E3泛素連接酶1對SOX2蛋白穩(wěn)定性的影響
2021-04-28 14:58:06孫瀟智劉玉杰李資益管恒毓
檢驗醫(yī)學 2021年4期
關(guān)鍵詞:水平
郭 平, 孫瀟智, 劉玉杰, 李資益, 楊 婧, 管恒毓, 廖 兵
(上海交通大學基礎(chǔ)醫(yī)學院組織胚胎學與遺傳發(fā)育學系,上海 200025)
人胚胎干細胞(human embryonic stem cell,hESC)是將著床前的人囊胚的內(nèi)細胞團細胞在體外培養(yǎng)建立的細胞系[1]。由于hESC擁有無限自我更新的特點和分化為所有成體細胞類型的發(fā)育潛能,能為再生醫(yī)學的研究和應用提供理想的細胞模型與無限的種子細胞來源,所以多年來一直是干細胞領(lǐng)域的研究熱點。基于hESC廣闊的應用前景,將其用于臨床治療前必須明確其調(diào)控自我更新和多能性維持及分化的分子網(wǎng)絡(luò)。hESC的特征維系涉及轉(zhuǎn)錄因子、信號通路和表觀遺傳修飾等的共同調(diào)節(jié)[2-3]。其中核心轉(zhuǎn)錄因子——性別決定區(qū)Y-box蛋白2(sex-determining region Y-box protein 2,SOX2)在hESC細胞分化決定過程中發(fā)揮著關(guān)鍵作用[4]。SOX2的變化將影響八聚體結(jié)合轉(zhuǎn)錄因子4(octamer-binding transcription factor 4,OCT4)、NANOG等其他轉(zhuǎn)錄因子的水平,進而影響維持hESC自我更新和多能性相關(guān)基因的表達與抑制[5]。目前,SOX2蛋白水平的調(diào)控機制尚未被完全闡明。因此,深入研究SOX2表達的調(diào)控機制無論對hESC的基礎(chǔ)研究還是臨床應用都具有重要意義。
蛋白質(zhì)翻譯后修飾在調(diào)控蛋白質(zhì)的結(jié)構(gòu)與功能方面發(fā)揮著重要作用,通過改變蛋白質(zhì)的功能特性或空間結(jié)構(gòu)影響各項生物學過程。蛋白質(zhì)翻譯后修飾的方式多達幾十種,其中泛素化修飾是細胞內(nèi)廣泛且重要的修飾方式之一。蛋白質(zhì)的泛素化修飾是由E1泛素活化酶、E2泛素結(jié)合酶和E3泛素連接酶(WW domain containing E3 ubiquitin protein ligase,WWP)催化的酶促級聯(lián)反應過程,通過建立泛素蛋白與底物蛋白的共價鍵連接,影響底物蛋白的亞細胞定位、活性和穩(wěn)定性等[6]。有研究結(jié)果顯示,在hESC中,WWP2能催化OCT4蛋白的泛素化修飾,并介導其經(jīng)26S蛋白酶體途徑的降解,調(diào)控OCT4蛋白處于合適的水平,從而維持hESC的自我更新[7]。為進一步揭示(2-WW-HECT家族WWP1和WWP2對核心轉(zhuǎn)錄因子OCT4、SOX2、NANOG蛋白水平的可能作用,本研究擬通過HEK 293FT細胞的共表達實驗,探討WWP1對SOX2蛋白的調(diào)控作用。
1 材料和方法
1.1 主要材料
1.1.1 質(zhì)粒 OCT4過表達質(zhì)粒(pCMVOCT4)、HA-SOX2過表達質(zhì)粒(pCDNA-HASOX2)、空白質(zhì)粒(pPyCAG-flag)、Flag-WWP1過表達質(zhì)粒(pPyCAG-flag-WWP1)、Flag-WWP2過表達質(zhì)粒(pPyCAG-flag-WWP2)、WWP1融合蛋白表達質(zhì)粒(pGEX-4T-1-WWP1)、WWP1-C890A突變質(zhì)粒(pGEX-4T-1-WWP1-C890A)、SOX2融合蛋白表達質(zhì)粒[pET-30a(+)-SOX2]均由本實驗室自行構(gòu)建。NANOG過表達質(zhì)粒(pSin-EF2-NANOG-Puro)購自美國Addgene公司。
1.1.3 抗體 SOX2抗體(批號AF2018,美國R&D公司)、OCT4抗體(批號sc-2079,美國Santa Cruiz公司)、NANOG抗體(批號3369-1,美國EPITOMICS公司)、α-微管蛋白(α-TUBULIN)抗體(批號T5168,美國Sigma公司)、Flag標簽(Flag-tag)抗體(批號F3165,美國Sigma公司)、多組氨酸標簽(Histag)抗體(批號GT359,美國Sigma公司)、泛素(Ubiquitin)抗體(批號3936,美國Cell Signaling公司)、辣根過氧化物酶偶聯(lián)二抗(羊抗人IgG抗體、鼠抗人IgG抗體、兔抗人IgG抗體,美國Cell Signaling公司)。
1.1.4 主要試劑 非干擾型蛋白濃度測定試劑盒、甘氨酸、三羥甲基氨基甲烷[Tris(hydroxymethyl)methyl aminomethane,Tris]、十二烷基硫酸鈉(sodium dodecyl sulfate,SDS)、氯化鈉、吐溫-20、甲醇、5'-三磷酸腺苷(adenosine 5'-triphosphate,ATP)、乙二胺四乙酸(ethylenediaminetetraacetic acid,EDTA)、甘油、NP-40溶液均購自生工生物工程(上海)股份有限公司。1×磷酸鹽緩沖液、0.25%胰酶、Opti-MEM減血清培養(yǎng)基、胎牛血清、100×非必需氨基酸、100×丙酮酸鈉均購自美國Gibco公司。E1泛素活化酶、E2泛素結(jié)合酶、His-Tagged-Ub均購自德國Merck公司。聚乙烯亞胺(poly ethylenimine,PEI)、DMEM高糖培養(yǎng)基均購自美國Sigma公司。Glutathione Sepharose 4B購自美國Amersham公司,Ni-NTA His-Bind resin購自德國Novagen公司。
1.1.5 免疫印跡法試劑配制 10×電泳緩沖液(甘氨酸144 g,Tris 30.3 g,SDS 10 g,加水定容至1 L),10×TBS-T[Tris 1 mol/L(pH值7.6)200 mL,氯化鈉80 g,吐溫-20 1%,加水定容至1 L];1×轉(zhuǎn)膜緩沖液(甲醇 200 mL,甘氨酸14.4 g,Tris 3.1 g,加水定容至1 L)。
1.2 方法
1.2.1 細胞培養(yǎng) 將HEK 293FT細胞在37 ℃、5% CO2、100%濕度條件下培養(yǎng),密度>80%后以0.25%胰酶消化傳代。
1.2.2 質(zhì)粒轉(zhuǎn)染 使用PEI進行質(zhì)粒轉(zhuǎn)染,轉(zhuǎn)染前1天用0.25%胰酶消化細胞重新鋪板。計算質(zhì)粒用量和PEI工作液(1 μg/μL)用量,確保PEI工作液體積與待轉(zhuǎn)質(zhì)粒質(zhì)量的比例為1∶3,將其加入Opti-MEM減血清培養(yǎng)基中,室溫靜置30 min后將混合液加入對應細胞中。轉(zhuǎn)染12 h后更換新培養(yǎng)液,換液后繼續(xù)培養(yǎng)36 h收取細胞。
1.2.3 免疫印跡法 細胞總蛋白提取、蛋白濃度測定及免疫印跡法操作步驟參照文獻[7]。
1.2.4 谷胱甘肽S轉(zhuǎn)移酶(glutathione S-transferase,GST)融合蛋白表達與純化將含目的基因的pGEX-4T-1質(zhì)粒轉(zhuǎn)化感受態(tài)菌BL-21(DE3),挑單克隆培養(yǎng),添加異丙基硫代半乳糖苷至終濃度0.2 mmol/L,37 ℃培養(yǎng)2 h,誘導融合蛋白表達。冰浴超聲波裂菌直至溶液澄清,加入Glutathione Sepharose 4B結(jié)合GST融合蛋白(4 ℃,轉(zhuǎn)動孵育60 min)。洗滌3次后加200 μL洗脫緩沖液(4 ℃,轉(zhuǎn)動洗脫30 min),離心取上清。將誘導前菌液、誘導后菌液、裂菌后離心所得沉淀、洗脫樣本進行SDS-PAGE電泳,考馬斯亮藍染色檢測融合蛋白的制備情況。
目前,可完全降解的塑料主要以淀粉、纖維素、糖和有機酸的一種或幾種為基本原料,經(jīng)過物理、化學或生物學方法加工而成的。受原材料成本、膜材料價格以及推廣應用的影響,農(nóng)用地膜仍然較多采用纖維素或淀粉為基本原料,包括淀粉基地膜和紙地膜等。其中,紙地膜以造紙工藝為基礎(chǔ),以植物纖維為基本原料,在植物紙漿的基礎(chǔ)上,通過添加濕強劑、防腐劑和透明劑等化學助劑,采用常規(guī)造紙工藝抄制出原紙,然后對其進行加工處理,使紙張具有農(nóng)用地膜所要求的機械強度和透光、透水、保溫、增溫、保墑性或其他增肥和除草功能[2-4]。
1.2.5 His融合蛋白表達與純化 將含目的基因的pET-30a(+)質(zhì)粒轉(zhuǎn)化感受態(tài)菌BL-21(DE3)37 ℃培養(yǎng)過夜。挑單克隆接培養(yǎng),添加異丙基硫代半乳糖苷至終濃度為1 mmol/L,37 ℃培養(yǎng)4 h誘導His融合蛋白表達。冰浴超聲波裂菌直至溶液澄清,加入His-Bind resin結(jié)合His融合蛋白(4 ℃,轉(zhuǎn)動孵育60 min)。洗滌3次后加洗脫緩沖液(4 ℃,轉(zhuǎn)動洗脫30 min),將His融合蛋白從beads洗脫。洗脫產(chǎn)物分裝后于-80 ℃分批次凍存待用。洗脫樣本采用SDSPAGE進行電泳與考馬斯亮藍染色檢測融合蛋白的純化情況。
1.2.6 GST pull-down實驗、免疫共沉淀(coimmunoprecipitation,Co-IP)實驗、體外蛋白質(zhì)泛素化修飾 參照文獻[8]的實驗步驟進行GST pull-down實驗、Co-IP實驗、體外蛋白質(zhì)泛素化修飾。
1.2.7 HEK 293FT細胞質(zhì)粒共表達實驗 通過質(zhì)粒轉(zhuǎn)染在HEK 293FT細胞中分別共表達OCT4、SOX2、NANOG和WWP1或WWP2。WWP1或WWP2的過表達質(zhì)粒用量分別為0、0.2、0.4、0.8 μg,OCT4、SOX2、NANOG過表達質(zhì)粒的用量均為0.1 μg。通過免疫印跡法檢測WWP1或WWP2過表達對OCT4、SOX2、NANOG蛋白水平的影響。根據(jù)蛋白條帶信號強度計算各蛋白的相對表達量。
1.2.8 SOX2蛋白合成抑制實驗 在HEK 293FT細胞中單獨過表達SOX2或與WWP1組成共表達體系,使用放線菌酮(cyclohexmide,CHX)處理細胞,抑制蛋白質(zhì)合成。通過免疫印跡法檢測SOX2在CHX處理8、16、24 h和未處理細胞中的蛋白水平。根據(jù)蛋白條帶信號強度計算各蛋白相對表達量并繪制SOX2蛋白的半衰期曲線。
1.2.9 SOX2蛋白降解抑制實驗 在HEK 293FT細胞中共表達SOX2與WWP1,轉(zhuǎn)染36 h后分別向細胞培養(yǎng)液中加入不同濃度蛋白酶體抑制劑MG132(25、50 μmol/L)或溶酶體抑制劑氯喹(100、200 μmol/L),對照組不加MG132或氯喹,4 h后收取細胞,采用免疫印跡法檢測。
1.3 統(tǒng)計學方法
采用SPSS 20.0軟件進行統(tǒng)計分析。所有實驗均重復3次。呈正態(tài)分布的數(shù)據(jù)以±s表示,多組間比較采用方差分析,2個組之間比較采用非配對Student'st檢驗。以P<0.05為差異有統(tǒng)計學意義。
2 結(jié)果
2.1 WWP1或WWP2對OCT4、SOX2、NANOG蛋白水平的影響
將WWP1或WWP2分別與OCT4、SOX2、NANOG組成共表達體系,并采用免疫印跡法檢測各蛋白的相對表達量。結(jié)果顯示,在WWP1與OCT4、WWP1與SOX2及WWP2與OCT4共表達體系中WWP1或WWP2均呈劑量依賴性地下調(diào)對應的共表達蛋白OCT4或SOX2水平,且以WWP1與SOX2共表達體系中SOX2蛋白的表達下降最為明顯(相對表達量為0.1)。在其他共表達體系中,WWP1和WWP2對對應的共表達蛋白均無明顯的下調(diào)作用。見圖1。

圖1 WWP1和WWP2對OCT4、SOX2、NANOG蛋白相對表達量的影響
2.2 HEK 293FT細胞中過表達WWP1對SOX2蛋白穩(wěn)定性的影響
以單獨過表達SOX2為對照體系,在未處理細胞樣本(0 h)中,WWP1與SOX2共表達體系中的SOX2蛋白水平低于對照體系(P<0.05);使用CHX處理8、16、24 h后,WWP1與SOX2共表達體系中的SOX2蛋白水平與對照體系之間差異無統(tǒng)計學意義(P>0.05)。SOX2蛋白半衰期曲線分析結(jié)果顯示,隨著CHX處理時間的延長,SOX2蛋白水平逐漸下降。與對照體系比較,WWP1與SOX2共表達體系中的SOX2蛋白水平在CHX處理24 h后下降速度加快(P<0.05)。見圖2。

圖2 CHX處理不同時間對照體系及WPP1與SOX2共表達體系中SOX2蛋白的變化
2.3 MG132和氯喹對SOX2蛋白降解的抑制作用添加25或50 μmol/L MG132后,WWP1與
SOX2共表達體系中SOX2蛋白水平隨WWP1水平的升高而逐漸降低。添加100或200 μmol/L氯喹后,隨著WWP1水平的升高,WWP1與SOX2共表達體系中的SOX2蛋白水平未出現(xiàn)明顯變化。見圖3。
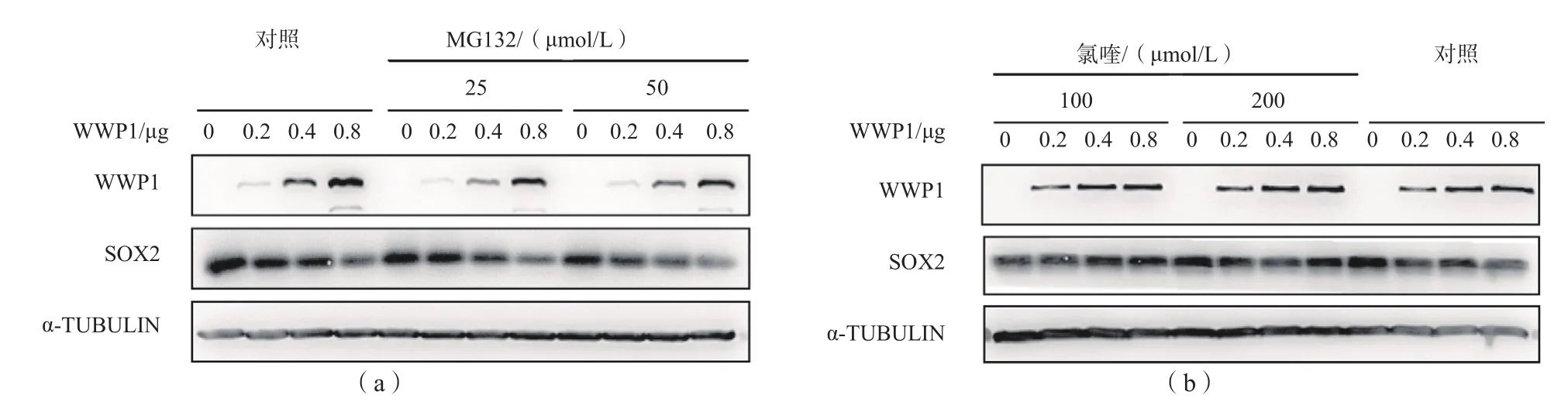
圖3 不同濃度MG132和氯喹對SOX2蛋白降解的抑制作用
2.4 重組融合蛋白的表達與純化結(jié)果
結(jié)果顯示,在相對分子質(zhì)量150 000附近出現(xiàn)GST-WWP1和GST-WWP1-C890A重組融合蛋白的特異性條帶。在相對分子質(zhì)量45 000附近出現(xiàn)His-SOX2重組融合蛋白特異性條帶。見圖4。

圖4 GST-WWP1、GST-WWP1-C890A 和His-SOX2重組融合蛋白考馬斯亮藍染色結(jié)果
2.5 GST pull-down實驗檢測WWP1與SOX2的直接相互作用
GST pull-down實驗結(jié)果顯示,Glutathione sepharose 4B可沉淀GST-WWP1(免疫印跡法陽性),且GST-WWP1與His-SOX2結(jié)合后陽性條帶更為明顯,而Glutathione sepharose 4B與His-SOX2(對照)僅有微弱的非特異性結(jié)合條帶。見圖5。表明GST-WWP1與His-SOX2在體外條件下能夠發(fā)生直接的相互作用。

圖5 GST pull-down實驗和免疫印跡法檢測WWP1與SOX2相互作用的結(jié)果
2.6 Co-IP實驗檢測外源性WWP1與SOX2在HEK 293FT細胞中的相互作用
免疫印跡法結(jié)果顯示,全細胞裂解液(whole cell lysis,WCL)樣本中可檢測到Flag-WWP1和HA-SOX2的過表達,在Flag抗體-免疫共沉淀蛋白復合物中可同時檢測到Flag-WWP1與HA-SOX2蛋白。在單獨過表達HA-SOX2的細胞CO-IP實驗樣本中,SOX2蛋白的表達量較低;與單獨過表達HA-SOX2的對照組相比較,在Flag-WWP1與HA-SOX2共表達體系中,免疫共沉淀蛋白復合物的HA-SOX2蛋白表達量較高。見圖6。
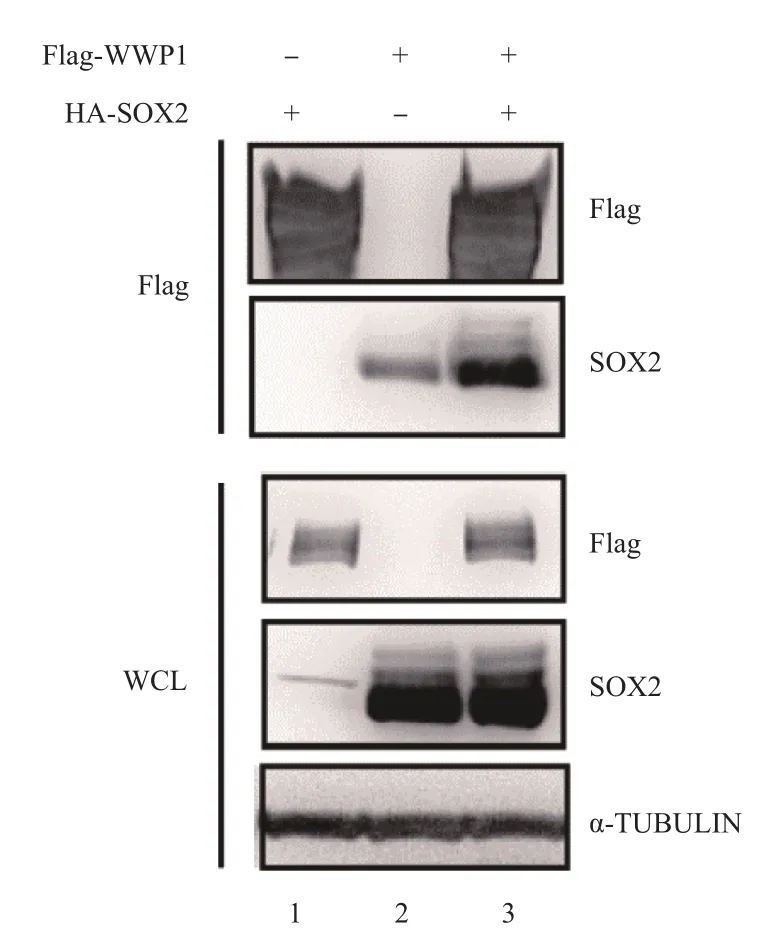
圖6 Co-IP實驗和免疫印跡法檢測WWP1與SOX2的相互作用
2.7 體外模擬WWP1催化蛋白質(zhì)泛素化修飾結(jié)果
免疫印跡法結(jié)果顯示,當使用泛素抗體進行檢測時會出現(xiàn)高相對分子質(zhì)量的條帶(圖7上半部分第3~5泳道),在不加His-Ub的情況下這些條帶消失(圖7上半部分第1泳道),說明蛋白泛素化修飾反應成功。
在不加入GST-WWP1的情況下,泛素化蛋白條帶消失(圖7上半部分第2泳道)。當突變WWP1蛋白關(guān)鍵酶活位點[第890位半胱氨酸突變?yōu)楸彼幔–890A)]時,泛素化修飾條帶消失(圖7上半部分第6泳道)。在不加入His-SOX2的情況下,泛素化修飾條帶仍會出現(xiàn)(圖7上半部分第3泳道)。使用SOX2抗體進行檢測,發(fā)現(xiàn)加入高劑量GST-WWP1后出現(xiàn)高相對分子質(zhì)量條帶(圖7下半部分第5泳道);當突變WWP1蛋白關(guān)鍵酶活位點后,這些條帶減弱(圖7下半部分第6泳道)。

圖7 體外泛素化修飾反應與免疫印跡法檢測WWP1催化的蛋白質(zhì)泛素化修飾
3 討論
細胞內(nèi)蛋白質(zhì)泛素化修飾通過對蛋白質(zhì)的活性、定位、穩(wěn)定性以及相互作用的調(diào)控參與包括DNA修復[9]、轉(zhuǎn)錄調(diào)控[10]、囊泡運輸[11]、細胞凋亡[12]等在內(nèi)的細胞生理活動,其最重要的功能之一是通過蛋白酶體途徑或溶酶體途徑降解蛋白質(zhì),以維持細胞生理功能及細胞穩(wěn)態(tài)。在hESC中,WWP2通過調(diào)控OCT4蛋白水平維持hESC的自我更新[8],但在未分化的小鼠胚胎干細胞(mouse embryonic stem cell,mESC)中敲低WWP2并不影響OCT4蛋白的泛素化修飾水平和總蛋白量,而當mESC處于分化狀態(tài)時WWP2又可促進OCT4泛素化修飾后經(jīng)由26S蛋白酶體的降解,從而迅速下調(diào)OCT4蛋白的水平,保障細胞分化的順利進行[7]。目前已對C2-WW-HECT家族WWP2調(diào)控核心轉(zhuǎn)錄因子OCT4翻譯后修飾的部分機制有了一定認識,但對WWP2同家族的WWP1能否影響OCT4、SOX2、NANOG蛋白的水平仍不清楚。為此,本研究在HEK 293細胞中過表達人源WWP1或WWP2以及核心轉(zhuǎn)錄因子OCT4、SOX2、NANOG,發(fā)現(xiàn)WWP1呈劑量依賴方式下調(diào)SOX2蛋白的水平,半衰期曲線顯示共表達WWP1可導致SOX2蛋白的穩(wěn)定性下降。考慮到WWP1是一種WWP,因此推測WWP1可能通過泛素化修飾下調(diào)SOX2蛋白的穩(wěn)定性和表達水平,在hESC中調(diào)控SOX2蛋白處于合適的水平,從而維持hESC的自我更新。
本研究通過體外泛素化實驗證明WWP1是SOX2的WWP。在不加GST-WWP1的反應中,泛素化反應信號條帶消失,提示W(wǎng)WP1可能是催化蛋白泛素化修飾的WWP。當突變WWP1蛋白關(guān)鍵酶活位點第890位半胱氨酸為丙氨酸(C890A)時,以上泛素化修飾信號條帶消失,進一步證明WWP1的WWP功能,同時也表明C890作為WWP1發(fā)揮泛素連接酶活性的關(guān)鍵位點催化了SOX2的泛素化修飾。本研究結(jié)果還顯示,在不加His-SOX2的反應中仍能檢測到泛素化修飾條帶,說明WWP1可能發(fā)生了自我泛素化修飾。為了區(qū)分泛素化修飾WWP1與SOX2,本研究使用SOX2抗體進行檢測,發(fā)現(xiàn)高劑量GSTWWP1組出現(xiàn)高相對分子質(zhì)量的條帶,而突變WWP1酶活位點后條帶顯著減弱,提示SOX2可能發(fā)生了多聚泛素化修飾。
當使用26S蛋白酶體抑制劑MG132或泛素溶酶體抑制劑氯喹處理過表達WWP1和SOX2的HEK 293FT細胞時,MG132并不能抑制WWP1對SOX2的降解,而氯喹可以在一定程度上抑制SOX2蛋白的降解,提示W(wǎng)WP1催化的泛素化修飾可能介導SOX2蛋白經(jīng)泛素-溶酶體途徑降解。在hESC中,細胞內(nèi)蛋白質(zhì)的降解主要通過泛素-蛋白酶體降解途徑,而溶酶體對hESC蛋白質(zhì)穩(wěn)態(tài)與細胞命運影響的研究相對較少。有研究結(jié)果顯示,在早期分化過程中,mESC和hESC均有自噬溶酶體激活的現(xiàn)象[13-14]。在誘導hESC向神經(jīng)外胚層分化時,自噬溶酶體與核因子E2相關(guān)因子2(nuclear factor E2-related factor 2,Nrf2)相互作用促進該分化事件的進行[15]。CHO等[16]證實泛素蛋白酶體途徑與泛素溶酶體途徑可以共同調(diào)節(jié)核心轉(zhuǎn)錄因子的蛋白水平。LV等[17]的研究結(jié)果顯示,表皮生長因子受體可通過抑制SOX2的溶酶體降解來提高口腔癌細胞的干細胞特性。但調(diào)控SOX2經(jīng)由泛素-溶酶體途徑降解的WWP一直未被發(fā)現(xiàn)。本研究結(jié)果顯示,WWP1可促進SOX2蛋白經(jīng)泛素-溶酶體途徑降解,為揭示泛素-溶酶體蛋白降解系統(tǒng)在hESC中的作用機制提供了新的證據(jù)。但泛素-溶酶體蛋白降解系統(tǒng)主要降解細胞質(zhì)中的蛋白質(zhì),而核蛋白SOX2是如何被該系統(tǒng)降解還有待進一步深入研究。本研究結(jié)果有助于闡明hESC中SOX2蛋白水平的調(diào)控機制,為揭示核心轉(zhuǎn)錄因子決定hESC細胞命運的調(diào)控網(wǎng)絡(luò)提供新的線索。
綜上所述,WWP1是SOX2特異的WWP,介導SOX2蛋白經(jīng)泛素-溶酶體途徑降解,進而下調(diào)SOX2蛋白的表達。
猜你喜歡
美與時代·美術(shù)學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(shè)(2019年6期)2019-10-08 08:55:48
人大建設(shè)(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(shè)(2017年6期)2017-09-26 11:50:44
學苑創(chuàng)造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
