農桿菌介導遺傳轉化獲得轉CP4基因秈稻的研究
2021-04-21 01:18:48曹含章呂尊富李飛飛
浙江農林大學學報 2021年2期
胡 歡,李 媛,丁 筠,曹含章,呂尊富,李飛飛
(浙江農林大學 農業與食品科學學院 浙江省農產品品質改良技術研究重點實驗室,浙江 杭州 311300)
在遺傳轉化獲得抗性植株時,轉化體的抗性篩選是遺傳轉化能否取得成功的關鍵步驟。通常在選擇培養基中加入合適種類和濃度的篩選劑,使其產生一定的篩選壓起到抗性篩選的作用。轉化體內選擇標記基因的表達產物可對特定篩選劑產生抗性,使轉化受體材料繼續保持正常的生長發育[1]。目前的研究中,卡那霉素、潮霉素等抗生素被普遍作為篩選劑使用[2-3]。但是由于水稻Oryza sativa胚性愈傷組織對抗生素具有生理抗性,以抗生素為選擇標記進行抗性篩選,不能起到很好的篩選效果,且經抗生素篩選后的轉化體在分化和再生階段易受抑制或產生白化苗[4-6]。以草甘膦作為篩選劑可以提高選擇的靈敏度,消除轉化體生理抗性對篩選結果的影響,克服了以往研究中抗生素篩選的局限性。可遺傳的草甘膦抗性基因突變率低,并可在后代中穩定表達,因此進行抗草甘膦作物的培育是可行的[7]。秈稻Oryza sativasubsp.indica和粳稻Oryza sativasubsp.japonica是栽培稻的2個亞種,隨著水稻遺傳轉化技術的發展,大部分粳稻品種已經建立了成熟的遺傳轉化體系,并成功引入抗蟲、抗病、生長發育調控等諸多有利基因[8]。而大多數秈稻品種組培特性不佳,愈傷組織誘導率低,繼代過程易褐化且分化再生頻率低,導致秈稻的遺傳轉化效率低,有的品種甚至難以轉化。尤其是對生產上廣泛推廣、農藝性狀優良的重要品種而言,其改良與育種進程受到嚴重限制[9]。CHAN等[10]于1992年嘗試利用農桿菌Agrobacterium tumefaciens介導法轉化秈稻幼根愈傷組織,對轉化體進行Southern印記雜交,結果表明:目的基因片段已成功轉入轉化體細胞中。后經酶活性檢測,目的基因可在轉化體中穩定表達。1994年,HIEI等[11]為建立高效穩定的農桿菌遺傳轉化體系,采用了“雙超元”載體,并通過在菌液添加乙酰丁香酮(As)活化Vir基因提高轉化效率等方法,推進了遺傳轉化技術在秈稻中的研究應用。目前,雖然已有轉抗草甘膦基因的秈稻遺傳轉化體系的報道,但是轉化效率低,還未建立一個高效的轉化體系[12]。基于此,本研究選取具有成功再生體系的秈稻‘中恢161’Oryza sativasubsp.indica‘Zhonghui 161’為材料,利用農桿菌介導法,轉入草甘膦抗性基因(CP4),探索適合的草甘膦質量濃度用于抗性篩選,并對農桿菌介導的轉化過程進行了合理優化,建立‘中恢161’農桿菌介導的轉化體系。
1 材料與方法
1.1 材料
秈稻‘中恢161’成熟胚;農桿菌菌株EHA105;含CP4基因的表達載體p1300-HC。
1.2 方法
1.2.1 成熟胚胚性愈傷組織的誘導和增殖 將成熟種子去殼,進行消毒[13],接種于誘導培養基R1[NB(N6+B5)+3.0 mg·L-12.4-D+0.5 g·L-1脯氨酸+0.1 g·L-1肌醇+0.3 g·L-1水解酪蛋白+30.0 g·L-1蔗糖+4.0 g·L-1Gelrite]上,接種20 粒·皿-1。放入培養條件為28 ℃,光照16 h/黑暗8 h的組培室中誘導培養。5~7 d后,可觀察到幼芽處有淡黃色愈傷組織,統計每皿的出愈數和出愈率。15 d后,剝下色澤鮮黃、結構緊密、生理狀態良好的胚性愈傷組織,分散平鋪于新鮮配制的胚性愈傷組織增殖培養基R1上進行增殖培養。繼代2~4次后,增殖并產生大量的胚性愈傷組織,可用做后期轉化的受體材料。
1.2.2 胚性愈傷組織的草甘膦敏感性測試 設置5組草甘膦質量濃度(100、200、300、400和500 mg·L-1),重復3次,設空白對照,接種20塊·皿-1。15 d后,觀察胚性愈傷組織的色澤、是否增殖等外觀形態,統計胚性愈傷組織褐化率,選出合適的草甘膦質量濃度范圍作為篩選壓。
1.2.3 胚性愈傷組織的遺傳轉化和抗性篩選 利用懸浮培養基R2(NB+0.5 g·L-1脯氨酸+0.1 g·L-1肌醇+0.3 g·L-1水解酪蛋白+30 g·L-1蔗糖+100 μmol·L-1乙酰丁香酮)將培養好的含CP4基因表達載體的農桿菌菌株EHA105稀釋至D(600)為0.5~0.8,用其侵染胚性愈傷組織[14]。將轉化好的胚性愈傷組織用無菌濾紙吸干多余的菌液,適當干燥后,用滅菌鑷子夾取愈傷組織分散地平鋪在鋪有1層無菌濾紙的共培養培養基 R3(NB+0.5 g·L-1脯氨酸+0.1 g·L-1肌醇+0.3 g·L-1水解酪蛋白+30.0 g·L-1蔗糖+100 μmol·L-1乙酰丁香酮+4.0 g·L-1Gelrite)上,20 塊·皿-1。于25 ℃培養室中暗培養2~3 d。取出共培養后的愈傷組織,用含100 mg·L-1羧芐青霉素的無菌蒸餾水清洗3~4次,直至清洗液澄清透明。適度干燥后,用鑷子夾取愈傷組織整齊均勻地平鋪在篩選培養基 R4(NB+3 mg·L-12.4-D+0.5 g·L-1脯氨酸+0.1 g·L-1肌醇+0.3 g·L-1水解酪蛋白+0.5 g·L-1谷氨酰胺+30.0 g·L-1蔗糖+4.0 g·L-1Gelrite+0.5 g·L-1頭孢霉素+不同質量濃度草甘膦)上。抗性篩選培養基中草甘膦質量濃度分別為300、350和400 mg·L-1。
1.2.4 分化、生根、移栽成活 將抗性愈傷組織系移至分化培養基R5(NB+0.5 mg·L-1NAA+3.0 mg·L-16-BA+0.5 g·L-1脯氨酸+0.1 g·L-1肌醇+0.3 g·L-1水解酪蛋白+0.5 g·L-1谷氨酰胺+30.0 g·L-1蔗糖+4.0 g·L-1Gelrite)上進行分化培養。約15~25 d,部分抗性愈傷組織長出綠點。待綠點進一步分化形成小苗,并長至2 cm左右時將其轉移至生根培養基R6(1/2NB+20.0 g·L-1蔗糖+0.1 g·L-1肌醇+8.0 g·L-1瓊脂)上生根培養。待幼苗生長出大量的茁壯根系,可將其從生根培養基中取出,小心洗凈其根系附著的培養基,置于培養箱中煉苗,增強幼苗對環境的適應性,1周后將健壯的幼苗移至大棚成活。
1.2.5 轉基因植株的分子檢測和CP4基因試紙條檢測蛋白表達 利用TPS法提取轉基因植株葉片DNA。利用CP4基因引物(CP4-F: TTCCTTTAGGATTTCAGCATCAGTG,CP4-R: TCCTTCATGTTCGGC GGTCTC)進行CP4基因的PCR擴增,目的片段大小為400 bp。擴增后的產物經質量分數為1%瓊脂糖凝膠電泳鑒定,統計陽性率。取陽性植株葉片,利用CP4基因表達檢測試紙條進行再生植株抗性檢測。
2 結果和分析
2.1 草甘膦抗性篩選最適質量濃度
圖1和圖2所示:‘中恢161’的胚性愈傷組織在不含草甘膦的培養基中可正常生長且大量增殖,未發生褐化現象;在含100 mg·L-1草甘膦的培養基中,絕大多數胚性愈傷組織可正常生長增殖,褐化率低,僅為5.00%,未起到選擇作用;在含200 mg·L-1草甘膦的培養基中褐化率為16.67%,選擇效果不明顯;在含300 mg·L-1草甘膦的培養基中,褐化率為41.67%,且與 200 mg·L-1相比差異顯著(P<0.05),選擇效果好,適合作為篩選壓;在含400 mg·L-1草甘膦的培養基中,大部分胚性愈傷組織發生褐化,少部分正常生長,褐化率為65.00%,選擇效果明顯;在含500 mg·L-1草甘膦的培養基中,胚性愈傷組織基本發生褐化,褐化率為91.67%,顯著高于其他質量濃度下的褐化率(P<0.05),說明選擇壓過大。結果表明:草甘膦質量濃度為300~400 mg·L-1時,胚性愈傷組織褐化率約50%,具有很好的篩選效果。
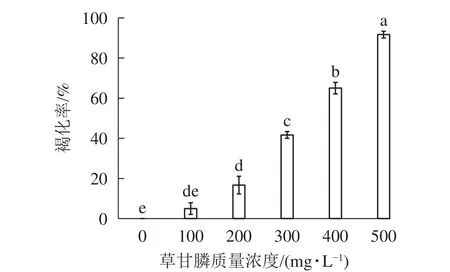
圖1 秈稻‘中恢161’胚性愈傷組織在不同質量濃度草甘膦培養基中培養20 d后的褐化率Figure 1 Browning rate of O.sativa subsp.indica ‘Zhonghui 161’ callus cultured in different concentrations of glyphosate for 20 days

圖2 秈稻‘中恢161’胚性愈傷組織在不同質量濃度草甘膦培養基中培養20 d后的褐化情況Figure 2 Browning rate of O. sativa subsp. indica ‘Zhonghui 161’ callus cultured in different concentrations of glyphosate for 20 days
2.2 抗性愈傷組織和轉基因植株的分子檢測
轉化后的胚性愈傷組織在含有300、350和400 mg·L-1草甘膦的選擇培養基上進行抗性篩選,分別獲得200、113、和84塊抗性愈傷組織,提取抗性愈傷組織DNA進行CP4基因的PCR檢測,陽性愈傷組織的PCR擴增產物經電泳可獲得長度為400 bp的條帶,與預期相符,表明CP4基因已成功整合到轉化體內。進行3個草甘膦質量濃度抗性篩選后愈傷組織陽性率分別為40.16%、61.72%和84.04%。共獲得67株再生植株,提取再生植株葉片DNA進行CP4基因的PCR檢測。其中陽性植株43株,再生植株陽性率為64.18%(圖3)。
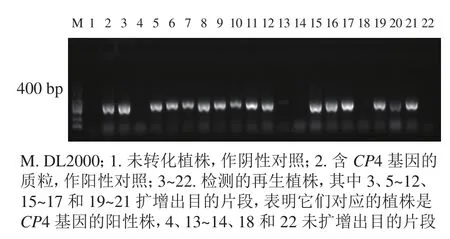
圖3 秈稻‘中恢161’再生植株CP4基因的PCR檢測Figure 3 PCR result of CP4 gene from glyphosate-resistance plants
2.3 CP4 基因試紙條檢測蛋白表達
抗性檢測結果(圖4)表明:檢測的43株PCR陽性植株中,有25株表現為CP4基因表達,表達率為58.13%。
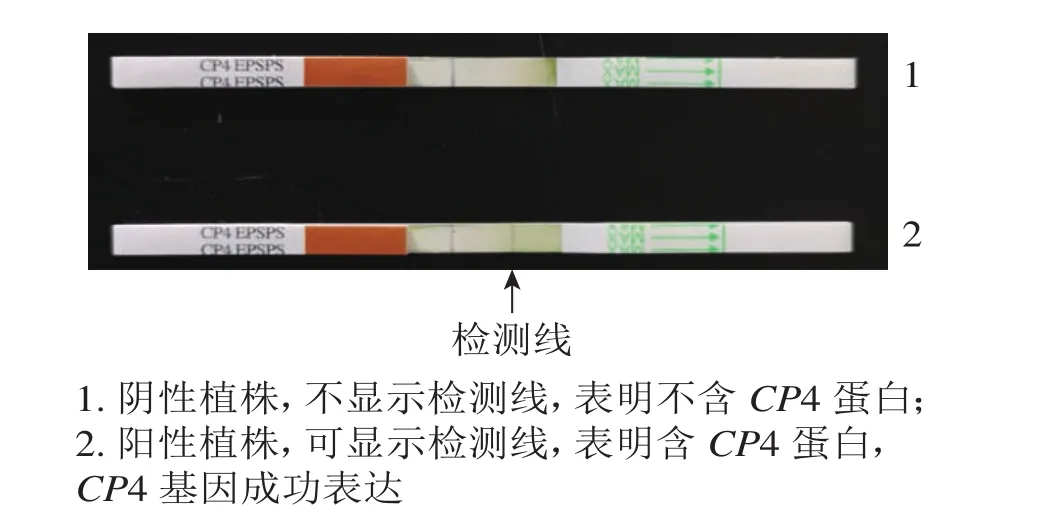
圖4 CP4蛋白活性試紙條檢測Figure 4 Protein activity of CP4 by strip test
2.4 ‘中恢 161’成熟胚遺傳轉化體系的建立
遺傳轉化再生過程如圖5所示:對秈稻‘中恢161’成熟胚進行胚性愈傷組織誘導,約7 d可誘導出胚性愈傷組織(圖5A)。胚性愈傷組織進行2~4次繼代增殖(圖5B),約40 d后進行遺傳轉化。轉化后的胚性愈傷組織在選擇培養基上進行抗性篩選(圖5C~D),一段時間后,抗性愈傷組織系會出現增殖(圖5E)。約50 d后,抗性愈傷組織于R5培養基上進行分化培養,約15~25 d,長出綠點(圖5F)。1個月左右,長出小苗(圖5G)。小心取出轉移至R6培養基進行生根培養(圖5H)。約15 d,幼苗長出大量的茁壯根系,將其從培養基中移出,小心洗凈根部培養基。置于培養箱中煉苗,煉苗1周后可移至大棚成活。對再生植株進行CP4基因的PCR檢測,保留陽性植株。從誘導胚性愈傷組織至獲得抗草甘膦再生植株的整個過程需要4~6個月。

圖5 CP4基因轉化秈稻‘中恢161’胚性愈傷組織的各個階段Figure 5 Various stages of transforming CP4 gene into embryogenic callus of O.sativa subsp. indica ‘Zhonghui 161’
3 結論與討論
3.1 草甘膦抗性篩選及其優勢
本研究建立了以草甘膦抗性基因CP4為選擇標記的‘中恢161’遺傳轉化體系。非轉化體的EPSPS酶活性較低,草甘膦可與S3P形成EPSPS-S3P-草甘膦復合體而競爭性抑制EPSPS酶活性,植物體內蛋白質合成受阻,生長受到抑制,不能正常生長分化[15]。而轉化體抗草甘膦基因CP4的表達產物EPSPS酶具有高催化活性和低草甘膦親和力不易與草甘膦結合,故能夠進行正常的生長分化。因此,通過草甘膦篩選可獲得轉抗草甘膦基因CP4的再生植株。相較于以抗生素抗性基因為選擇標記,草甘膦抗性基因不僅能作為篩選標記也能作為目的基因,使受體植物獲得除草劑抗性,而且草甘膦比潮霉素等抗生素便宜[7]。不同植物細胞對草甘膦的抗性存在差異,選擇合適的草甘膦質量濃度作為抗性篩選的篩選壓是影響轉化效率的關鍵因素。本研究將轉化后的胚性愈傷組織分別在含有300、350和400 mg·L-1草甘膦的選擇培養基上進行抗性篩選,進一步分化、成苗,共獲得67株再生植株,進行CP4基因的PCR檢測,其中陽性植株43株,再生植株陽性率為64.18%,達到很好的選擇效果。
3.2 秈稻遺傳轉化體系的優化
3.2.1 受體材料的選擇及胚性愈傷組織的代齡 能否成功進行遺傳轉化的重要前提是選擇適合的植物材料作為轉化受體。水稻幼胚分裂能力強,易形成大量優質胚性愈傷組織,但受季節的影響,水稻幼胚利用不便,且在組織培養過程中易受微生物污染,轉化效率不高,因此作為轉化體存在一定的困難[16]。成熟胚方便儲存與利用,不受季節限制和胚性愈傷組織誘導率較高,通常被作為遺傳轉化和再生的良好的外源體材料。蘇軍[17]比較了不同代齡的胚性愈傷組織,發現第4、5代的胚性愈傷組織轉化效率較高,并且分化能力也較強。早代愈傷組織不易接受外源遺傳物質。但晚代愈傷組織容易出現質地軟、水漬化等現象,影響遺傳轉化成功率。本研究選擇胚性愈傷組織代齡為3~4代,可有效減少愈傷組織老化、色澤暗黃、結構松散和褐化率高等問題,有效提高了遺傳轉化效率。
3.2.2 轉化過程的優化 為提高遺傳轉化效率,本研究采取一系列措施對轉化過程進行合理優化。①轉化階段選用色澤鮮黃、外觀形態良好、結構緊致的愈傷組織與農桿菌共培養,淘汰外觀發白發軟發褐的愈傷組織。②在共培養基R2和懸浮培養基R3中加入100 μmol·L-1乙酰丁香酮,可誘導農桿菌Vir基因的活化,從而促進外源基因的整合,極大提高轉化效率[18]。③農桿菌菌液經懸浮培養液R3稀釋后,D(600)為0.5~0.8。此時為最適菌液濃度,既不會使農桿菌在轉化體表面過多繁殖影響其正常生長,又具一定的侵染能力,提高了轉化效率。④黑暗條件下共培養2~3 d為適合的共培養時長。共培養時間過短,目的基因不能成功整合至轉化體細胞內[19-20]。
