FAT/CD36表達及轉位在有氧運動改善老年小鼠骨骼肌胰島素敏感性中的作用
2021-04-06 13:40:00孫婧瑜蘇亞娟秦黎黎董靜梅
中國體育科技 2021年2期
孫婧瑜,蘇亞娟,秦黎黎,董靜梅
關鍵字:衰老;運動干預;脂肪酸轉位酶;胰島素信號通路
隨著社會老齡化的發展,老年人糖尿病的發病率急劇上升(Evans et al.,2013)。研究表明,衰老能夠誘導胰島素抵抗的發生(Hamanoue et al.,2017)。胰島素抵抗是高血壓、肥胖、2型糖尿病、非酒精性脂肪肝等代謝性疾病的主要危險因素。骨骼肌作為能量代謝的主要器官,隨著年齡的增長,骨骼肌胰島素敏感性下降(Dagdeviren et al.,2017;Hamanoue et al.,2017)。因此,骨骼肌被認為是衰老誘導胰島素抵抗的主要部位(Biswas et al.,2016)。
近年來,有研究發現脂質代謝紊亂是衰老誘導骨骼肌胰島素敏感性下降的重要因素(Biswas et al.,2016)。長鏈脂肪酸(long-chainfatty acids,LCFAs)從脂肪組織運輸到骨骼肌線粒體需要經過一系列脂肪酸轉運載體的調控。其中,脂肪酸轉位酶(fatty acid translocase,FAT/CD36)廣泛存在于骨骼肌中(Nickerson et al.,2009),并且FAT/CD36向細胞膜的轉位功能可能是所有脂肪酸轉運載體中所特有的(Romic et al.,2017)。在人類肥胖模型及嚙齒類動物胰島素抵抗模型研究中發現,細胞膜FAT/CD36與脂肪酸攝取、甘油三酯堆積以及胰島素刺激下葡萄糖攝取的減少密切相關(Bonen et al.,2004)。因此,骨骼肌細胞膜FAT/CD36在調節脂肪酸轉運過程中可能起到了中心作用。有研究發現,與衰老相關的胰島素抵抗發生時,細胞膜FAT/CD36含量增多不是由于FAT/CD36合成的增加,而是由于永久固定于細胞膜上的FAT/CD36增多。這種永久固定于細胞膜上的FAT/CD36增多可能是導致2型糖尿病早期發病的主要因素;而線粒體功能障礙引起的過量脂質沉積可能是糖尿病后期發病的主要因素(Steinbusch et al.,2011)。因此,對骨骼肌FAT/CD36的表達及轉位機制進行研究可能是糖尿病防治的新思路。
由于運動的安全性、有效性和多重靶點作用等優勢,越來越多的研究聚焦其潛在的糖尿病防治機理。然而,運動對衰老誘導的胰島素抵抗骨骼肌FAT/CD36的作用及作用機理尚不明確。因此,本研究首先利用siRNA干擾技術,在C2C12細胞中進行FAT/CD36基因敲低,探討FAT/CD36缺乏對骨骼肌細胞胰島素信號通路的影響。其次,利用老年小鼠,探討運動對老年小鼠骨骼肌FAT/CD36總蛋白含量的影響。有研究發現,與FAT/CD36總蛋白相比,運動誘導的FAT/CD36向細胞膜的轉位功能可能對骨骼肌脂肪酸代謝更為重要(Mcfarlan et al.,2012)。電壓依賴性陰離子通道蛋白(voltage-dependent anion channel,VDAC)是線粒體外膜上含量極為豐富的孔狀蛋白,也是線粒體的重要標志物之一(Zhang et al.,2019);小窩蛋白(caveolins)在FAT/CD36向細胞膜脂筏錨定的過程中起到了重要的調控作用(Eyre et al.,2007)。因此,本研究進一步探討運動對老年小鼠骨骼肌FAT/CD36向細胞膜和/或線粒體膜轉位的影響及其在細胞膜脂筏中的定位。根據研究推測,運動可能通過減少FAT/CD36的表達、增加向細胞膜脂筏轉位的雙重作用,激活AKT/ERK信號通路,從而改善衰老誘導的骨骼肌胰島素敏感性。
1 實驗材料與方法
1.1 C2C12細胞培養及siRNA干擾
小鼠骨骼肌C2C12細胞系在6孔板內培養,以生長培養基(高糖DMEM、10%小牛血清、1%谷氨酸、1%青霉素/鏈霉素)培養2天后,改用分化培養基(高糖DMEM、2%馬血清、1%谷氨酸、1%青霉素/鏈霉素)進行分化培養4~6天。在C2C12細胞分化第3天時,換用無抗生素的分化培養基,分別對各組細胞進行siRNA干擾:siCont、siCD36-1、sicd36-2。siCont:5’-UUCUCCGAACGUGUCACGUTT-3’;siCD36-1:5’-GGAUGACAACUUCACAGUUTT-3’;siCD36-2:5’-CCACAUUUCCUACAUGCAA-TT-3’。
6孔板中每孔加入A液(150 μl Optimem+1.5 μl 20mol/L siRNA)和B液(150 μl Optimem+3 μl Lipofecta-mine RNAi-MAX)。A液混入B液后,將混合物于室溫放置5 min,加入C2C12細胞,于37℃培養箱孵育6 h后,換回分化培養基,繼續分化培養2天。然后,對細胞進行低糖DMEM及無血清饑餓過夜,并用含磷酸酶抑制劑的蛋白裂解液裂解細胞,用于后續檢測。
1.2 實驗動物與分組
8周齡C57BL/6雄性小鼠自由進食飲水,環境溫度20℃~24℃,相對濕度40%~60%,通風良好,晝夜12 h/12 h循環照明(所有操作均嚴格遵守同濟大學道德倫理委員會的要求)。小鼠自然飼養至56周齡后,隨機分為老年對照組(aging control,AC;n=10)和老年運動組(aging exercise,AE;n=10)。老年對照組無特殊運動訓練,進行16周中等強度運動訓練。
1.3 小鼠運動模型構建
小鼠進行適應性跑臺訓練1周,每周6次,每次訓練時間由20 min/次逐漸遞增至60 min/次,跑速由10 m/min逐漸遞增至20 m/min。第2周開始,小鼠跑臺訓練以20 m/min跑速,6次/周,60 min/次進行并持續16周。訓練時間固定在每周一至周六下午,周日休息。
1.4 腹腔注射葡萄糖耐量實驗
第72周末對小鼠進行腹腔注射葡萄糖耐量實驗(intraperitoneal glucose tolerance test,IPGTT)。小鼠禁食16 h后,腹腔注射葡萄糖溶液1 g/kg,并分別在腹腔注射0 min、15 min、30 min、60 min、120 min及180 min時進行取血。采用羅氏卓越精采型血糖儀及其配套試紙進行血糖測量,并繪制血糖曲線,計算血糖曲線下面積(area under the curve,AUC)(Sun et al.,2018)。
1.5 腹腔注射胰島素耐量實驗
第72周末對小鼠進行腹腔注射胰島素耐量實驗(intraperitoneal insulin tolerance test,IPGTT)。小鼠禁食16 h后,腹腔注射胰島素2 g/kg,并分別在腹腔注射0 min、15 min、30 min、60 min及120 min時進行取血。采用羅氏卓越精采型血糖儀及其配套試紙進行血糖測量,并繪制血糖曲線,計算血糖曲線下面積(AUC)(Sun et al.,2018)。
1.6 動物取材
實驗干預后,小鼠禁食12 h,自由飲水。選用7 mL/kg 20%戊巴比妥鈉對小鼠進行腹腔麻醉注射。主動脈取血后迅速分離雙側腓腸肌,進行RT-PCR、WB及免疫熒光相關指標檢測。剩余組織置于?80℃冰箱冷凍待用。
1.7 血脂檢測
甘油三酯(triglyceride,TG)、總膽固醇(total cholesterol,T-CHO)、高密度脂蛋白膽固醇(high density lipopro?tein cholesterol,HDL?C)及低密度脂蛋白膽固醇(low density lipoprotein chesterol,LDL-C)4項血脂指標按照相應試劑盒說明書進行酶聯免疫檢測儀檢測。
1.8 RNA的提取和RT-PCR
60~80 mg腓腸肌剪碎后加入500 ul Trizol(抽提總RNA。室溫靜置5 min后,加入100 μl氯仿。12 000 r/min,4℃離心15 min后吸取170 μl上清,并加入等體積異丙醇,室溫靜置 10 min。12 000 r/min,4℃離心10 min,棄上清。75%乙醇洗滌沉淀2次后,沉淀溶于10 μl RNase-free水中并測定OD值。取1 μg總RNA,按照cDNA合成試劑盒說明書進行反轉錄。
RT-PCR 反應體系為 10 μl,包括 100 ng cDNA、5 μl 2×FastStart Universal SYBR GreenMaster、0.5 μl Primer F/R(10 μmol/L)、4 μl RNase Free Water。表1為 PCR 引物正向和反向序列。反應條件為:起始模板預變性95℃,10 min;PCR循環反應95 ℃,10 s;60 ℃,10 s,72 ℃,10 s,共40個循環。用內參β-actin對樣品進行標準化,并用2-△△Ct方法計算mRNA的相對含量。

表1 熒光定量PCR引物序列Table 1 Primer Sequences for Fluorescence Quantitative PCR
1.9 免疫印跡
取60~80 mg腓腸肌置于含磷酸酶抑制劑的RIPA裂解液中,剪碎后充分勻漿(或C2C12細胞裂解液),冰上靜置30 min后,12 000/min,4℃離心10 min,取上清;BCA法測定蛋白樣品濃度后進行煮沸變性。蛋白樣品經過SDSPAGE電泳、轉膜、5%脫脂奶粉室溫封閉1 h后,一抗4℃孵育過夜。TBST洗膜3次,5 min/次,二抗室溫孵育1 h。采用ECL發光法檢測目標蛋白條帶,并用Image J軟件分析條帶灰度值。
1.10 免疫熒光
制備骨骼肌冰凍切片,置于4%多聚甲醛冰上固定60 min,封閉60 min后,分別加入CD36/Caveolin-1混合一抗(FAT/CD36,1:200;Caveolin-1,1:200),CD36/VDAC 混合一抗(FAT/CD36,1:200;VDAC,1:200),4 ℃孵育過夜。混合 Alexa Fluor 594(紅)、Alexa Fluor 488(綠)標記的二抗,室溫孵育60 min。滴加含DAPI的抗熒光淬滅劑封片。采用Zeiss LSM700激光共聚焦顯微鏡觀察并采集圖像,用ZEN分析軟件分析熒光強度及共定位程度。熒光強度反映FAT/CD36蛋白表達的變化情況,共定位程度反映FAT/CD36在胞內的轉位情況。
1.11 數據處理
實驗數據采用SPSS 20.0軟件處理,結果以平均數±標準誤(M±SE)表示。組間比較采用獨立樣本t檢驗,顯著性差異水平設置為P<0.05。
2 實驗結果
2.1 FAT/CD36缺乏對骨骼肌細胞胰島素信號通路AKT/ERK激活的影響
經siCD36干擾后的C2C12細胞,FAT/CD36蛋白表達水平顯著下降(圖1),說明本實驗中的FAT/CD36基因敲低效率較好。進一步研究發現,經siCD36干擾后的C2C12細胞,AKT/ERK磷酸化水平顯著增加,說明FAT/CD36基因缺乏能夠激活骨骼肌細胞胰島素信號通路。
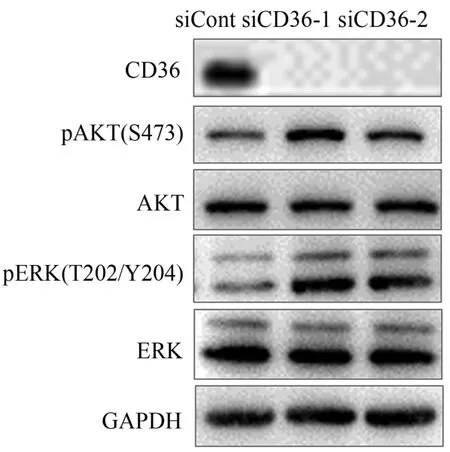
圖1 FAT/CD36缺乏對C2C12小鼠骨骼肌細胞相關蛋白表達水平的影響Figure 1. Effect of FAT/CD36 Deficiency on Related Protein Expression Levels in C2C12 Skeletal Muscle Cells
2.2 運動對老年小鼠糖耐量水平的影響
小鼠腹腔注射葡萄糖0 min、15 min、30 min、60 min、120 min、180 min后血液葡萄糖水平及IPGTT血糖曲線下面積(AUC)(圖2)。結果顯示,AE組AUC顯著低于AC組(P<0.05),說明運動能改善老年小鼠葡萄糖耐量。
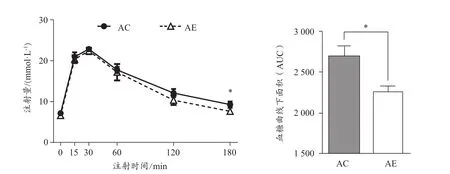
圖2 運動對老年小鼠葡萄糖耐量的影響Figure 2. Effect of Exercise Intervention on Glucose Tolerance inAged Mice
2.3 運動對老年小鼠胰島素耐量水平的影響
小鼠腹腔注射胰島素 0 min、15 min、30 min、60 min、120 min后血液葡萄糖水平及IPITT血糖曲線下面積(圖3)。結果顯示,AE組AUC顯著低于AC組(P<0.05),說明運動能改善老年小鼠胰島素耐量。
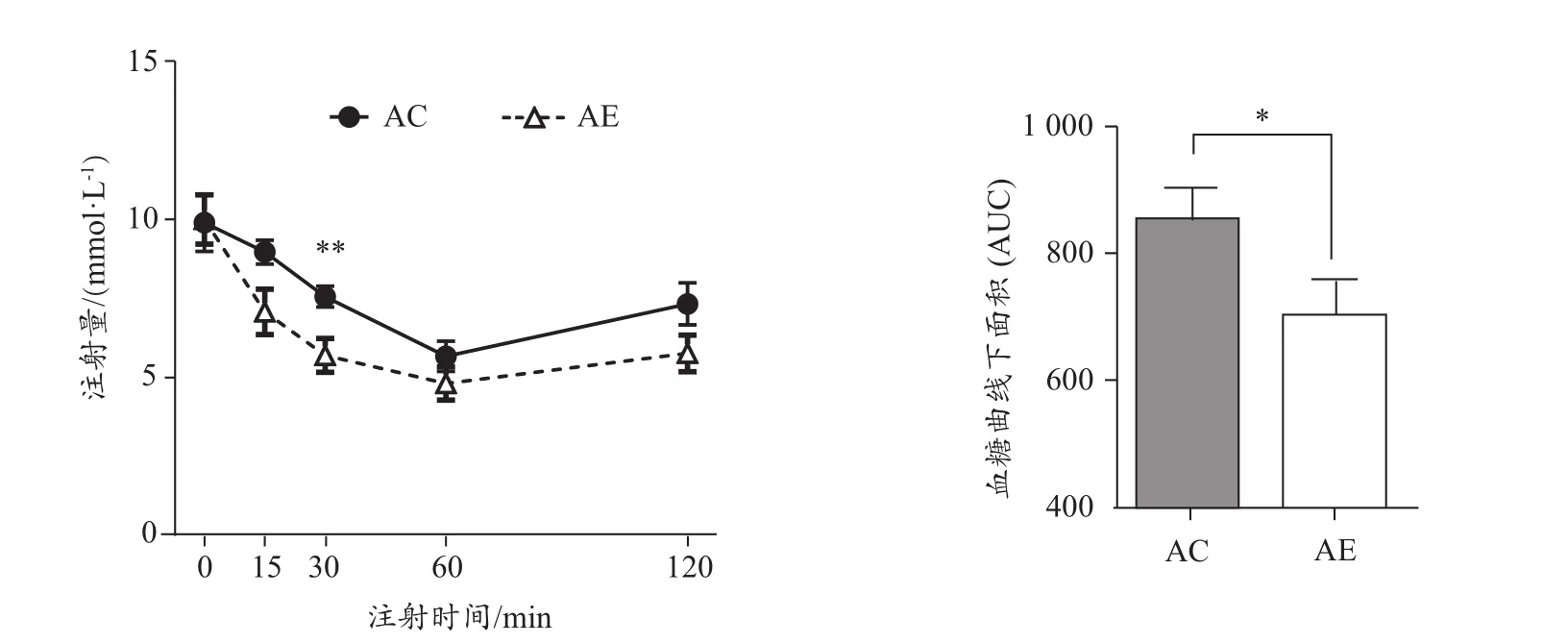
圖3 運動對老年小鼠胰島素耐量的影響Figure 3. Effect of Exercise Intervention on Insulin Tolerance inAged Mice
2.4 運動對老年小鼠血脂的影響
與AC組相比,AE組小鼠血清甘油三酯(TG)、總膽固醇(T-CHO)及低密度脂蛋白膽固醇(LDL-C)含量無顯著性差異;與AC組相比,AE組小鼠高密度脂蛋白膽固醇(HDL-C)水平顯著增加(P<0.01),說明運動能顯著改善老年小鼠HDL-C含量(圖4)。

圖4 運動對老年小鼠血脂的影響Figure 4. Effect of Exercise Intervention on Blood Lipid Content inAged Mice
2.5 運動對老年小鼠骨骼肌胰島素信號通路激活的影響
與AC組相比,AE組小鼠骨骼肌AKT/ERK磷酸化水平顯著增加(P<0.05,圖5),說明中等強度運動訓練能顯著改善老年小鼠骨骼肌胰島素敏感性。
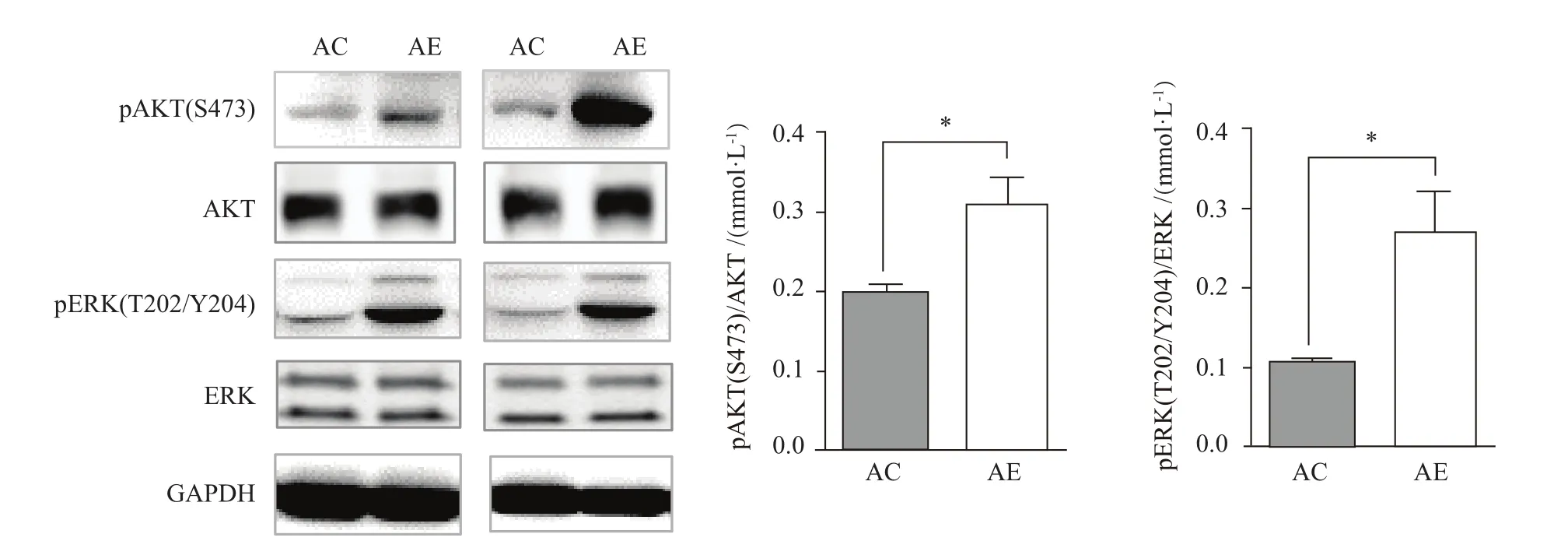
圖5 運動對老年小鼠骨骼肌胰島素信號通路激活的影響Figure 5. Effect of Exercise on theActivation of Insulin Signaling Pathway in the Skeletal Muscle ofAged Mice
2.6 運動對老年小鼠骨骼肌FAT/CD36及其他脂肪酸轉運載體mRNA表達水平的影響
與AC組相比,AE組小鼠骨骼肌脂肪酸結合蛋白(fatty acid binding protein,FABPpm)、脂肪酸轉運蛋白(fatty acid transport protein 4,FATP4)mRNA表達水平未見顯著變化,而FAT/CD36、肉堿脂酰轉移酶-1(carnitine acyltransferase-1,CPT-1)mRNA表達水平顯著降低(P<0.05,圖6),說明與其他脂肪酸轉運載體相比,FAT/CD36、CPT-1在運動改善老年小鼠骨骼肌胰島素敏感性中可能發揮了更重要的作用。與AC組相比,AE組小鼠骨骼肌過氧化物酶增殖物激活受體γ共化合物1α(peroxisome proliferator activated receptorγcoactivator-1α,PGC-1α)和過氧化物酶體增殖物激活受體γ(peroxisome proliferatoractivated receptor-γ,PPAR-γ)mRNA表達水平未見顯著性變化,說明PGC-1α、PPARγ可能不是運動調控FAT/CD36表達變化的主要轉錄因子。
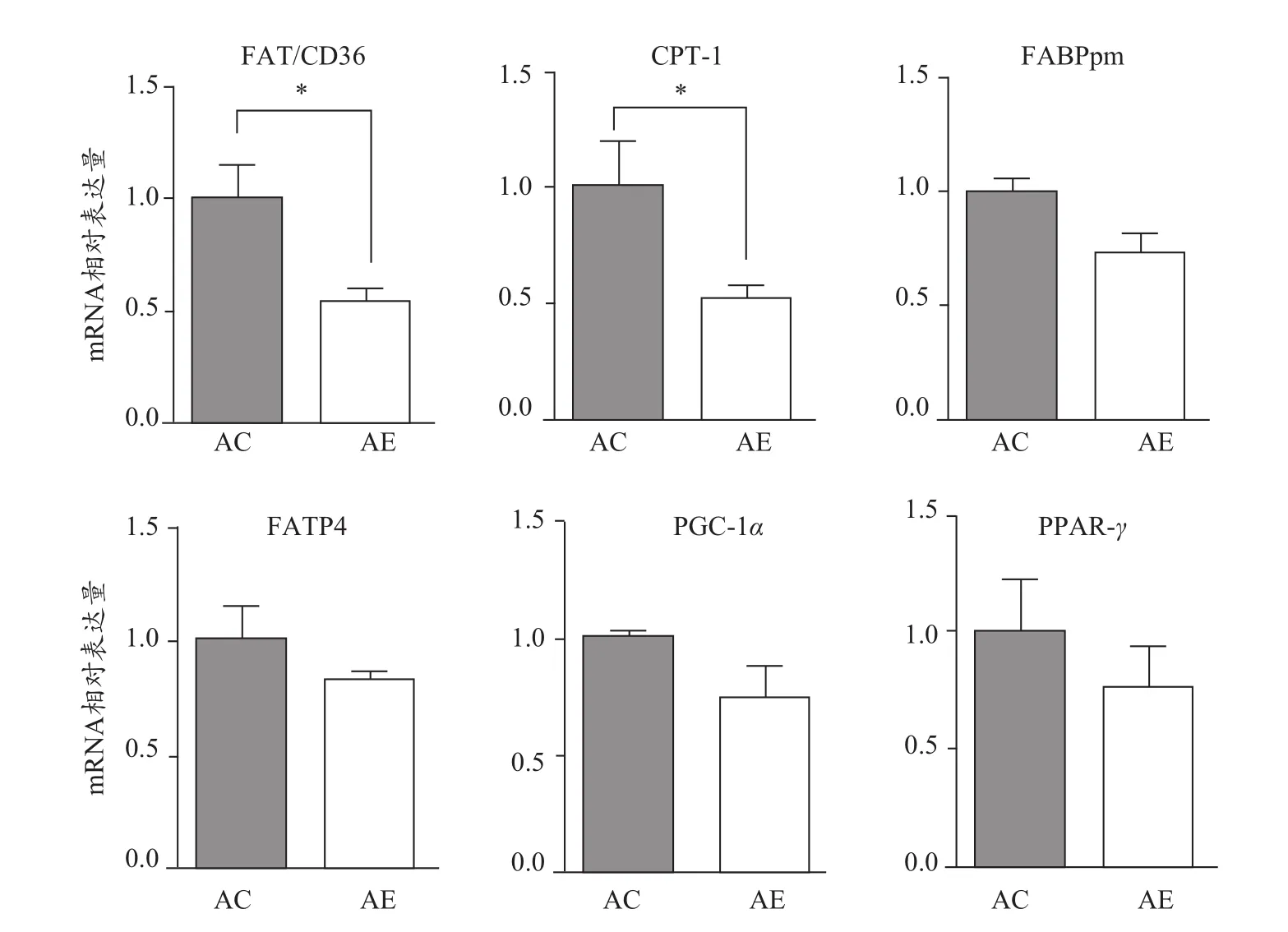
圖6 運動對老年小鼠骨骼肌脂肪酸轉運載體mRNA表達水平的影響Figure 6. Effect of Exercise on mRNAExpression Levels of FattyAcid Transporters in the Skeletal Muscle ofAged Mice
2.7 運動對老年小鼠骨骼肌FAT/CD36蛋白表達水平的影響
與AC組相比,AE組小鼠骨骼肌FAT/CD36的蛋白表達水平顯著減少(P<0.05,圖7),說明運動能顯著降低老年小鼠骨骼肌FAT/CD36的蛋白表達水平。
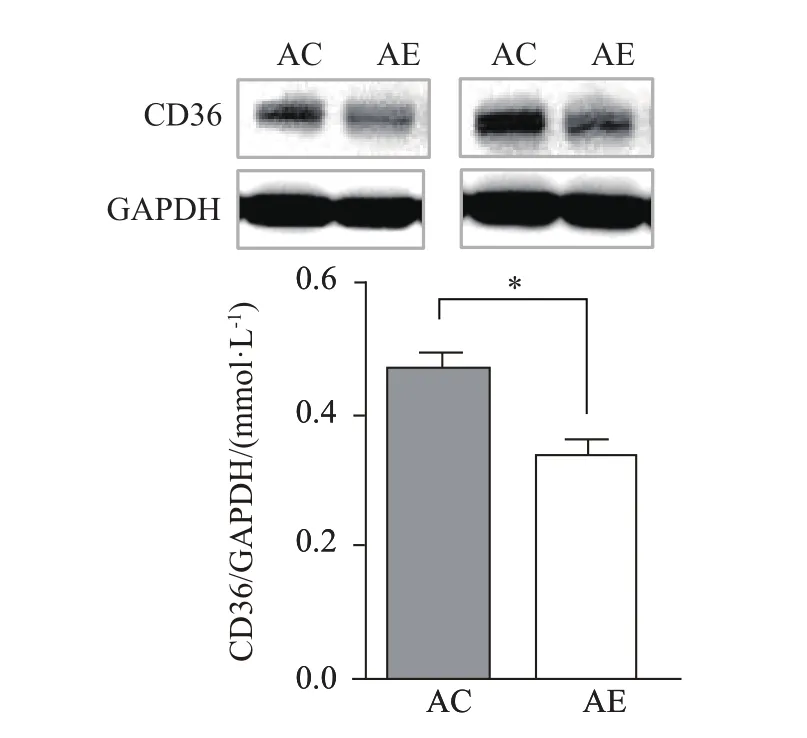
圖7 運動對老年小鼠骨骼肌FAT/CD36蛋白表達水平的影響Figure 7. Effect of Exercise on FAT/CD36 Protein Expression Levels in the Skeletal Muscle ofAged Mice
2.8 運動對老年小鼠骨骼肌FAT/CD36轉位功能的影響
小鼠骨骼肌組織CD36/Caveolin-1免疫熒光雙標檢測結果顯示(圖8),FAT/CD36被標記為綠色熒光,Caveolin-1被標記為紅色熒光,二者的共定位重疊區域為黃色,細胞核被DAPI標記為藍色。與AC組相比,AE組FAT/CD36綠色熒光強度顯著減少(P<0.05),說明運動能減少老年小鼠骨骼肌FAT/CD36的蛋白表達水平;AC組顯示,黃色共定位區域明顯,說明FAT/CD36存在于細胞膜脂筏中;與AC組相比,AE組黃色共定位程度進一步增加,說明運動能夠進一步誘導老年小鼠骨骼肌FAT/CD36向細胞膜轉位,并定位于細胞膜脂筏中。
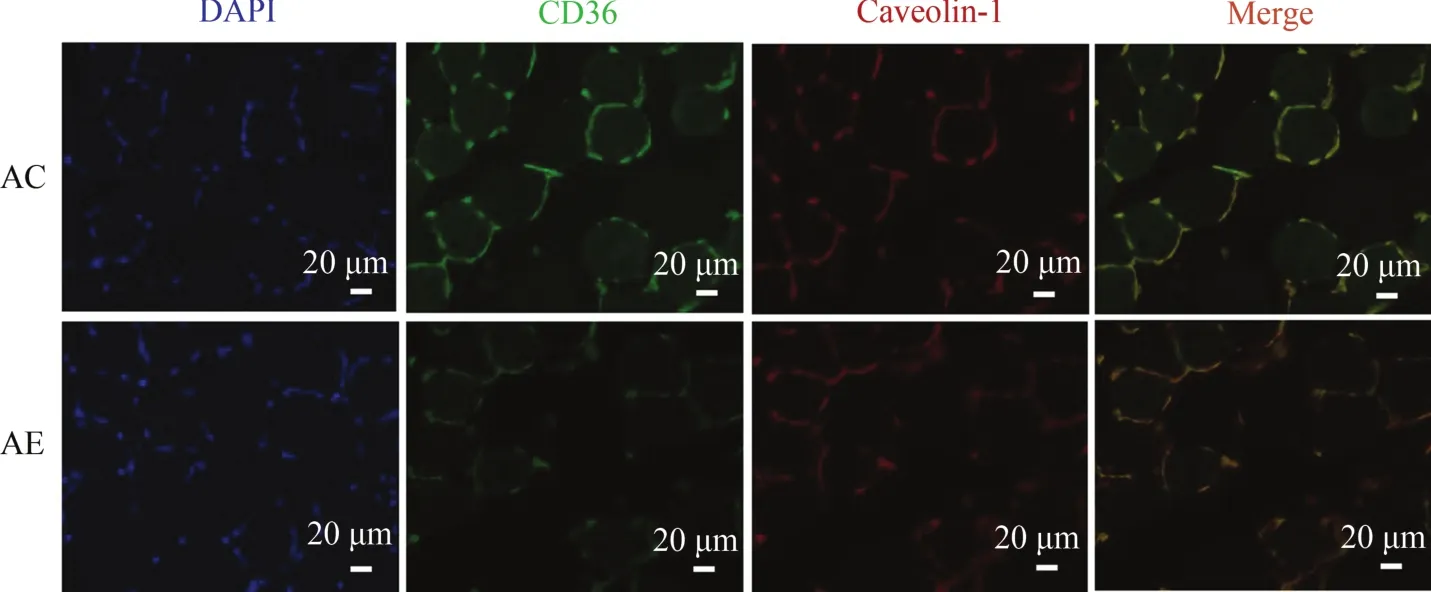
圖8 運動誘導老年小鼠骨骼肌FAT/CD36向細胞膜轉位Figure 8. Exercise Induced FAT/CD36 Trafficking to Plasma Membrane in the Skeletal Muscle ofAged Mice
2.9 運動未能誘導老年小鼠骨骼肌FAT/CD36向線粒體膜轉位
小鼠骨骼肌組織CD36/VDAC免疫熒光雙標檢測結果顯示(圖9),FAT/CD36被標記為綠色熒光,VDAC被標記為紅色熒光,二者的共定位重疊區域為黃色,細胞核被DAPI標記為藍色。AC組顯示,黃色共定位區域不明顯,說明FAT/CD36不存在于線粒體膜上;與AC組相比,AE組黃色共定位程度未有明顯改變,說明運動干預亦未能誘導老年小鼠骨骼肌FAT/CD36向線粒體膜轉位。
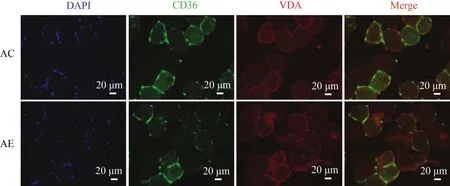
圖9 運動未誘導老年小鼠骨骼肌FAT/CD36向線粒體膜轉位Figure 9. Effect of Exercise Induced FAT/CD36 Trafficking to Mitochondrial Membrane in the Skeletal Muscle ofAged Mice
3 分析與討論
3.1 運動對老年小鼠骨骼肌胰島素敏感性的改善作用
胰島素抵抗是許多衰老相關疾病的獨立危險因素,如2型糖尿病、高血壓、動脈粥樣硬化等。骨骼肌是人體攝取和利用葡萄糖最重要的組織。隨著年齡的增長,骨骼肌的質量和功能出現衰退,導致胰島素抵抗的發生。因此,提高老年人骨骼肌胰島素敏感性是預防衰老相關疾病的重要措施。然而,除了運動干預之外,目前尚未有成熟的骨骼肌衰退治療方法。本研究結果顯示,中等強度運動訓練能顯著改善老年小鼠血糖、血脂及骨骼肌胰島素敏感性。
衰老導致過量脂肪酸流入骨骼肌細胞,脂肪酸在骨骼肌細胞內的過量沉積,導致骨骼肌脂肪酸氧化能力下降,線粒體生物合成發生障礙,線粒體功能損傷,慢肌纖維向快肌纖維轉變增多以及肌萎縮等變化,這些變化均與骨骼肌脂肪酸代謝紊亂有關,最終引起胰島素抵抗的發生(Oseph et al.,2012)。傳統觀點認為,外源性脂肪酸通過自由擴散的方式實現向骨骼肌細胞內的轉運;然而近年來研究發現,一些膜蛋白,如FAT/CD36、FABPpm及FATPs在協助脂肪酸跨過細胞膜,向細胞膜內轉運過程中起到了重要作用(Glatz et al.,2010)。尤其是FAT/CD36與骨骼肌細胞內脂肪酸的異位沉積關系密切(Bonen et al.,2004)。有研究結果表明,外源性脂肪酸通過激活FAT/CD36的轉位機制,對骨骼肌細胞內的脂肪酸攝取與氧化平衡起到了重要的調控作用(Samovski et al.,2015)。然而,靜息狀態下FAT/CD36可能不是最為重要的脂肪酸轉運載體,CPT-1可能更為重要;在運動等能量需求較大時,與其他脂肪酸轉運載體相比,FAT/CD36對骨骼肌脂肪酸代謝的調控作用更為明顯(Holloway et al.,2009)。因此,本研究著重探討FAT/CD36在運動改善老年小鼠骨骼肌胰島素敏感性中的作用及作用機制。
3.2 FAT/CD36蛋白表達水平在運動改善老年小鼠骨骼肌胰島素敏感性中的作用
研究顯示,胰島素抵抗骨骼肌中FAT/CD36的表達水平顯著增加(Bonen et al.,2004),而運動訓練對骨骼肌FAT/CD36表達水平的改變呈現多元化的調控。有研究顯示,6周高強度間歇訓練能顯著增加骨骼肌細胞FAT/CD36總蛋白表達水平(Talanian et al.,2010)。Kiens等(2004)發現,長期耐力訓練并不能增加人類股外側肌FAT/CD36總蛋白表達水平。這可能是因為FAT/CD36對運動的適應性變化,主要表現在FAT/CD36向細胞膜的轉位機制,而非總蛋白水平上的改變。另外,運動誘導的骨骼肌FAT/CD36總蛋白表達水平的變化可能存在時效性及運動干預方式上的差異,特別是在運動適應的初期,FAT/CD36誘導的骨骼肌脂肪酸調控機制較為復雜。骨骼肌FAT/CD36蛋白表達水平的增加可能是對運動訓練初期的一個暫時性的適應過程。隨著運動時間的延長,這種適應過程可能逐漸消失。本研究結果顯示,中等強度運動訓練顯著降低了老年小鼠骨骼肌組織FAT/CD36及CPT-1的mRNA表達,然而FABPpm及FATP4脂肪酸轉運載體的mRNA表達水平未見顯著性變化。該結果與Palacios等(2009)研究相一致,說明了與其他脂肪酸轉運載體相比,FAT/CD36及CPT-1在運動改善骨骼肌胰島素敏感性中可能發揮了重要的調控作用。另外,與FAT/CD36 mRNA表達水平變化相一致,16周中等強度運動訓練顯著降低了胰島素抵抗骨骼肌組織FAT/CD36的蛋白表達水平。
PGC-1α含量的增加不僅能夠誘導線粒體的生物發生,促進脂肪酸氧化代謝,還可以增加FAT/CD36的含量(Benton et al.,2010)。本研究結果顯示,16周中等強度運動訓練能夠顯著降低老年小鼠骨骼肌組織FAT/CD36的含量,但未能降低PGC-1α及PPARγ的mRNA表達水平,說明運動誘導的衰老小鼠骨骼肌FAT/CD36表達水平的變化可能不完全通過PGC-1α及PPARγ轉錄因子的調控。其他途徑對老年小鼠骨骼肌FAT/CD36表達水平的調控機制,需要進一步研究。
3.3 FAT/CD36轉位在運動改善老年小鼠骨骼肌胰島素敏感性中的作用
FAT/CD36普遍分布于骨骼肌細胞膜及胞質儲存囊泡中。研究顯示,機械收縮或胰島素刺激均能誘導細胞膜FAT/CD36含量的增加,而骨骼肌細胞內FAT/CD36總蛋白含量并沒有增加(Koonen et al.,2005)。由此推測,FAT/CD36在細胞膜和胞漿儲存體內可能是以循環的形式存在。當FAT/CD36的胞內轉位機制發生障礙時,細胞膜上的FAT/CD36便無法正常回收至胞質儲存囊泡,導致過量外源性脂肪酸的攝入,從而導致了骨骼肌脂肪酸代謝異常(Bonen et al.,2004)。但是對于FAT/CD36是否存在于線粒體膜上,尚存爭議。Lopaschuk等(2010)認為,FAT/CD36存在于線粒體膜上,并通過調節脂肪酸進入線粒體的數量,對脂肪酸氧化代謝進行調控。Holloway等(2007)認為,FAT/CD36既不存在于線粒體膜上,又不能通過調節脂肪酸進入線粒體的數量來對脂肪酸氧化進行調控。本研究結果顯示,FAT/CD36高表達于老年小鼠骨骼肌細胞膜中,而線粒體膜上未見表達。進一步研究發現,FAT/CD36向細胞膜的轉位功能是運動過程中脂肪酸氧化代謝調節的重要機制(Mcfarlan et al.,2012)。本研究結果與該結論相一致,即16周有氧運動能夠進一步誘導老年小鼠骨骼肌FAT/CD36向細胞膜的轉位,然而運動未能誘導老年小鼠骨骼肌FAT/CD36向線粒體膜的轉位。這可能是因為運動過程中需要大量的能量供應,外源性LCFA可以為骨骼肌收縮提供能量底物,而FAT/CD36可能主要在LCFA進入骨骼肌細胞膜過程中起到重要作用,在LCFA進入線粒體膜過程中的作用不如其他脂肪酸轉運蛋白,如FABPpm、CPT-1等。
3.4 FAT/CD36向細胞膜脂筏上的定位在運動改善老年小鼠骨骼肌胰島素敏感性中的作用
衰老誘導的胰島素抵抗伴有胰島素信號通路上的多個分子表達或活化水平的異常,如胰島素與受體的結合能力下降、胰島素受體、胰島素受體底物-1(insulin receptor substrate-1,IRS-1)的酪氨酸磷酸化以及磷脂酰肌醇-3激酶(phosphatidylinositol 3-kinase,PI-3K)活化水平下降等,這些均被認為是衰老誘導骨骼肌胰島素抵抗發生的可能機制。但是,目前對胰島素信號通路分子表達或活化水平改變的上游機制尚不清楚。隨著脂筏的發現,人們開始關注小GTP結合蛋白Rab GTPases在胰島素抵抗中的作用。脂筏是細胞膜上含有特殊脂質及特殊蛋白質的微結構域(microdomains)。有研究證明,脂肪酸的跨膜轉運離不開脂筏結構域(Ehehalt et al.,2006),這就為探討有氧運動改善胰島素敏感性的作用機制提供了一個新視角。
小窩是脂質膜上直徑約為50~100 nm的囊泡凹陷結構,在上皮細胞、脂肪細胞和肌細胞的細胞膜上表達豐富(Lo et al.,2015)。脂筏、小窩能夠通過聚集某些轉運蛋白,如葡萄糖、膽固醇轉運蛋白,來調控營養物質的攝取(Ortegren et al.,2007)。研究顯示,相關的長鏈脂肪酸轉運蛋白與脂筏、小窩結構關系密切(Ehehalt et al.,2006)。因此,推測小窩蛋白可能參與了FAT/CD36在細胞膜上的定位及功能。FAT/CD36只有錨定在細胞膜表面的脂筏微結構域內,并與小窩蛋白共同作用,才能對脂肪酸的攝取進行調控(Ehehalt et al.2008);若在回收過程中,FAT/CD36未能正確定位于脂筏和/或小窩中,可能會影響脂肪酸的攝取及轉運。然而,運動能否誘導衰老小鼠骨骼肌FAT/CD36向細胞膜脂筏和/或小窩上的正確定位,尚不清楚。本研究結果顯示,運動能夠誘導衰老小鼠骨骼肌FAT/CD36向細胞膜轉位并準確定位于細胞膜上的脂筏中。脂筏被看作是一個信號募集、發生、轉導的平臺,含有多種信號分子(Philip et al.,2007)。因此,本研究結果進一步說明了運動可能通過增加FAT/CD36在細胞膜脂筏上的定位,增強了FAT/CD36與下游信號分子的相互作用,如Src家族蛋白激酶(Fyn、Lyn和Yes)(Yamada et al.,2010),從而激活了相關胰島素信號通路,進而對衰老誘導的骨骼肌胰島素敏感性進行調控。在今后的研究中,可以選取運動初期、運動中期以及運動末期等多個時間點,從而可以更清晰地反映出FAT/CD36的表達及轉位功能在運動適應中的變化趨勢,進一步說明FAT/CD36在運動防治衰老誘導的骨骼肌胰島素抵抗中的靶點價值。
4 結論
FAT/CD36在調節骨骼肌胰島素信號通路中具有重要作用,并且運動可能通過減少FAT/CD36的表達、增加向細胞膜脂筏轉位的雙重作用,激活AKT/ERK信號通路,從而改善衰老誘導的骨骼肌胰島素敏感性下降(圖10)。
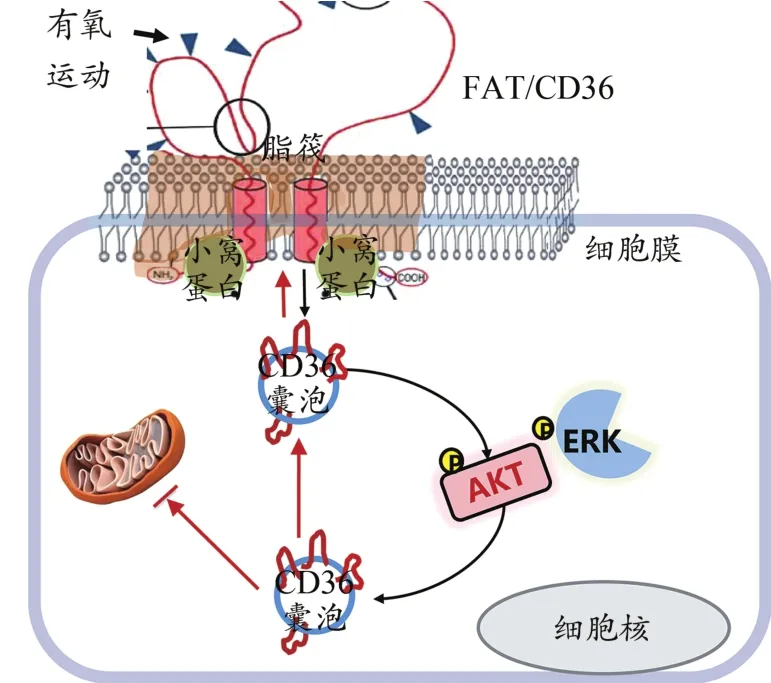
圖10 FAT/CD36在運動改善衰老骨骼肌胰島素敏感性中的作用及可能機制Figure 10. The Role of FAT/CD36 in Improving Insulin Sensitivity of Skeletal Muscle inAged Mice by Exercise
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
人人健康(2023年26期)2023-12-07 03:55:46
學苑創造·A版(2020年9期)2020-10-13 09:41:02
中國生殖健康(2019年2期)2019-08-23 08:12:10
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
中國衛生標準管理(2015年1期)2016-01-14 03:41:27
藥學與臨床研究(2015年4期)2015-06-05 11:35:51
中國醫藥科學(2015年15期)2015-02-27 12:32:27
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
中國全科醫學(2013年36期)2013-01-25 06:20:58
