
暗紋東方鲀MyoD基因的克隆及生物信息學分析
2021-05-11 16:08:47祁艷鳳尚鳳芹余云登周金旭王寧王秀利
安徽農業科學 2021年8期
祁艷鳳 尚鳳芹 余云登 周金旭 王寧 王秀利
摘要 提取暗紋東方鲀肌肉的總RNA,采用RT-PCR的方法來克隆暗紋東方鲀MyoD基因的cDNA序列,并通過生物信息學分析,對該基因所編碼蛋白的理化性質、親疏水性、亞細胞定位以及蛋白質結構等進行初步的分析和預測。結果顯示,該基因編碼區全長為846 bp,共編碼了281個氨基酸,編碼蛋白為親水性非跨膜類蛋白,無信號肽結構,其亞細胞定位于細胞核中。系統進化樹分析結果表明其與紅鰭東方鲀同源性最高,親緣關系最近。
關鍵詞 暗紋東方鲀;MyoD基因;基因克隆;生物信息學
中圖分類號 S 917.4文獻標識碼 A
文章編號 0517-6611(2021)08-0089-05
doi:10.3969/j.issn.0517-6611.2021.08.024
開放科學(資源服務)標識碼(OSID):
Cloning and Bioinformatics Analysis of MyoD Gene of Takifugu obscurus
QI Yan-feng,SHANG Feng-qin,YU Yun-deng et al (College of Fisheries and Life Science,Dalian Ocean University,Dalian,Liaoning 116023)
Abstract The total RNA of Takifugu obscurus muscle was extracted,and the cDNA sequence of MyoD gene of Takifugu obscurus was cloned by RT-PCR method.And through bioinformatics analysis,preliminary analysis and prediction of the physicochemical properties,hydrophilicity and hydrophobicity,subcellular localization and protein structure of the protein encoded by the gene were carried out.The results showed that the full length of the coding region of the gene was 846 bp,which encoded a total of 281 amino acids.The encoded protein was a hydrophilic non-transmembrane protein with no signal peptide structure,and its subcellular location was in the nucleus.The results of phylogenetic tree analysis showed that it had the highest homology with the redfin puffer and the closest genetic relationship.
Key words Takifugu obscurus;MyoD gene;Gene cloning;Bioinformatics
生肌調節因子MRFs家族包括MyoD、MyoG、Myf5和Myf6 這4種調節因子,該家族在成肌細胞的發育和分化方面都起著絕對作用[1-6]。MyoD基因作為生肌調節因子家族中的重要一員,主要負責調控脊椎動物胚胎期的肌肉發育,促進骨骼肌的形成與分化,并可以通過多種途徑來達到激活肌肉轉錄的目的,從而促進成肌細胞的分化;如果MyoD基因缺失,可導致成肌細胞無法進行增殖和分化,從而影響個體正常的生長和發育[7-13]。暗紋東方鲀的可食用部位主要是肌肉,而MyoD基因對肌肉生長的調控作用是至關重要的,所以對暗紋東方鲀MyoD基因的研究對改良其肉質有重要意義。
暗紋東方鲀(Takifugu obscurus)別稱橫紋多紀鲀,屬于鲀形目、鲀科、東方鲀屬。暗紋東方鲀屬于暖溫性底層洄游性魚類,生活于近海地帶,在我國主要分布于黃海、東海、長江中下游區域。春季親魚溯河生殖洄游進入淡水江河直流進行繁殖,幼魚在淡水中成長[14]。暗紋東方鲀肉質雪白且少刺,味道尤為鮮嫩可口,不僅含有大量的蛋白質和脂肪,而且還有多種人體所含的必需氨基酸以及其他營養物質,自古就享有“魚中之王”和“菜肴之冠”的美名。在一些亞洲國家如中國、日本和朝鮮都有著久遠的食用暗紋東方鲀的飲食習慣,特別是在日本市場尤為暢銷,是我國十分重要的經濟魚類。暗紋東方鲀能成功地在純淡水中養殖,有著很高的經濟價值,所以暗紋東方鲀養殖業的發展是十分迅速的,目前養殖規模也比較大,其養殖區域正從沿海向內地迅速擴展。近幾年,沿岸的河豚資源量因過度開發而表現出每年急劇減少的趨勢,為滿足市場的需求,就要擴大養殖河鲀的生產量[15-16]。目前在暗紋東方鲀的養殖技術方面研究比較廣泛,但對其基因水平的了解知之甚少,還沒有暗紋東方鲀MyoD基因組全序列的報道。該試驗使用反轉錄和RT-PCR方法,對暗紋東方鲀MyoD基因進行克隆及生物信息學分析,研究其基因序列的結構、進化關系及理化性質,以期為進一步探究魚類MyoD基因的結構與功能奠定理論基礎。
1 材料與方法
1.1 材料
試驗材料來自江蘇南通中洋集團,隨機選擇健康的3尾3月齡暗紋東方鲀幼魚作為基因克隆的試驗樣本;提取RNA的Trizol試劑、反轉錄試劑盒和Taq酶均購自TaKaRa生物工程有限公司。
1.2 方法
1.2.1 暗紋東方鲀總RNA提取與cDNA鏈合成。
剪取適量的暗紋東方鲀肌肉,加入液氮后進行充分研磨,使用Trizol法來提取暗紋東方鲀的總RNA。提取后通過凝膠電泳檢測來驗證RNA純度是否可以用于后續試驗。使用TaKaRa反轉錄試劑盒將RNA 反轉錄為cDNA,反應體系為20 μL,反應條件為37 ℃ 15 min、85 ℃ 5 s,共1個循環,4 ℃保存。反應結束后將產物保存于-20 ℃冰箱以備后續試驗使用。
1.2.2 基因引物設計與合成。
參考紅鰭東方鲀的MyoD基因序列(NM_001032771)設計暗紋東方鲀MyoD基因 CDS區PCR引物。使用Primer Premier 5.0軟件來設計克隆基因編碼區全長所需的引物。引物序列由北京六合華大基因科技有限公司合成,所合成的引物序列如表l所示。
1.2.3 基因克隆。以反轉錄合成的cDNA為模板,采用RT-PCR方法進行目的片段的擴增,PCR反應體系(25 μL)為ddH2O 14 μL、10×Buffer 2.5 μL、dNTP 2 μL,上游引物MyoD-F和下游引物MyoD-R各1 μL,模板 4 μL,Taq酶0.5 μL。PCR反應程序:94 ℃預變性5 min進入循環;94 ℃變性30 s,59 ℃ 退火30 s,72 ℃延伸30 s,共計35個循環,4 ℃保存。將PCR產物在濃度為1%的瓊脂糖凝膠中進行電泳,結果所得條帶清晰符合測序要求,送往北京六合華大基因科技有限公司測序。
1.2.4 編碼蛋白生物信息學分析。該研究主要使用了生物信息學分析網站(表2)來預測和分析了編碼蛋白質理化性質、蛋白跨膜區域、信號肽、親水性、疏水性、亞細胞定位、磷酸化位點、蛋白質二三級結構及系統進化樹,以期能夠為后續分析該基因的功能提供基礎數據。其中進化樹構建中各物種氨基酸序列來自NCBI數據庫,分別為紅鰭東方鲀(NP_001027941.1)、人(NP_002469.2)、斑馬魚(NP_571337.2)、羅非魚(XP_031601968.1)、金頭鯛(NM_030427111.1)、大口黑鱸(XP_643446.1)、非洲爪蟾(NP_001079366.1)、草魚(AFL56774.1)、大鼠(NP_788268.2)、豬(NP_001002824.1)、雞(NP_989545.2)、黃牛(NP_001035568.2)、綿羊(NP_001009390.1)、抹香鯨(XP_028357079.1)。
2 結果與分析
2.1 暗紋東方鲀總RNA提取及檢測
使用Trizol法提取暗紋東方鲀肌肉的總RNA,其凝膠電泳結果顯示,提取的RNA效果比較好,沒有蛋白的污染。圖1所展示的就是凝膠電泳檢測條帶,5S、18S和28S這3條帶明亮清晰,且寬度也比較適宜,再通過使用分光光度儀進行檢測,結果顯示D260nm/D280nm的值位于1.8~2.0,表明暗紋東方鲀肌肉的RNA提取效果良好,可以用于之后的試驗研究。
2.2 基因克隆
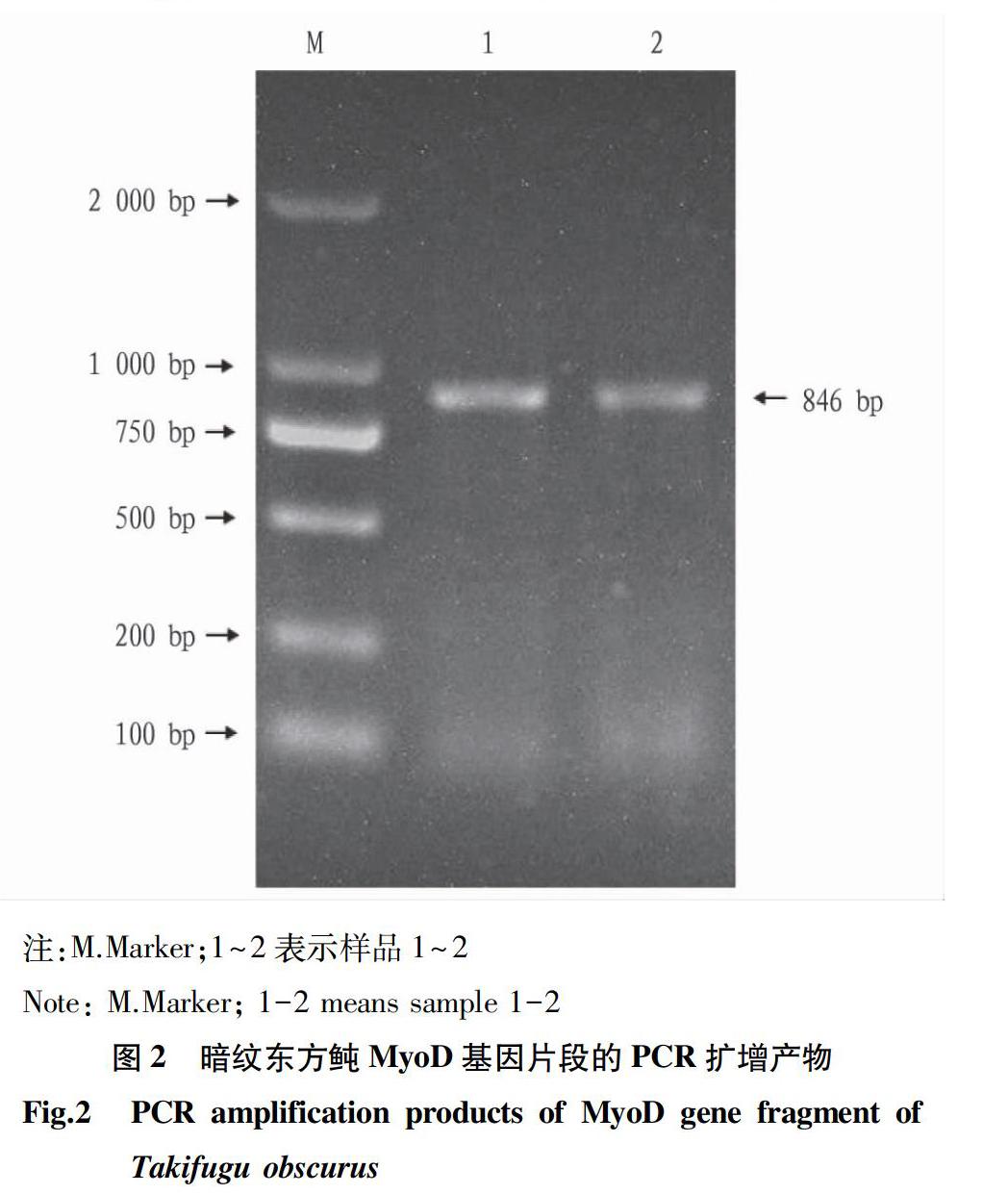
通過PCR程序擴增所得到的目的產物的凝膠電泳檢測結果如圖2所示。在750~1 000 bp出現了一條清晰的DNA特異條帶,與預測擴增片段大小一致。由北京六合華大基因科技有限公司測序,結果所得目的基因片段長度為846 bp。將測序所得結果通過NCBI中的Blast程序與紅鰭東方鲀基因組數據庫該基因全長序列進行對比后,二者相似度高達99%,僅在412 bp處發生1個堿基的變化(A→G),對應所編碼的氨基酸也由蘇氨酸變成了丙氨酸。
2.3 生物信息學分析
通過反轉錄和RT-PCR對暗紋東方鲀MyoD基因的cDNA進行了克隆,得到了暗紋東方鲀MyoD基因CDS區序列。該編碼區長度一共為846 bp,起始密碼子和終止密碼子分別為ATG、TAA。通過Primer Premier 5.0將核苷酸序列翻譯成氨基酸序列,共翻譯編碼281個氨基酸,對其所編碼蛋白繼續進行生物信息學分析。
2.3.1 蛋白質理化性質分析。登陸ProtParam(https://web.expasy.org/cgi- bin/protparam/ protparam)分析MyoD基因氨基酸理化性質,結果表明,氨基酸總數為281,其中Ser、Leu和Arg含量較多,分別為16.0%、8.5%和7.8%,不含Pyl和Sec,帶負電的殘基總數(Asp + Glu)為35,帶正電的殘基總數(Arg + Lys)為32,分子式為C1321H2078N398O436S14,分子量為30 960.28 Da,脂肪指數為62.56,理論等電點為6.41,親水性平均值(GRAVY)為-0.667,半衰期為30 h,不穩定指數為70.51,為不穩定蛋白。
2.3.2 蛋白跨膜區域預測。
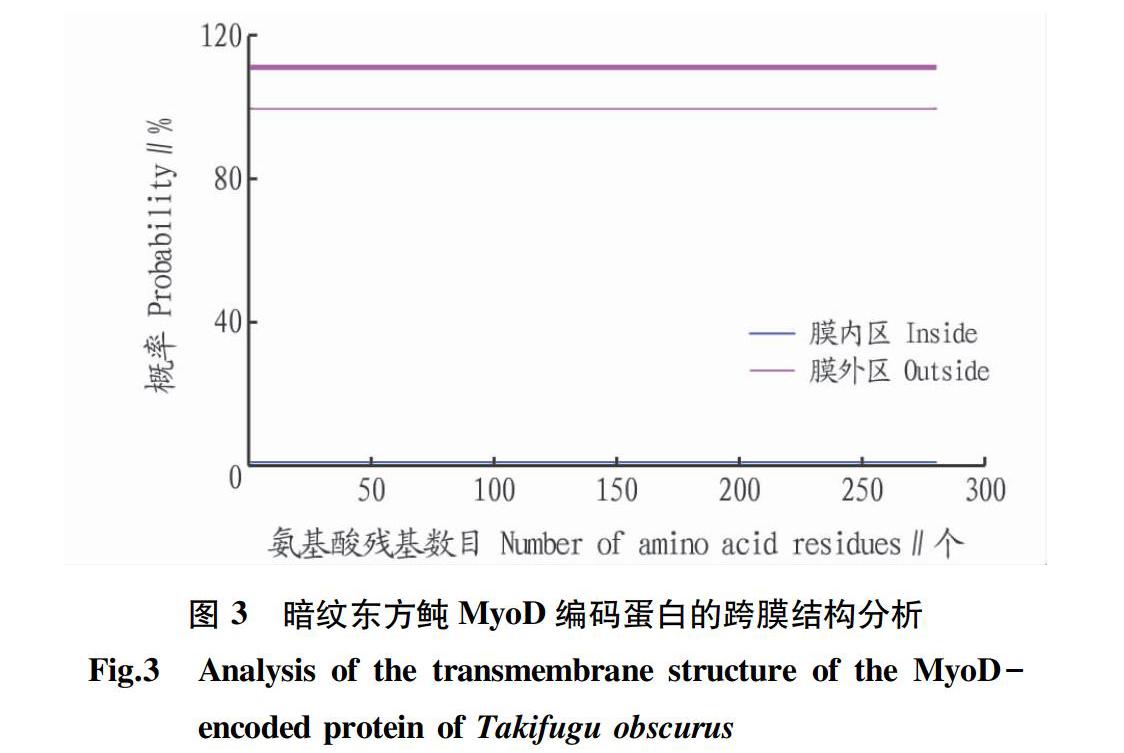
通過TMHMM Sever v.2.0(http://www.cbs.dtu.dk/ services/TMHMM/)輸入暗紋東方鲀MyoD蛋白序列。暗紋東方鲀MyoD基因編碼蛋白跨膜結構如圖3所示,其中X軸代表氨基酸殘基的數目,Y軸表示跨膜結構可能性的分值(概率);紅色線(transmembrane)代表跨膜區;藍色線(inside)代表在膜內部的概率,概率極低;粉色線(outside)代表在膜外的概率,幾乎100%。由此可見,該蛋白不存在跨膜區,且該蛋白全部在膜外。
2.3.3 暗紋東方鲀MyoD基因編碼蛋白的信號肽分析。
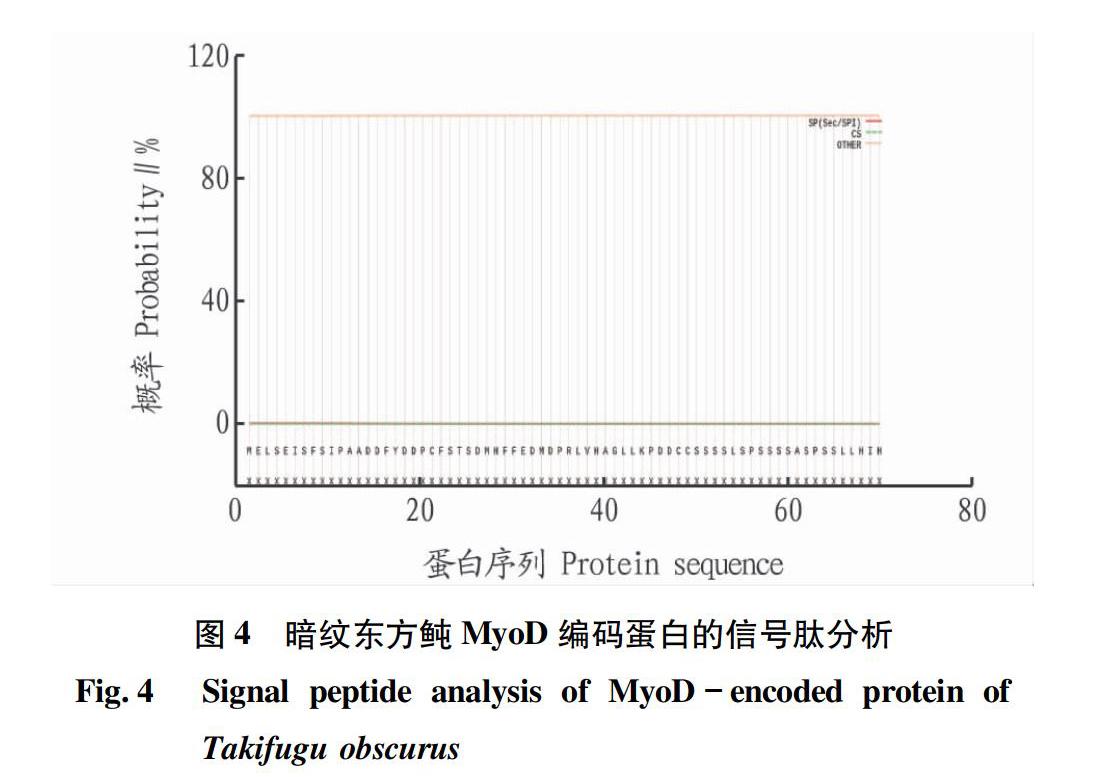
利用軟件SignalP 5.0在線分析蛋白質的信號肽,發現無信號肽(圖4),說明MyoD基因所編碼的不是分泌蛋白。
2.3.4 暗紋東方鲀MyoD蛋白的親水性/疏水性預測與分析。
利用位于Expasy的ProtScale在線分析軟件進行蛋白質的親疏水性分析,Amino acid scale選擇默認的Hphob./Kyte & Doolittle,Window size設置為9,線性加權模型(圖5)。圖形的高峰值(正值)的區域體現的是疏水區域,而負值的“低谷”區域體現的是親水區域。正值越大的氨基酸具備更大的疏水性,負值越小的氨基酸則具備更大的親水性。通過對圖5的分析可得,該蛋白的氨基酸在97位處有最大值(1.544),在75和76位處有最小值(-2.533),MyoD蛋白表現為親水性。
2.3.5 蛋白亞細胞定位分析。
利用PSORT Ⅱ Prediction軟件對暗紋東方鲀MyoD蛋白進行亞細胞定位分析,結果顯示,該蛋白在細胞核中分布最多,為56.5%;在細胞骨架中分布最少,為8.7%;細胞質和線粒體分別為21.7%和13.0%。
2.3.6 暗紋東方鲀MyoD蛋白的磷酸化位點預測。
利用NetPhos 3.1軟件來預測其磷酸化位點,在暗紋東方鲀MyoD基因編碼的氨基酸中共發現有82個Ser磷酸化位點、15個Thr磷酸化位點、5個Tyr磷酸化位點(圖6)。
2.3.7 基因編碼蛋白質結構預測。
2.3.7.1 蛋白質二級結構預測。
蛋白質的理化性質和各功能取決于其空間結構,利用PSIPRED在線分析軟件對MyoD蛋白二級結構進行預測(圖7),從圖7可以看出二級結構預測的置信度(Conf)、二級結構圖標(Cart)、二級結構字符表示(Pred)和氨基酸序列及編號(AA)。暗紋東方鲀MyoD基因編碼蛋白序列中只含有α螺旋(Helix)、無規則卷曲(Coil)和β折疊(Strand),不含有β轉角以及其他已知的結構域。2.3.7.2 蛋白質三級結構預測。
登陸SWISS-MODEL網站(https://www.swissmodel.ex-pasy.org/)預測該基因編碼蛋白質三級結構,預測結果如圖8所示。
2.3.8 系統進化樹。
使用MEGAX分子遺傳進化分析軟件,對暗紋東方鲀(Takifugu obscurus)和人(Homo sapiens)、斑馬魚(Danio rerio)、羅非魚(Oreochromis aureus)、金頭鯛(Sparus aurata)、大口黑鱸(Micropterus salmoides)、紅鰭東方鲀(Takifugu rubripes)、非洲爪蟾(Xenopus laevis)、草魚(Ctenopharyngodon idella)、大鼠(Rattus norvegicus)、豬(Sus scrofa)、雞(Gallus gallus)、黃牛(Bos taurus)、綿羊(Ovis aries)、抹香鯨(Physeter catodon)14個物種以臨近相連算法構建系統進化樹,紅點為暗紋東方鲀分支。由進化樹可表明,暗紋東方鲀與紅鰭東方鲀的親緣關系最近;其次為羅非魚、大黑口鱸、金頭鯛。
3 討論與結論
MyoD基因第一次被成功克隆出來是由Davis在1987年所完成的。作為生肌調節因子MRFs家族的主要成員之一,MyoD基因不論是對骨骼肌的形成還是分化都起著十分重要的作用[17-19]。該研究首次克隆并測出暗紋東方鲀MyoD基因的CDS區核苷酸序列,進而對其進行了生物信息學分析,得到其片段長度為846 bp,共編碼281個氨基酸,這與紅鰭東方鲀MyoD基因編碼區一致。該試驗結果顯示,暗紋東方鲀MyoD基因A、G、T、C堿基分別為21.40%、26.12%、18.44%和34.04%,與紅鰭東方鲀MyoD基因堿基序列相似度為99%,僅在412 bp處有堿基A→G的變化,這一堿基的突變引起了氨基酸序列的改變,因此該位點突變應為錯義突變。將該序列與其他物種的MyoD基因進行同源性比較,結果發現暗紋東方鲀與紅鰭東方鲀同源性最高,與哺乳動物的同源性較低。這與傳統的生化特征分類和形態學分類的進化地位是相符合的,進一步說明 MyoD基因在對物種與物種間的系統進化關系的研究中是具有一定價值的[20-24]。
生物信息學分析結果顯示,暗紋東方鲀MyoD基因分子量為30 960.28 Da,理論等電點為6.41,親水性平均值(GRAVY)為-0.667,脂肪指數為62.56,半衰期為30 h,不穩定指數為70.51,其多肽鏈表現出親水性,為不穩定蛋白。通過軟件TMHMM和SignalP 5.0在線分析可知,MyoD蛋白全部在膜外且不存在跨膜區也無信號肽,說明其不是分泌蛋白,這個結果與MyoD基因只在骨骼肌細胞特異表達的組織特異性一致。二級結構預測結果顯示,暗紋東方鲀MyoD基因編碼蛋白序列中只含有α螺旋(Helix)、無規則卷曲(Coil)和β折疊(Strand),不含有β轉角以及其他已知的結構域。由于暗紋東方鲀MyoD三級結構僅有部分模型,因此無法預測完整的三級結構。在線預測發現,MyoD基因存在磷酸化位點共102個,其中有82個Ser磷酸化位點、15個Thr磷酸化位點、5個Tyr磷酸化位點,由此可以敲定,MyoD 蛋白在肌肉與骨骼生長發育中所起作用在一定程度上與這些磷酸化位點有著緊密聯系,其亞細胞主要定位在細胞核中。
該試驗通過RT-PCR克隆了MyoD基因的編碼區,并對其進行了基礎的生物信息學分析,為以后對該基因的結構和功能的深入研究奠定了一定基礎,以期能夠為改良暗紋東方鲀的肉質作出貢獻。
參考文獻
[1]
SABOURIN L A,RUDNICKI M A.The molecular regulation of myogenesis[J].Clinical cenetics,2000,57(1):16-25.
[2]ALVES H J,ALVARES L E,GABRIEL J E,et al.Influence of the neural tube/notochord complex on MyoD expression and cellular proliferation in chicken embryos[J].Brazilian journal of medical & biological research,2003,36(2):191-197.
[3]ZOU G W,ZHU Y Y,LIANG H W,et al.Association of pituitary adenylate cyclase-activating polypeptide and myogenic factor 6 genes with growth traits in Nile tilapia (Oreochromis niloticus)[J].Aquaculture international,
2015,23(5):1217-1225.
[4]ZHONG X,WANG Q Q,LI J W,et al.Ten-eleven translocation-2 (Tet2) is involved in myogenic differentiation of skeletal myoblast cells in vitro[J].Scintific reports,2017,7:1-11.
[5]于凌云,白俊杰,葉星,等.大口黑鱸MyoD cDNA的克隆和序列分析[J].生物技術通報,2008(S1):301-306.
[6]項露頡,孫麗敏,姜懷志.生肌決定因子(MyoD)基因家族調控動物生產性能發揮的研究概述[J].現代畜牧獸醫,2016(2):54-58.
[7]王立新.草魚MyoD基因cDNA克隆和表達研究[D].楊凌:西北農林科技大學,2005.
[8]AKIZAWA Y,KANNO H,KAWAMICHI Y,et al.Enhanced expression of myogenic differentiation factors and skeletal muscle proteins in human amnion-derived cells via the forced expression of MYOD1[J].Brain & development,2013,35(4):349-355.
[9]BLUM R,DYNLACHT B D.The role of MyoD1 and histone modifications in the activation of muscle enhancers[J].Epigenetics,2013,8(8):778-784.
[10]LIU C,MCFARLAND D C,VELLEMAN S G.Effect of genetic selection on MyoD and myogenin expression in turkeys with different growth rates[J].Poultry science,2005,84(3):376-384.
[11]AVEROUS J,GABILLARD J C,SEILIEZ I,et al.Leucine limitation regulates myf5 and myoD expression and inhibits myoblast differentiation[J].Experimental cell research,2012,318(3):217-227.
[12]成嘉,褚武英,張建社.魚類肌肉組織發生和分化相關基因的研究進展[J].生命科學研究,2010,14(4):355-362.
[13]盧中華,俞菊華,李紅霞,等.奧利亞羅非魚、尼羅羅非魚MyoD1和MyoD2基因特征及差異[J].中國水產科學,2010,17(5):903-912.
[14]蔣天明,羅興春,吳江.暗紋東方鲀養殖技術[J].淡水漁業,2001,31(4):14-17.
[15]馬愛軍,陸麗君,陳超,等.東方鲀屬主要經濟魚種繁育養殖、育種和基因研究現狀[C]//第三屆全國現代生態漁業管理與技術研究.北京:中國水利技術信息中心,2011.
[16]廖章斌,徐后國,衛育良,等.河鲀營養研究進展及展望[J].動物營養學報,2018,30(4):1286-1296.
[17]KABLAR B,KRASTEL K,TAJBAKHSH S,et al.Myf5 and MyoD activation define independent myogenic compartments during embryonic development[J].Developmental biology,2003,258(2):307-318.
[18]DAVIS R L,WEINTRAUB H,LASSAR A B.Expression of a single transfected cDNA converts fibroblasts to myoblasts[J].Cell,1987,51(6):987-1000.
[19]STUHLMILLER T J,GARCA-CASTRO M I.Current perspectives of the signaling pathways directing neural crest induction[J].Cellular and molecular life sciences,2012,69(22):3715-3737.
[20]劉宏祥,徐文娟,宋衛濤,等.高郵鴨群體 MSTN、MyoD1和 MyoG 基因外顯子中SNP位點的分析[J].江蘇農業學報,2015,31(3):604-612.
[21]王立新,白俊杰,葉星,等.草魚My5oD cDNA的克隆和序列分析[J].中國農業科學,2005,38(10):2134-2138.
[22]GALLOWAY T F,BARDAL T,KVAM S N,et al.Somite formation and expression of MyoD,myogenin and myosin in Atlantic halibut (Hippoglossus hippoglossus L.) embryos incubated at different temperatures:Transient asymmetric expression of MyoD[J].Journal of experimental biology,2006,209(13):2432-2441.
[23]FERNANDES J M O,KINGHORN J R,JOHNSTON I A.Differential regulation of multiple alternatively spliced transcripts of MyoD[J].Gene,2007,391(1/2):178-185.
[24]ANDERSEN ?,DAHLE S W,VAN NES S,et al.Differential spatio-temporal expression and functional diversification of the myogenic regulatory factors MyoD1 and MyoD2 in Atlantic halibut (Hippoglossus hippoglossus)[J].Comparative biochemistry & physiology part B:Biochemistry & molecular biology,2009,154(1):93-101.
