花器官液泡pH調控花色形成的研究進展
2021-05-14 06:01:36張蓋天祁惠楊鎖寧褚志云田甜袁素霞劉春
生物技術通報 2021年4期
張蓋天 祁惠 楊鎖寧 褚志云 田甜 袁素霞 劉春
(中國農業科學院蔬菜花卉研究所,北京 100081)
觀賞器官形狀、色彩以及香氣是開花植物吸引傳粉動物、維持繁衍進化基石中最重要的幾個因素[1]。花色的成因背景十分復雜,由顯色物質、著色細胞生理環境、著色細胞形狀與外界環境如光照、雄蕊完整性及授粉情況等因素共同決定,其中顯色物質被認為是最重要的因素[2-3],因此有關花色研究的大部分內容主要集中在顯色物質上[4-6]。顯色物質主要有類胡蘿卜素、類黃酮(包括花青素)等色素及金屬離子構成,其中色素,尤其是花青素是花和果實呈色的主要的顯色物質[2]。花青素主要有6大種,分別為天竺葵素、矢車菊素、芍藥花素、矮牽牛素、飛燕草素和錦葵花素,其中芍藥素派生于矢車菊素,矮牽牛素與錦葵花素派生于飛燕草素。藍花多含飛燕草素及其衍生物,紅花往往含天竺葵素。花色苷修飾過程中,會先發生糖基化,后可在糖基化位點上經過酶促反應進一步酰基化,使花色苷本身更為穩定。此外,B-ring上羥基增多會使由花青素衍生出的色素顏色變藍,而3′,5′羥基甲基化會使色素顯紅[7]。Noda等[8]進一步研究發現,在利用轉基因技術培育藍色菊花的過程中,在相同的遺傳背景下,介導入相同的基因后,分別出現了紫羅蘭色到藍色的不同顏色的菊花,在藍色的菊花中,飛燕草素-3′,5′-葡萄糖苷的占比要顯著高于紫羅蘭色的菊花。
花色苷單體在介質中會出現紫色而非藍色,輔色素與花色苷形成共色素沉著后,能夠維持顯色物質結構穩定,可呈現出藍色[9]。在體外模擬八仙花萼片成色實驗中發現,不同輔色素在相同的溶液背景中,呈現出淺紫、深紫、藍這3種不同的顏色[10],這說明不同輔色素結合其它色素物質后的成色并不相同。
部分植物金屬離子可與沉著物螯合顯示特異的顏色。部分粉色繡球花品種,在酸性土壤條件下施鋁,萼片組織細胞就會吸收過量鋁離子,為避免鋁毒害,經鋁調蛋白作用將鋁離子轉運至液泡內,鋁離子在液泡內與共色素沉著物形成螯合物后,花瓣顯現藍色[11]。郁金香品種“Murasakizuisho”的花瓣基底部為藍色,上半部為紫色,測定發現這兩部分花色苷、輔色素、液泡pH均無顯著性差異,但藍色部分的Fe3+含量是紫色部分的25倍[12]。鴨跖草藍花的色素物質,是由2個Mg2+結合6個飛燕草素花色苷、2個Mg2+結合6個類黃酮,而形成的的超分子配合物[13]。矢車菊蘭花的色素類物質,是由1個Fe3+、1個Mg2+分別結合3個矢車菊花色苷、2個Ca2+分別結合3個芹黃素而形成的超分子配合物[14]。
液泡是由液泡膜包被的充滿液體的區室,也是植物儲存色素的器官,大而成熟的細胞中液泡體積占細胞體積的90%[15]。液泡內顯色物質的穩定性及存在狀態由著色細胞生理環境所決定[16]。研究表明,隨著液泡pH的升高,部分植物花器官會出現由粉、紅到紫、藍的轉變[17]。這種顏色變化現象的產生與花色苷類物質存在的狀態密切相關,不同狀態的花色苷類物質吸收光譜有不同的紅移或藍移效應[16]。
1 花器官液泡pH對花色的影響
在花青素類觀賞植物中,顯色物質組成相同,顯色物質存在的狀態不同,呈現的花色不同。保持這類植物顯色物質穩定性的方式有花色苷的自締合作用和花色苷與輔色素結合的共色素沉著作用。自締合作用中多個花青素發色團疊加在同一個左旋軸上,不同角度的遷移導致分裂的圓偏光二色性曲線產生,即光譜有不同的紅移或藍移趨勢[16](圖1);共色素沉著是顯色物質疏水堆積使結構穩定,對花色苷與輔色素濃度、種類有要求,即濃度越高,疏水力越強,顯色物質越穩定,顏色可由紅向藍紫移動。這些全部依賴于一定的液泡pH,液泡pH改變時水合能力改變,著色物質為適應水合環境,調整自身使其穩定,顏色發生改變[10,18]。合適的花色苷與輔色素形成共色素沉著后,還可避免花色苷不穩定的無水堿基形成甲基醇從而不顯色,且pH在3-6的區間內輔色素槲皮苷才能和花色苷、金屬離子形成絡合物[19]。
大豆紫花胞液pH為5.73-5.77,而藍花胞液pH為6.07-6.10[20]。類似地,矮牽牛紅色花花瓣勻漿pH在5.5左右,而紫色花瓣勻漿pH在6.0左右[21];飛燕草花瓣表皮組織在紫紅色時pH為5.5,紫藍色時pH為6.6[22]。另外,一些植物花瓣液泡的pH也顯示出類似的變化趨勢。日本牽牛從花蕾到綻放,花瓣液泡pH一直升高,花蕾時期紫紅色的細胞液泡pH為6.6,而開花后藍色細胞液泡的pH為7.7[23-24];同樣,在八仙花上,用質子選擇微電極精準測量著色細胞液泡,藍色花的液泡pH均值為4.1,而紅色花為3.3[25]。葡萄風信子的穗狀花序上半部分為藍色,下半部分為紫色,紫色部分的勻漿pH為5.84,藍色部分為5.91[26]。葛根花序上同時有不同時間開放的花,新開的花為紫紅色,pH為5.2,老花為紫羅蘭色,pH為5.5[27]。
自然界中沒有藍色月季花,月季的DRF基因無法將二氫楊梅酮作為底物反應生成飛燕草素花色苷,致使無藍色月季的誕生。研究人員在試驗了多種月季品種后選擇了輔色素含量和液泡pH均高的品種作為寄主,將三色堇的F3′5′H基因與荷蘭鳶尾的DFR基因構建二元載體導入寄主,在月季DFR基因SiRNA表達的共同作用下,寄主花瓣最終呈現淡紫色。這對藍色月季的培育具有里程碑式的意義[28]。由此可以得出結論,除飛燕草素花色苷外,較高的液泡pH與高含量的輔色素也是藍色月季花形成必不可少的元素,這兩個重要因子在在仙客來、菊花和康乃馨上同樣有所印證[28-29]。
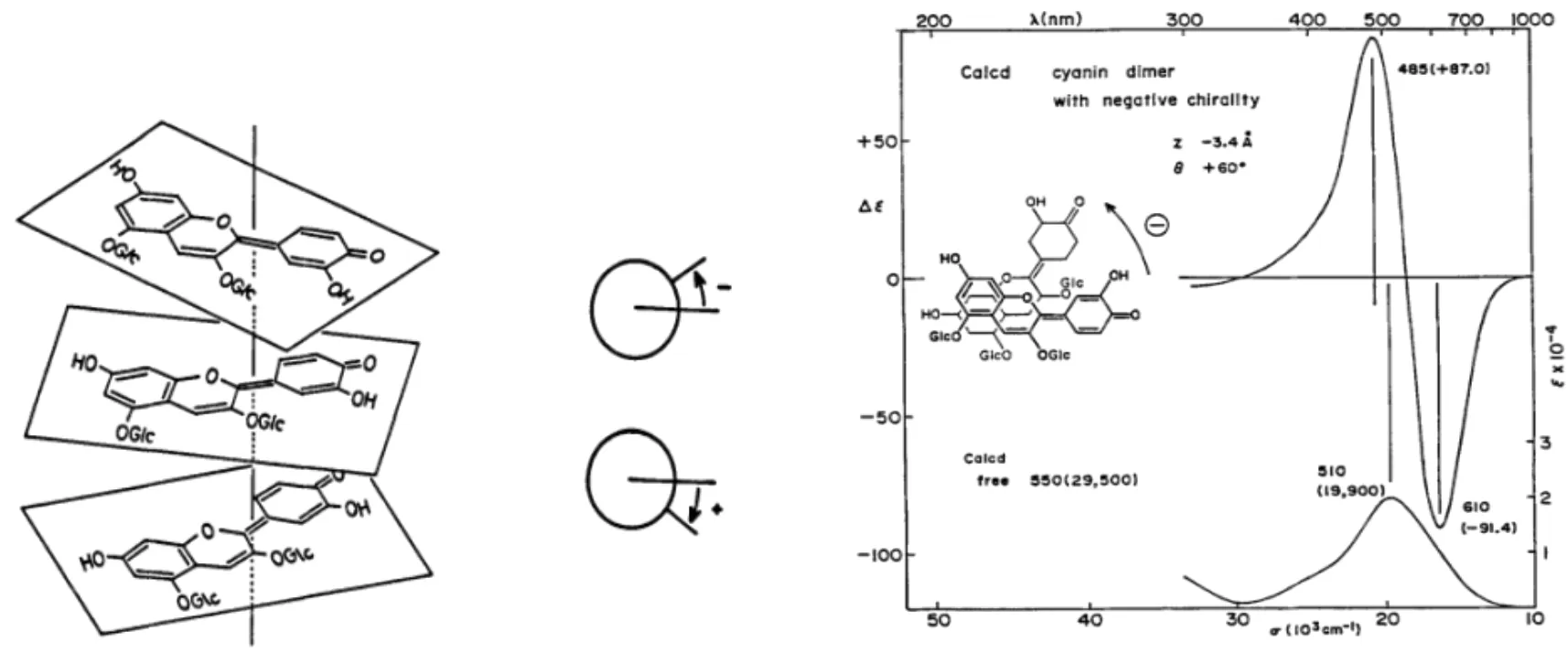
圖1 花色苷堆積偏旋模式及CD曲線展示[17]Fig. 1 Anthocyanins stacking pattern and CD curve display[17]
2 調控花器官液泡pH的分子機理
Wilson等[30]在水稻和玉米上發現,將原來的pH為5.0的營養液調至pH為8.5,兩種作物根部液泡pH均出現一定的下降趨勢,5-10 min 后恢復原來的pH;在玉米營養液中加入NH3,使營養液pH升至8.5,玉米根部液泡pH由5.5左右升至6.1左右,并穩定下來;在水稻營養液中加入NH3,水稻根部液泡pH由5.4左右升至5.9左右,很快恢復至pH為5.4左右,并趨于穩定,這表明液泡具有一套隔離外界環境、獨立調控內部pH的系統。
由于著色細胞液泡的pH在花色調控中起重要作用,因此研究調控花器官液泡pH的分子機理對于花色調控具有非常重要的價值。前人在調控著色細胞液泡pH的研究中,以日本牽牛與矮牽牛作為模式植物,闡明了著色細胞液泡堿化與酸化的分子機理[31]。
2.1 液泡酸化
野生型矮牽牛花冠呈紅色,矮牽牛突變體出現與原有表型有色差的扇形色塊,或花冠整體顏色偏紫。研究發現,不同顏色區域,其液泡pH也不同。
通過進一步分離雜交實驗,共分離出PH1-PH7 7個調控著色細胞液泡pH酸化的基因[32-34]。其中PH4編碼R2R3 MYB蛋白(myeloblastosis 蛋白),PH6(后更名為AN1)屬于bHLH 蛋白(basic helixloop-helix 蛋白),這兩個蛋白與WDR蛋白(WD是由40個氨基酸組成,以色氨酸W、天冬氨酸D結尾的結構域;WDR是WD重復)AN11結合,形成MBW(MYB-bHLH-WD)復合物,調節花青素晚期合成通路;PH4-AN1-AN11還激活PH3轉錄,PH3編碼WRKY蛋白(含WRKYGQK 7個保守的氨基酸序列和16個氨基酸的鋅指結構域的轉錄因子),可與AN11綁定,與PH4-AN1-AN11形成復合蛋白,在前饋環中轉錄PH5,同時,PH1也受PH4-AN1-AN11 以 及 PH3 的 調 控[20-21,35-37]( 圖 2)。PH5 表達還受AN2轉錄激活調控;PH1、PH5表達均略晚于DRF基因的表達,可能是因為PH1、PH5表達需要花青素積累;PH1、PH5表達不影響花青素種類、數量及結構的變化,不影響細胞形狀[38-40]。
矮牽牛在PH3調控下花冠呈紅色,ph3突變體中,PH1、PH5表達減少,花冠表皮著色細胞液泡pH升高,顏色呈灰紫色。即使PH5過表達,也不足以使突變體表型恢復正常,但是當PH1也同時過表達時,ph3突變體可以恢復正常表型。這說明PH1、PH5形成復合體后,向花冠表皮著色細胞液泡泵入H+的能力成倍提高[40]。PH5編碼一個P3AATPase質子泵,該質子泵主要負責向花冠表皮著色液泡泵入H+。PH1編碼的P3B-ATPase,缺乏陽離子結合與易位的關鍵——保守天冬氨酸殘基,故不能行使質子泵功能,故液泡酸化過程中作為PH5的輔因子,與PH5形成雜聚肽復合物,共定位于花冠表皮著色液泡上,增強PH5轉運H+的能力,實現液泡高酸化[41]。PH1、PH5的同源基因廣泛分布于被子植物中[31]。

圖2 PH基因編碼蛋白調控液泡酸化可能的模式圖Fig.2 The possible pattern of vacuole reduced pH by PH genes encoded protein
除控制液泡酸化之外,PH基因還在花青素合成通路、花器官著色細胞小型液泡類似物vacuolino與中央大液泡融合以及花器官發育等其它方面起著特殊作用。
PH1與膜蛋白運輸有關。Faraco等[42]在矮牽牛花冠著色細胞與月季的花瓣表皮著色細胞中,發現了一類在花器官表皮著色細胞中存在、而在葉肉細胞中并不存在的液泡類似物vacuolino。復合質子泵PH1-PH5出現在vacuolino膜上,中央大液泡上的蛋白受體識別PH1后,二者形成鏈栓結構,相互融合,PH1-PH5進入之前無這兩種蛋白的中央大液泡,這是一種膜蛋白運輸方式的分支。矮牽牛突變體ph3和ph4中,vacuolinos無法產生;ph1突變體中vacuolinos與中央大液泡無法融合。
矮牽牛PH3基因與擬南芥TTG2基因高度同源且功能上可以互相代替,由于TTG2參與植株毛狀物的形成以及原花青素積累和液泡酸化,因此PH3基因除了液泡酸化功能外,也參與植株毛狀物的形成以及原花青素積累[35]。
矮牽牛中純合ph3會導致雌性不育,且會遏制F3′5′H基因的表達;花褪色顯性等位基因Fa,只在ph3ph3和ph4ph4背景下表達[41];隨著矮牽牛花冠的老化,變紫的背景下出現的紅色斑點與扇區和ph7 有關[33]。
另外,PH基因還參與果實風味調控。從柑橘果實液囊中分離出CsPH5,CsPH5為PhPH5的同源基因,主要負責向果肉液囊中泵入氫離子,提高果實風味[43]。但甜瓜中的PH基因編碼的蛋白相似性最高的蛋白是PINs家族的蛋白,即H+/auxin泵,且該蛋白定位于內質網中,基因沉默后并未發生生長素缺失的表型[44]。
對這兩種調控植物花器官與果實器官酸度的不同基因家族的研究表明,不同植物、不同部位PH基因需要深入研究其功能與意義。
2.2 液泡堿化
NHX基因屬于CPA(一價陽離子逆轉蛋白)家族,該家族廣泛的分布于細菌、真菌及高等動植物中,參與調控細胞周期與增殖,耐鹽性,囊泡販運與生物發生[45]。研究發現,NHX1基因編碼的蛋白被定位于液泡上,可以使液泡外Na+/K+置換液泡內H+,使得液泡pH升高。此前有關NHX1的研究大多集中于抗鹽脅迫,除此之外它還具備了調控植物花色的本領[46]。
日本牽牛從花蕾到開放顏色由粉至紫或藍的過程中,色素類物質并不改變,僅液泡pH不斷地在提高[47]。Fukada-Tanaka等[24]發現,日本牽牛花的Pr基因負責升高液泡pH使花冠變藍,Pr基因與擬南芥和水稻Na+/H+交換器閱讀框高度同源,且能補充酵母nhx1突變,故命名 Pr基因編碼的蛋白為lnNHX1。lnNHX1也是第一個被鑒別出來調控液泡pH升高使花色變藍的蛋白。lnNHX1基因在開花前12 h的花瓣中表達最豐富,開花0 h時lnNHX1所編碼的蛋白含量最高,在這一液泡pH升高的過程中,并沒有鹽脅迫的情況發生[23]。
進一步的研究表明,僅在日本牽牛開花時的著色細胞液泡膜上觀察到lnNHX1,其是使液泡堿化花瓣變藍的主要原因[48]。實驗結果也表明,除lnNHX1外可能沒有其它主要基因在花期促進液泡堿化,盡管也有研究表明部分lnNHX1缺失突變體可以在開花時花枝顏色變紫,但這一現象被推測為是其它一些基因參與協作的結果。此外,lnNHX2主要在葉片、莖、根中表達,研究表明lnNHX2也可使牽牛花開放時部分細胞液泡堿化花瓣變紫[49],說明lnNHX2也具有液泡堿化的部分功能。
3 展望
花色研究一直是觀賞植物研究的重點領域,花器官有色細胞液泡pH作為影響花色呈現的重要因素,其研究對象受限于以下兩點:第一,試驗材料顏色不同并非因色素類物質種類、含量不同;第二,試驗材料顏色不同并非因有色細胞形狀發生變化(如皺縮等)。所以,研究花器官液泡pH對花色影響的植物種類十分重要。目前,有關花器官液泡pH對花色的分子調控在日本牽牛和矮牽牛中的研究較為深入。在矮牽牛中,而PH1與PH5在液泡pH酸化過程中起關鍵作用;而在日本牽牛中,液泡的pH變化主要依靠NHX1行使功能。
但是,有色細胞液泡pH參與調控其他觀賞植物花器官顏色變化的機理研究相關資料甚少,這導致我們對于有色細胞液泡pH變化的機理不甚了解。從現有研究來看,在矮牽牛中,有色細胞液泡pH調控的相關基因與花色苷形成和其育性密切相關[28-32]。這證明了在進化過程中,不論是有色細胞液泡pH調控還是花色苷形成均與植物繁殖有關。
綜上所述,有關于調控花器官有色細胞液泡pH的基因及其功能,挖掘還不夠全面,部分基因僅有名稱報道,但未見功能驗證,故有關的pH基因有待于進行深入研究,以期為進一步良種繁育以及創建花色形成的環境調控技術體系奠定理論基礎。
隨著基因工程育種體系的進一步發展,轉基因技術在觀賞植物上的應用逐漸增多,科研與商業化要求對基因在現有植株中的功能及物種進化中的作用需要有非常清晰的認知,以方便開展轉基因及人為促進物種進化的相關工作。研究花器官著色細胞液泡pH對花色的調控,明確液泡pH相關基因功能及花色調控機制,在觀賞植物的花色調控和分子育種中具有重要的意義。
