高羊茅逆境脅迫蛋白基因FaUSP 的克隆、表達及生物學功能分析
2021-04-26 01:22:56陳瑩陳錫王茜王小利
生物技術通報 2021年2期
關鍵詞:植物
陳瑩 陳錫 王茜 王小利
(貴州省農業科學院 貴州省草業研究所,貴陽 550006)
高羊茅(Festuca arundinacea)是禾本科多年生地被植物,可作為觀賞性草坪草,也可作為牧草飼養牲畜,作為我國主要草品種之一,在草坪建植與水土保持方面發揮著巨大的作用。干旱、鹽、高溫或低溫、重金屬和氧化脅迫等非生物脅迫嚴重影響了它的生長與發育,期間涉及多種復雜機制,通過激活應激反應基因將外部信號轉化為下游效應因子,稱之為受體對信號的感知。效應蛋白介導植物一些形態和生理的改變,幫助植物更好地適應脅迫條件。在多種生物中被發現。在擬南芥中,已經研究發現許多應激反應基因家族,逆境脅迫蛋白(Universal stress protein,USP)家族是其中之一。
逆境脅迫蛋白USP 又稱應激蛋白,普遍存在于細菌和植物的細胞質中,通常由111-167 個氨基酸殘基組成。逆境脅迫蛋白基因UspA 超家族最初是在大腸桿菌中發現的[1],在多重應激條件和營養缺乏等逆境條件下,其表達急劇增加,在非生物應激反應中發揮積極作用。USP 的結構域屬于疏水蛋白,在USP 肽鏈上有8 個磷酸化位點,具有應激性,并作用于蘇氨酸與絲氨酸的蛋白磷酸化。一些USP 蛋白具有使DNA 結合、修復和重新折疊的功能,可以支持生物體保護其核酸免受外部壓力,有的USP 蛋白可以與ATP 結合,可能會參與激素的合成、產物代謝、細胞內物質運輸等[2]。植物USP 蛋白中發現的催化修飾物包括絲氨酸/蘇氨酸激酶、酪氨酸激酶、U-box、SWI2/snf2、Mudr(SWIM)-鋅指結構、HDzip、陽離子交換器與IKI3。植物USP 的催化修飾物可能是在不同脅迫下選擇性的演化過程中產生,導致了不同催化修飾物與USP 域的融合,這個過程為植物提供了多種調控方式來保護它們免受外來脅迫的傷害。前人已在很多高等植物中發現USP基因,David 等[3]在擬南芥中發現了44 個USP 相似的蛋白,系統發育樹分析將其分為2 種類型,一種包含ATP 結合位點;另一種不包含ATP 結合位點。MsUspA 可能參與調控激素和次生代謝物合成以提高蘋果的抗旱性[4]。在亞洲棉中,Maqbool 等[5]成功克隆出USP1與USP2發現,其在干旱脅迫下表達量更高,推測USP基因能在干旱脅迫下對亞洲棉起到保護作用。Jung 等[6]研究表明AtUsp啟動子能被非生物誘導性脅迫與植物激素高度誘導,并產生多種脅迫的抗逆性。過表達AtUsp植株表現出較強的耐熱和抗氧化性[7]。之后陸續在大豆[8]、黃檗[9]、擬南芥[10]、紫云英[11]中發現USP基因,該基因編碼的蛋白幾乎參與所有的逆境脅迫,因此吸引了大量國內外學者對其功能進行深度研究。高羊茅中有關USP基因的抗旱性研究仍鮮見報道。
本研究通過cDNA 末端快速擴增技術獲得高羊茅逆境脅迫蛋白基因FaUSP全長,利用生物信息學技術,分析FaUSP的結構與其他物種USP 的同源性。利用RT-PCR 技術研究FaUSP在不同非生物脅迫條件下的表達情況。將FaUSP在菊苣中過量表達,研究在干旱脅迫下過量表達FaUSP對菊苣生理特性的響應,探索USP基因家族的抗逆功能和機理,旨為制備具有抗旱性和高產性的優良品種奠定理論基礎。
1 材料與方法
1.1 材料
采用的高羊茅黔草1 號是2005 年由貴州省草業研究所育成的國家牧草新品種(登記號:299)。
1.2 方法
1.2.1 材料的處理 選取飽滿的高羊茅種子,播種于花盆中,置于溫度為(23±2)℃、光照12 h/黑暗12 h、濕度是70%的光照培養箱中,定期澆水。待生長一段后進行高溫、干旱、高鹽、低氮脅迫處理。
高溫脅迫:將發芽的植株移栽到土壤基質中生長30 d,然后將高羊茅植株轉入42℃持續處理24 h,對照組的生長條件為23℃。
干旱處理:將生長14 d 的高羊茅幼苗放于30%的PEG 溶液中處理24 h,對照組為正常Hoagland 營養液。
高鹽處理:將生長14 d 的高羊茅幼苗分別放置于含有400 mmol/L NaCl 的Hoagland 營養液和正常Hoagland 營養液中處理24 h。
低氮處理:將生長30 d 幼苗分別轉移到Hoagland 營養液與無氮水培液中(營養液中NO3-被Cl-取代)進行生長處理15 d。
按照取樣時間分別取0、0.5、1、2、6、12 和24 h 共7 個點,采集處理組與對照組高羊茅的葉片用液氮速凍,-80℃保存待用。
1.2.2 RNA 提 取 采 用TaKaRa RNAiso Reagent 試劑盒進行高羊茅葉片的RNA 提取,參照RevertAid First Strand cDNA Synthesis Kit 試劑盒說明反轉錄mRNA,以總mRNA 為模板合成第一鏈cDNA[12]。
1.2.3FaUSP的克隆 參考高羊茅轉錄組測序結果中USP 相關基因片段,采用Primer Premier 5.0 設計PCR 擴增特異引物(表1)。采用5'RACE 試劑盒(Invitrogen)與3'RACE 試劑盒(Clontech)擴增獲得FaUSP的5'端與3'端,PCR 產物回收純化后送至上海生工公司測序。
1.2.4 實時熒光定量PCR 檢測 以反轉錄成的高羊茅cDNA 為模板,采用qRT-PCR 技術檢測高溫、低氮、干旱與高鹽逆境脅迫下高羊茅葉片中FaUSP的表達量,引物見表1。反應體系為6 μL ddH2O、2 μL cDNA、1 μL 上游引物、1 μL 下游引物和10 μL 2×SYBR Premix Ex Taq,置于熒光定量PCR 儀上進行反應,95℃ 3 min;94℃ 10 s,60℃ 10 s,72℃ 30 s,45 個循環,內參基因為UBI(表1)。每個取樣點3個生物學重復,3 個技術重復。

表1 高羊茅FaUSP 克隆及熒光定量PCR 的引物序列
1.2.5 生物信息學分析 使用NCBI 網站搜索進行Blast 比對,并分析編碼蛋白功能域(https://www.ncbi.nlm.nih.gov),使用SOPMA 網站預測FaUSP 蛋白的基本性質、二級結(https://npsa-prabi.ibcp.fr/cgi-bin/ npsaautomat.pl?page=npsa_sopma.html)。 使用MEGA6.0 軟件進行氨基酸序列同源性分析,構建FaUSP 蛋白序列的系統進化樹;在ExPASY 網站分析FaUSP 蛋白的相對分子量、等電點及疏水性(http://www.expasy.ch/tools/proscale.html)。
1.2.6 遺傳轉化載體的構建與菊苣的遺傳轉化 參考李小冬等[13]方法構建過量表達載體及菊苣的遺傳轉化,USP基因為逆境脅迫基因,能被多種逆境脅迫誘導,為了進一步研究USP基因的功能,將準備好的菊苣外植體與含有目標質粒的農桿菌放入無菌培養皿中浸染,使農桿菌與外植體充分接觸,將侵染好的外植體轉入培養皿中,放入人工氣候室培養,溫度為24℃,用加100 mg/mL 頭孢霉素的無菌水清洗外植體3 次,直至長出愈傷組織,每星期繼代一次,待長出根系移植到培養基質中(泥炭土∶蛭石=1∶1),最后提取DNA 進行陽性檢測。
1.2.7 干旱脅迫下轉基因菊苣生理特性的響應 提取菊苣RNA,采用熒光定量PCR 檢測野生型和轉基因植株中FaUSP的表達,引物參照表1。將野生型與轉基因的菊苣進行干旱脅迫處理,在脅迫后的0、5 和10 d 時取樣進行生理指標測定。
2 結果
2.1 高羊茅FaUSP的克隆與核酸序列分析
以高羊茅cDNA 為模板,使用特異引物FU-fwd1與FU-rev1,擴增獲得一條長度為470 bp 條帶(圖1),回收、測序后獲得高羊茅FaUSP的核心片段。利用合成的3' RACE 與5' RACE 引物(表1)進行PCR擴增,獲得3'端364 bp與5'端的315 bp的cDNA片段。利用DNAMAN 軟件將以上3 段序列進行拼接,得到總長度為844 bp 的基因全長cDNA 序列(圖2),含501 bp 的開放閱讀框,共編碼166 個氨基酸,該基因包含70 bp 的5'端非編碼區和273 bp 3'端非編碼區,命名為FaUSP。
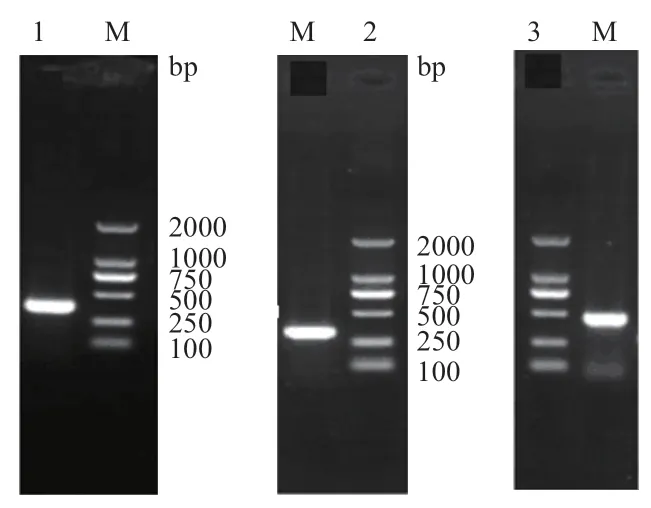
圖1 FaUSP 基因的RACE 擴增

圖2 FaUSP 核苷酸及推導的氨基酸序列
保守結構域分析表明,該基因編碼的蛋白屬于USP 家族。經系統進化樹分析,FaUSP 蛋白與禾本科植物小麥、大麥、二穗短柄草、山羊草的USP 蛋白親緣關系較近,同源性均在70%以上(圖3)。
2.2 高羊茅FaUSP編碼蛋白的生物信息學分析
利用SOPMA 軟件預測FaUSP 蛋白的二級結構,結果顯示,FaUSP 蛋白結構域主要由α-螺旋(39.76%)、延伸鏈(19.88%)、β-轉角(4.82%)和無規則卷曲(35.54%)組成。理化性質分析表明,FaUSP編碼的蛋白質的相對分子質量為18.06 kD,理論等電點為5.93,不穩定系數為32.49,疏水性分析發現,FaUSP 蛋白疏水性最大值為2.067,最小值為-3.2,為親水蛋白。
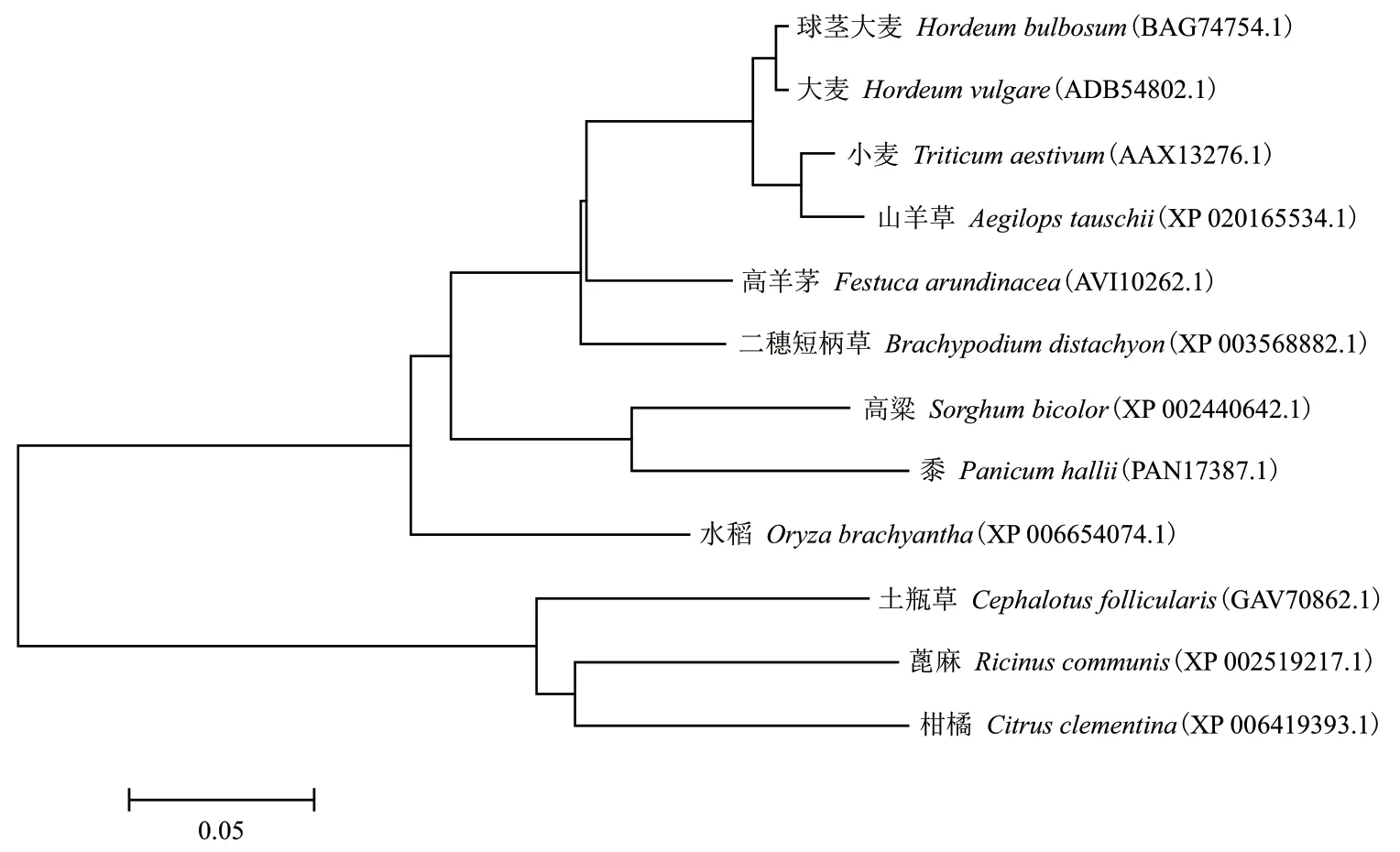
圖3 植物USP 同源基因氨基酸序列的系統進化樹分析
2.3 在非生物脅迫下高羊茅逆境脅迫FaUSP的表達模式分析
采用實時熒光定量PCR 檢測高羊茅葉片中逆境脅迫FaUSP在高鹽、低氮、干旱與高溫脅迫處理下的時空特異性的表達規律(圖4),在高溫脅迫下,FaUSP的表達在脅迫處理0.5 h 后被誘導上升,處理6 h 達到峰值,在脅迫處理12 h 后受到抑制,然而與對照相比,在脅迫12-24 h 時FaUSP表達顯著降低;在干旱脅迫下,脅迫處理早期(0-1 h)FaUSP的表達與正常條件相當,呈現下降趨勢,在脅迫6 h時,FaUSP的表達受到極顯著的誘導。在低氮脅迫下,與正常條件下相比,FaUSP的表達在脅迫處理0.5-2 h 時顯著降低,在脅迫6-24 h 后FaUSP表達被誘導顯著上升。在高鹽脅迫0-1 h 時FaUSP表達與對照相當,在脅迫12-24 h,FaUSP表達顯著被誘導上升,在脅迫24 h 時表達量達到峰值,為對照組的3 倍,說明高羊茅FaUSP受干旱、高溫、氮脅迫及鹽脅迫的誘導,該基因可能與抗性相關。
2.4 FaUSP轉基因菊苣的表型與表達分析
野生型菊苣(WT)與轉基因菊苣(FaUSP超量表達)在相同的培養條件下呈現不同的表型(圖5),培養62 d 后,野生型菊苣處于抽穗期,而超表達FaUSP的菊苣呈現晚花表型。以野生型為對照,檢測5 株轉基因菊苣FaUSP表達量(圖6),與野生型相比,FaUSP的表達量顯著上調,升高了7-11 倍。
2.5 干旱脅迫對FaUSP轉基因菊苣生理特性的影響
轉基因與野生型的菊苣隨著干旱脅迫時間的延長,葉綠素含量呈現先升后降的趨勢(圖7),干旱脅迫下轉基因組和對照組的葉綠素含量相比差異不顯著。而轉基因組菊苣的可溶性蛋白含量隨干旱脅迫時間的延長呈上升的趨勢,在脅迫的第10 天達到峰值,顯著高于對照組。說明干旱脅迫能有效誘導菊苣的可溶性蛋白的合成,使其含量上升。
谷胱甘肽還原酶(Glutathione reductase,GR)作為植物體內重要的抗氧化劑與自由基清除劑,保護機體不受逆境脅迫的傷害(圖7),隨著干旱脅迫時間的延長,轉基因與野生型的菊苣的GR 含量均呈現先升后降的趨勢,轉基因組在干旱脅迫下的GR含量顯著高于對照組,說明干旱脅迫下轉基因組菊苣體內GR 的合成能力顯著強于對照組。
抗氧化酶系統是植物遭受逆境脅迫重要的防御體系,超氧化物岐化酶(SOD)、過氧化物酶(POD)與過氧化氫酶(CAT)是抗氧化酶系的主要組成部分。隨著干旱脅迫時間的延長,轉基因組與對照組的CAT 與POD 活性呈現先升后降的趨勢,均在脅迫第5 天活性最高,轉基因組CAT 與POD 活性顯著高于對照組。轉基因組SOD 的活性隨著干旱脅迫時間的延長呈現升高趨勢,在脅迫10 d 達到了峰值,說明干旱脅迫下轉基因組能夠使菊苣葉片抗氧化酶活性顯著升高。

圖4 非生物脅迫下FaUSP 基因在高羊茅中的轉錄水平
3 討論
逆境脅迫蛋白USP 是一個保守的蛋白家族,參與大量逆境脅迫應答反應。最早于大腸桿菌中被發現[1]。在高等植物中,USP 的發現起始于水稻。USP 蛋白一般存在于細胞質中,是普遍存在于植物中的抗性相關蛋白,在植物受到外界脅迫時會受誘導表達上調,增強植物的抗逆性[14-15]。趙莘等[16]發現抗病的中國野生華東葡萄株系在接種白粉病后VpUS表達量呈明顯的增加趨勢,表明VpUSP在與白粉病菌互作過程中具有表達活性等。本研究利用cDNA 末端快速擴增技術獲得高羊茅逆境脅迫蛋白FaUSP,序列全長844 bp,含501 bp 的開放閱讀框,共編碼166 個氨基酸。保守結構域分析表明,該基因編碼的蛋白屬于USP 家族。對其氨基酸序列進行分析,結果表明,高羊茅FaUSP 蛋白與禾本科植物小麥、大麥、二穗短柄草的USP 蛋白親緣關系較近,在進化樹分析中高羊茅FaUSP 與禾本科植物的USP聚在一起。這可能是同為禾本科的植物,與其他同科植物的家族成員具有共同的保守區,而逆境脅迫蛋白FaUSP 在其他科屬中存在著分化。

圖5 轉基因菊苣的表型

圖6 轉基因菊苣FaUSP 的表達分析

圖7 轉基因菊苣在干旱脅迫下的生理應答差異
Usp基因在植物中普遍存在,其表達受多種逆境如干旱、高溫、澇害等條件調控,被認為與植物脅迫反應有關。研究發現USP 基因表達受多種逆境條件誘導,如Sauter 等[17]發現水稻OsUSP1受乙烯與澇害的誘導表達;小麥脅迫相關基因W1受干旱、低溫的誘導[18];黃姍等[8]發現大豆在鹽、ABA、干旱脅迫下大豆Usp1被誘導表達量上調。本研究進行實時熒光定量PCR 表達分析顯示,高羊茅FaUSP受干旱、高溫、氮脅迫及鹽脅迫的誘導表達上調,表明該基因與抗性相關。關于高羊茅FaUSP的分子作用機制,進一步構建過表達載體遺傳轉化轉入菊苣進行功能驗證研究。
在對植物構成威脅的環境因素中,干旱是最常見的非生物脅迫之一,它對植物生長發育的影響比其他環境因素更為嚴重[19-20]。為了減少干旱的危害,植物已經進化出了幾種方式應對,包括提高植物自身耐受性、抵御干旱的傷害與適應干旱的能力,這些適應性可以通過調控一系列不同的信號轉導基因加以實現[21-23]。逆境脅迫蛋白基因USP 在干旱脅迫下被誘導并發揮積極作用,提高了植物的抗逆能力。Loukehaich 等[24]發現過量表達的SpUsp明顯提高了番茄抗旱性。生理生化指標的測定結果表明轉基因SpUsp煙草可以保護細胞免受氧化損傷[25]。這與本研究結果一致,干旱脅迫下過量表達FaUSP菊苣可溶性蛋白、GR、SOD、POD、CAT 的合成顯著高于對照組,說明FaUSP能夠提高植物的抗旱能力。
4 結論
從高羊茅中克隆得到逆境脅迫蛋白FaUSP,含501 bp 的開放閱讀框,共編碼166 個氨基酸,該基因編碼的蛋白屬于USP 家族。其受干旱、高溫、氮脅迫及鹽脅迫的誘導。干旱脅迫下菊苣中過表達FaUSP可以顯著提高抗旱能力。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13