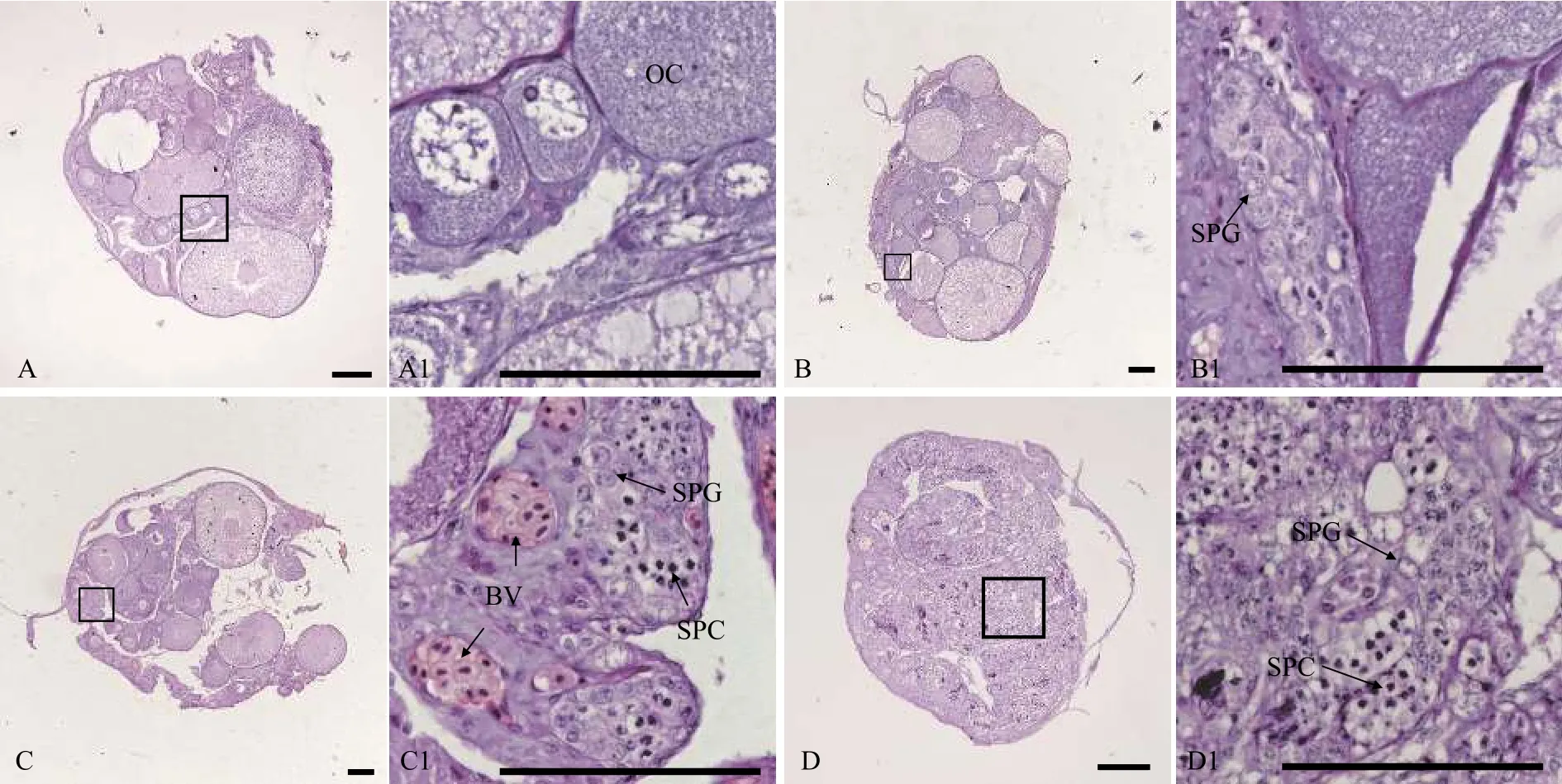
圖1 五月齡黃鱔的不同性別狀態(tài)性腺組織結(jié)構(gòu)Fig.1 Gonadal histology of 5-month-old ricefield eels of different sexual stages

表1 不同年齡段黃鱔的性別比例Tab.1 Sex ratios of ricefield eels at different ages
2.3 黃鱔性轉(zhuǎn)變過(guò)程中性腺PCNA免疫組織化學(xué)分析
對(duì)同一黃鱔個(gè)體的不同發(fā)育階段的性腺進(jìn)行PCNA免疫組織化學(xué)分析, 結(jié)果表明, 在卵黃發(fā)生期黃鱔卵巢中, 卵母細(xì)胞的細(xì)胞核及其外周部分濾泡層細(xì)胞核具有明顯的PCNA免疫陽(yáng)性信號(hào)(圖6A1);卵巢中生殖褶很薄, 僅見(jiàn)個(gè)別細(xì)胞的細(xì)胞核具有PCNA免疫陽(yáng)性信號(hào)(圖6A2)。
在間性早期黃鱔性腺中, 除卵母細(xì)胞的細(xì)胞核及其外周部分濾泡層細(xì)胞核可觀(guān)察到明顯的PCNA免疫活性信號(hào)(圖6B1)外, 增厚的生殖褶中也出現(xiàn)明顯的PCNA免疫信號(hào)。這些PCNA免疫信號(hào)主要分布在生殖褶兩類(lèi)細(xì)胞, 其中一類(lèi)為間質(zhì)細(xì)胞, 另一類(lèi)為雄性生殖細(xì)胞, 包括精原細(xì)胞和初級(jí)精母細(xì)胞(圖6B2)。
在間性晚期黃鱔性腺中, 退化卵母細(xì)胞上未見(jiàn)PCNA免疫信號(hào)(圖6C1), 增厚的生殖褶中可見(jiàn)不同發(fā)育階段的雄性生殖細(xì)胞, 其中精原細(xì)胞和初級(jí)精母細(xì)胞有明顯的PCNA免疫信號(hào)(圖6C2), 部分間質(zhì)細(xì)胞上也有PCNA免疫信號(hào), 但次級(jí)精母細(xì)胞上未觀(guān)察到PCNA免疫信號(hào)(圖6C1和C2)。
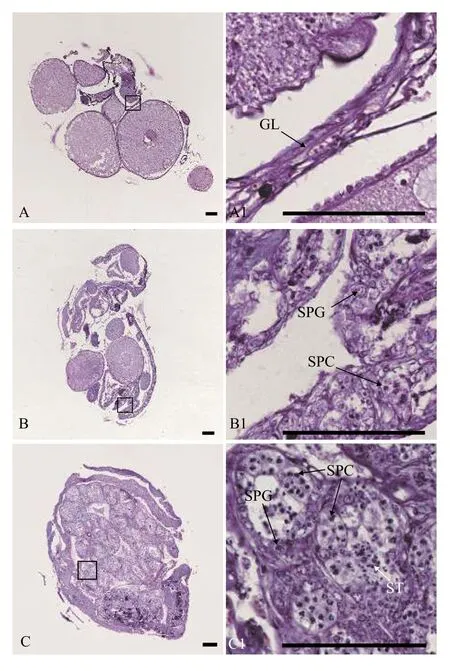
圖3 34月齡黃鱔的不同性別狀態(tài)性腺組織結(jié)構(gòu)Fig.3 Gonadal histology of 34-month-old ricefield eels of different sexual stages
以PBS代替PCNA抗體, 免疫組織化學(xué)分析結(jié)果表明, 黃鱔性腺切片中未呈現(xiàn)任何免疫陽(yáng)性信號(hào)(圖6D1和D2), 提示上述黃鱔性腺切片中PCNA免疫信號(hào)具有特異性。
3 討論
3.1 黃鱔性轉(zhuǎn)變與年齡、體長(zhǎng)、體重的關(guān)系
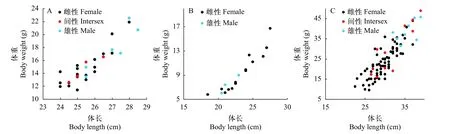
圖4 不同年齡段黃鱔的性別與體重和體長(zhǎng)的關(guān)系Fig.4 The relationship between sex, body weight, and body length of ricefield eels at different ages

圖5 兩條34月齡黃鱔的性腺組織結(jié)構(gòu)在跟蹤1個(gè)月前后的對(duì)比Fig.5 Tracing for gonadal changes of two 34-month-old ricefield eels in a one-month period
已有研究表明, 黃鱔在2齡之前都為雌性, 2齡后開(kāi)始性轉(zhuǎn)變[6,13]。本文分析了實(shí)驗(yàn)室內(nèi)從受精卵或幼苗養(yǎng)殖至不同年齡黃鱔的性腺發(fā)育狀況, 發(fā)現(xiàn)在5月齡黃鱔中就存在有間性和雄性個(gè)體, 提示實(shí)驗(yàn)室內(nèi)養(yǎng)殖的黃鱔在5月齡之前就已經(jīng)開(kāi)始性轉(zhuǎn)變,比之前的報(bào)道至少提前了1.5年。

圖6 黃鱔性轉(zhuǎn)變過(guò)程中性腺PCNA免疫組織化學(xué)分析Fig.6 PCNA immunohistochemistry in gonads of ricefield eels during sex change
在實(shí)驗(yàn)室養(yǎng)殖的5月齡、12月齡和34月齡的黃鱔中, 都有一定比例的不同性別的個(gè)體, 表明在同一個(gè)群體中黃鱔性轉(zhuǎn)變年齡存在個(gè)體差異。劉修業(yè)等[13]也發(fā)現(xiàn)3—5齡黃鱔性別處于過(guò)渡期, 性別過(guò)渡期達(dá)2年之久。那么, 黃鱔在什么年齡完成性轉(zhuǎn)變, 即在什么年齡全部變?yōu)樾坌? 對(duì)于這個(gè)問(wèn)題, 目前仍沒(méi)有定論。王良臣等[14]在野外采集的6齡和7齡黃鱔樣品中沒(méi)有發(fā)現(xiàn)雌性和間性個(gè)體, 全部為雄性。但是, 本實(shí)驗(yàn)室有3條養(yǎng)殖8年的黃鱔, 其性腺仍處于間性階段(數(shù)據(jù)未顯示)。這些不同研究之間黃鱔性別與年齡關(guān)系的差異可能是因?yàn)閷?shí)驗(yàn)黃鱔生存環(huán)境不同所致, 而同一群體中黃鱔個(gè)體性轉(zhuǎn)變年齡的差異可能是由遺傳因素所決定的[8]。
除年齡外, 黃鱔性別與體長(zhǎng)關(guān)系也比較密切[13]。與其他研究類(lèi)似, 本研究表明, 在5月齡和34月齡黃鱔中, 間性和雄性個(gè)體較雌性個(gè)體偏大。在5月齡和34月齡黃鱔中, 間性黃鱔最小個(gè)體體長(zhǎng)分別為24.5和26.5 cm, 比較接近鄱陽(yáng)湖和洞庭湖黃鱔性轉(zhuǎn)變的臨界體長(zhǎng)30 cm[21,22]和粵港地區(qū)黃鱔性轉(zhuǎn)變臨界體長(zhǎng)20 cm[7], 而與天津[14]和信陽(yáng)地區(qū)[23]黃鱔性轉(zhuǎn)變臨界體長(zhǎng)(40.0 cm 左右)差異較大。與萬(wàn)隆地區(qū)黃鱔性轉(zhuǎn)變臨界體長(zhǎng)(16 cm左右)[6]也差異較大。這些不同地區(qū)黃鱔性轉(zhuǎn)變臨界體長(zhǎng)差異可能與所在區(qū)域的緯度和氣候條件相關(guān), 低緯度地區(qū), 黃鱔生長(zhǎng)發(fā)育較快, 性成熟早, 發(fā)生性轉(zhuǎn)變也早, 性轉(zhuǎn)變時(shí)體長(zhǎng)也較小[11]。
本研究所觀(guān)察的12月齡黃鱔的體重和體長(zhǎng)比5月齡偏小, 這可能是由于這批黃鱔在7月齡時(shí)發(fā)生疾病, 出現(xiàn)大規(guī)模死亡現(xiàn)象, 攝食和生長(zhǎng)都受到嚴(yán)重影響。與5月齡和34月齡雄性黃鱔不同, 12月齡黃鱔的雄性個(gè)體較雌性偏小, 導(dǎo)致這些12月齡雄性黃鱔個(gè)體偏小的原因目前仍不清楚。Chan和 Phillips[7]曾報(bào)道有一些小個(gè)體的雌性黃鱔卵巢中卵母細(xì)胞數(shù)量少, 并有明顯的精巢組織出現(xiàn), 推測(cè)它們的雌性階段很短。因此, 由這些小個(gè)體的雌魚(yú)性轉(zhuǎn)變產(chǎn)生的雄魚(yú), 其個(gè)體也應(yīng)該偏小。
3.2 產(chǎn)卵并非雌鱔性轉(zhuǎn)變的必經(jīng)事件
研究者通常認(rèn)為, 雌性黃鱔的性轉(zhuǎn)變是在經(jīng)過(guò)一次或多次性成熟產(chǎn)卵后發(fā)生的[7,11,24,25]。本研究分析了實(shí)驗(yàn)室內(nèi)從受精卵或幼苗養(yǎng)殖至不同年齡黃鱔的性腺發(fā)育狀況。在養(yǎng)殖過(guò)程中, 沒(méi)有發(fā)現(xiàn)黃鱔有產(chǎn)卵現(xiàn)象或繁殖行為。前期研究表明, 實(shí)驗(yàn)室內(nèi)養(yǎng)殖的雌性黃鱔1齡時(shí)可發(fā)育至性成熟[19]。這些實(shí)驗(yàn)室內(nèi)養(yǎng)殖的雌性黃鱔之所以不產(chǎn)卵, 可能是由于室內(nèi)養(yǎng)殖條件不適合其產(chǎn)卵和繁殖。性腺組織切片結(jié)果表明, 在94條34月齡黃鱔中, 有12條間性個(gè)體, 占比12.8%; 30條5月齡黃鱔中, 有4條間性個(gè)體, 占比13.3%, 這些結(jié)果提示, 雌性黃鱔不經(jīng)過(guò)產(chǎn)卵過(guò)程也可以發(fā)生性轉(zhuǎn)變。另外, 本研究對(duì)36條34月齡雌性黃鱔的性腺發(fā)育進(jìn)行了跟蹤觀(guān)察, 發(fā)現(xiàn)16.7%的黃鱔在這一跟蹤過(guò)程中發(fā)生性轉(zhuǎn)變, 進(jìn)一步表明, 產(chǎn)卵并不是雌性黃鱔性轉(zhuǎn)變必須要經(jīng)歷的事件。
就目前所掌握的資料來(lái)看, “黃鱔產(chǎn)卵后性轉(zhuǎn)變”的觀(guān)點(diǎn)最早由Chan和Phillips[7]于1967年提出的。但是, 早在1963年, Liem[6]從黃鱔受精卵孵化出黃鱔幼苗后養(yǎng)殖, 第30個(gè)月取樣時(shí)發(fā)現(xiàn)有雌性和間性黃鱔個(gè)體, 第36個(gè)月時(shí)取樣發(fā)現(xiàn)有雌性、間性和雄性黃鱔個(gè)體。但是, 該論文沒(méi)有說(shuō)明這些30月齡的間性黃鱔是否在實(shí)驗(yàn)室內(nèi)經(jīng)歷了產(chǎn)卵過(guò)程, 因此, 不能判斷產(chǎn)卵過(guò)程是否是黃鱔性轉(zhuǎn)變必經(jīng)事件。在2004年, 周秋白等[21]報(bào)道, 懷卵未產(chǎn)雌鱔經(jīng)過(guò)1周年飼養(yǎng)后有11%的個(gè)體轉(zhuǎn)變?yōu)殚g性。這些結(jié)果似乎與“黃鱔產(chǎn)卵后性轉(zhuǎn)變”的觀(guān)點(diǎn)不一致。但是, 該論文并沒(méi)有提供這些懷卵未產(chǎn)雌鱔的生物學(xué)參數(shù)和年齡等信息, 也沒(méi)有提及這些懷卵未產(chǎn)雌鱔是否初次性成熟, 之前的年份是否曾經(jīng)歷產(chǎn)卵過(guò)程,因此, 論文的討論部分并沒(méi)有對(duì)“黃鱔產(chǎn)卵后性轉(zhuǎn)變”的觀(guān)點(diǎn)提出質(zhì)疑[21]。
那么, 雌性黃鱔產(chǎn)卵后是否就進(jìn)入間性階段發(fā)育? 答案是否定的。周秋白等[21]發(fā)現(xiàn), 已產(chǎn)卵黃鱔在第二年的雌性比例仍占51.1%。因此, 雌性黃鱔產(chǎn)卵過(guò)程與性轉(zhuǎn)變啟動(dòng)并沒(méi)有必然的因果關(guān)聯(lián)。
黃鱔性轉(zhuǎn)變的誘因目前仍不清楚。研究表明,環(huán)境因素如溫度和社會(huì)等級(jí)變化等影響?hù)~(yú)類(lèi)性別決定和分化[26,27]。應(yīng)激激素皮質(zhì)醇可能介導(dǎo)了溫度誘導(dǎo)魚(yú)類(lèi)雄性化和社會(huì)行為誘導(dǎo)雌雄同體魚(yú)類(lèi)性轉(zhuǎn)變的過(guò)程[28]。在雌性先熟的性轉(zhuǎn)變魚(yú)類(lèi)斜帶石斑魚(yú)中, 腹腔注射皮質(zhì)醇可誘導(dǎo)其從雌性轉(zhuǎn)變?yōu)樾坌訹29]。因此, 有關(guān)環(huán)境因素和皮質(zhì)醇等與黃鱔性轉(zhuǎn)變的關(guān)聯(lián)值得進(jìn)一步探討。
3.3 黃鱔是否有原發(fā)性雄性
研究發(fā)現(xiàn), 同屬合鰓目的美洲花鱔也具有先雌后雄的性轉(zhuǎn)變現(xiàn)象, 這種魚(yú)的雄性有2種來(lái)源, 即原發(fā)性雄性(不經(jīng)雌性階段直接發(fā)育為雄性)和次發(fā)性雄性(雌魚(yú)經(jīng)性轉(zhuǎn)變發(fā)育成雄魚(yú))[17,30]。次發(fā)性雄魚(yú)精巢是在卵巢結(jié)構(gòu)基礎(chǔ)上改造而來(lái), 因此精巢為單個(gè), 但是原發(fā)性雄魚(yú)精巢是成對(duì)存在的[17]。
在黃鱔種群中, 是否存在原發(fā)性的雄性可能因地理位置而異。Liem[30]報(bào)道, 在泰國(guó)、馬來(lái)西亞沙撈越、馬來(lái)半島和日本沖繩的黃鱔樣品中發(fā)現(xiàn)原發(fā)性雄性, 而在印度尼西亞萬(wàn)隆、日本Uda River和中國(guó)北碚的黃鱔樣品中沒(méi)有發(fā)現(xiàn)原發(fā)性雄性。與美洲花鱔原發(fā)性雄性一樣, 黃鱔原發(fā)性雄性的精巢也是成對(duì)存在, 由兩個(gè)精巢葉和連接兩者的結(jié)締組織構(gòu)成, 結(jié)締組織中央有一條精巢靜脈, 兩側(cè)各有一條輸精管, 分別與兩側(cè)的精巢葉對(duì)應(yīng); 而次發(fā)性雄性黃鱔的精巢保留了卵巢的解剖結(jié)構(gòu), 原來(lái)的卵巢腔轉(zhuǎn)變?yōu)榫驳妮斁? 原來(lái)的輸卵管的空間仍存在, 但可能在精巢中不行使功能。
Chan和Phillips[7]報(bào)道了香港和廣東地區(qū)的黃鱔精巢有2種結(jié)構(gòu)類(lèi)型, 一種為full-cord, 另一種為half-cord。前者整個(gè)生殖褶充滿(mǎn)精巢小葉, 而后者精巢小葉僅分布在生殖褶內(nèi)側(cè)。盡管Liem[30]推測(cè)含full-cord精巢的黃鱔為原發(fā)性雄性, 但是這種推測(cè)似乎缺乏證據(jù), 因?yàn)镃han和Phillips[7]在文中并沒(méi)有提出half-cord和full-cord兩種精巢在解剖結(jié)構(gòu)上有任何差異。
本研究在5月齡和12月齡黃鱔中都發(fā)現(xiàn)雄性個(gè)體, 占比分別為17.6%和20.0%, 比例較接近。組織切片分析表明, 這些雄性個(gè)體精巢中沒(méi)有任何退化的卵母細(xì)胞, 但解剖結(jié)構(gòu)上與卵巢類(lèi)似, 即有2條生殖褶, 2個(gè)性腺內(nèi)腔及一個(gè)性腺外腔[8], 這與Liem[30]所描述的黃鱔原發(fā)性雄性精巢結(jié)構(gòu)不同。因此, 本研究推測(cè)這些5月齡和12月齡雄性黃鱔也可能是通過(guò)性轉(zhuǎn)變所產(chǎn)生的, 其性轉(zhuǎn)變發(fā)生在卵巢發(fā)育早期階段。當(dāng)然, 另一種可能性也不能完全排除, 即這些5月齡和12月齡雄性黃鱔是原發(fā)性的雄性, 特別是12月齡的雄性黃鱔, 其個(gè)體較雌性黃鱔偏小, 與雌性轉(zhuǎn)化為雄性的自然性轉(zhuǎn)變過(guò)程中個(gè)體大小變化趨勢(shì)存在矛盾。如果這些雄性黃鱔是原發(fā)性雄性, 那么其精巢的早期發(fā)生與卵巢類(lèi)似, 而與Liem[30]所描述的黃鱔原發(fā)性雄性不同。因此, 中國(guó)本土黃鱔是否存在原發(fā)性雄性仍需進(jìn)一步探討。
3.4 黃鱔性轉(zhuǎn)變過(guò)程中精巢組織細(xì)胞的發(fā)生
黃鱔從雌性轉(zhuǎn)變?yōu)樾坌缘倪^(guò)程是性腺中的卵巢組織不斷被精巢組織所取代的過(guò)程[7], 因此性轉(zhuǎn)變過(guò)程伴隨著精巢組織體細(xì)胞和生殖細(xì)胞的增殖。黃鱔精巢體細(xì)胞和生殖細(xì)胞發(fā)生于性腺生殖褶內(nèi)側(cè), 其中的性原細(xì)胞(Gonocyte)在雌性階段就已存在, 性轉(zhuǎn)變開(kāi)始時(shí), 這些性原細(xì)胞增殖, 產(chǎn)生精原細(xì)胞, 同時(shí)間質(zhì)細(xì)胞也分化發(fā)育, 形成Leydig細(xì)胞[7,8]。在黃鱔間性早期性腺增厚的生殖褶中, 本研究也觀(guān)察到早期雄性生殖細(xì)胞包括精原細(xì)胞和精母細(xì)胞, 及精巢間質(zhì)細(xì)胞。這些間質(zhì)細(xì)胞呈卵圓形, 染色質(zhì)濃縮, 與核膜相鄰, 存在于精小囊之間[31]。但是, 由于缺乏相應(yīng)細(xì)胞的分子標(biāo)記, 黃鱔間性性腺中性原細(xì)胞、精原細(xì)胞和間質(zhì)細(xì)胞等的鑒定主要依據(jù)細(xì)胞形態(tài)和細(xì)胞在組織中所處的位置, 黃鱔性逆轉(zhuǎn)初期性腺組織細(xì)胞學(xué)變化仍很不清晰。
增殖細(xì)胞核抗原(Proliferating Cell Nuclear Antigen, 簡(jiǎn)稱(chēng)PCNA)是真核細(xì)胞DNA合成所必需的一種核蛋白, 在不同物種間具有高度的保守性, 已被用作多種組織中細(xì)胞增殖的生物標(biāo)志物[32—34]。本研究應(yīng)用免疫組織化學(xué)方法, 分析了卵巢、間性早期和間性晚期黃鱔性腺中PCNA免疫活性分布。發(fā)現(xiàn)PCNA免疫活性信號(hào)分布在黃鱔卵巢和間性早期性腺的卵母細(xì)胞, 間性晚期性腺的精原細(xì)胞和初級(jí)精母細(xì)胞等生殖細(xì)胞, 及卵巢中部分顆粒細(xì)胞中,這與上述這些性腺細(xì)胞具有較強(qiáng)的增殖能力相一致, 進(jìn)一步提示商品化的PCNA抗體可用于分析黃鱔性腺細(xì)胞增殖。
在間性早期黃鱔性腺的生殖褶中, 除精原細(xì)胞和初級(jí)精母細(xì)胞為PCNA免疫陽(yáng)性外, 精巢間質(zhì)細(xì)胞也含有PCNA免疫陽(yáng)性信號(hào)。在美洲花鱔性轉(zhuǎn)變?cè)缙诘男韵俳Y(jié)締組織中, 精巢間質(zhì)細(xì)胞也呈現(xiàn)很強(qiáng)的PCNA免疫信號(hào)[17]。這些結(jié)果提示, 精巢間質(zhì)細(xì)胞和精原細(xì)胞的增殖可能與黃鱔性轉(zhuǎn)變的啟動(dòng)密切相關(guān)。但是, 這些間質(zhì)細(xì)胞和精原細(xì)胞的起源仍有待進(jìn)一步研究。因此, 黃鱔性轉(zhuǎn)變?cè)缙谛韵僦芯搀w細(xì)胞和生殖細(xì)胞相關(guān)分子標(biāo)記的開(kāi)發(fā), 結(jié)合PCNA免疫組化分析, 將能更好地揭示這些精巢體細(xì)胞和生殖細(xì)胞的動(dòng)態(tài)變化, 有助于闡明黃鱔性轉(zhuǎn)變調(diào)控機(jī)制。
致謝:
感謝中山大學(xué)生命科學(xué)學(xué)院崔鑫、李曦、張瀾馨、劉敏琦、何智、許星和張有為同學(xué)協(xié)助實(shí)驗(yàn)黃鱔飼養(yǎng)管理。
