甲烷作為電子供體合成聚羥基脂肪酸酯的研究進展
2021-05-20 12:12:26岳穎蓉方芳徐潤澤操家順
應用化工 2021年4期
岳穎蓉,方芳,徐潤澤,操家順
(1.河海大學 淺水湖泊綜合治理與資源開發教育部重點實驗室,江蘇 南京 210098;2.河海大學 環境學院,江蘇,南京 210098)
聚羥基脂肪酸酯(Polyhydroxyalkanoates,PHA)是由微生物合成的一種胞內聚酯,其具有與傳統石油化工類塑料相似的性質,可廣泛應用于工業、農業、建筑、制藥及生物醫學等領域[1]。雖然人們廣泛關注PHA的生產,但是高成本的碳源限制了PHA市場的進一步發展[2]。甲烷是自然界中一種重要的溫室氣體,具有很高的化學穩定性和很強的溫室效應。大部分的甲烷排放源于人工,主要有化石燃料、畜牧業、厭氧廢物處理、煤炭開采和廢水處理等[3]。目前研究表明甲烷氧化菌可以以甲烷為唯一碳源和能源,在營養限制條件下合成生物聚合物PHA[4]。因而將甲烷作為低成本的原料生物轉化為高附加值的PHA,既可以增加污水中資源的回收,又有助于緩解氣候變化問題。
近年來,研究者們已經就甲烷氧化菌利用甲烷作為電子供體合成PHA進行了大量的研究,并取得了一定的進展。但是,大多數研究仍局限于小試實驗,未進行大規模的實際應用,且合成的PHA種類較為單一。因此,本文首先就PHA的性質及應用展開闡述;然后綜述了關于甲烷氧化菌合成PHA的能力與生物代謝途徑的研究,并強調了不同環境因素對甲烷氧化菌利用甲烷合成PHA的影響;最后針對目前研究存在的問題提出了相應的策略,為未來研究利用甲烷高效合成PHA提供解決思路。
1 聚羥基脂肪酸酯(PHA)
1.1 PHA 的結構及分類
聚羥基脂肪酸酯(PHA)是一種由微生物在碳源充足而N、P、微量元素等營養限制的條件下合成的細胞內聚酯[5]。其顆粒大小為0.2~0.5 μm,可作為碳源和能源貯存物,在碳源不足的情況下進行利用。PHA主要由羥基脂肪酸(HA)聚合而成,大多數單體的鏈長為3~14個碳原子,側鏈為高度可變的芳香族或脂肪族基團[6]。根據單體碳鏈長度的不同,可將PHA分為短鏈、中鏈和長鏈三類。短鏈PHA的碳原子數為3~5個,在古細菌、革蘭氏陽性和陰性細菌中廣泛分布,可形成常見的聚合物,如聚羥基丁酸酯(PHB)、聚羥基戊酸酯(PHV)等;中長鏈PHA的碳原子數為6~14個,大多存在于假單胞Pseudomonas中;長鏈PHA的碳原子數超過14,但并不常見且研究較少。此外,根據單體連接樣式不同,可將PHA分為均聚、無規共聚、嵌段共聚三類。均聚PHA中只含有一種單體,無規共聚PHA由多種單體隨機聚合而成,嵌段共聚PHA由多個鏈段構成,且每個鏈段只含有一種單體[7]。
1.2 PHA的性質及應用
由于PHA的單體結構具有可變性,使得其物理化學性質和機械性能,如韌度、脆性、熔點、玻璃態溫度和抗溶劑性等,都會隨著單體數量和組成的變化而發生改變[8],可根據不同的需求針對性地選擇不同的PHA。短鏈PHA結晶度較高,可制得性質堅硬而脆的硬塑料;而中長鏈PHA結晶度較低,可制得性質韌而軟的彈性體塑料。通過物理和化學等方法還可改變PHA結晶性能,提高其機械性能,從而拓寬PHA的應用范圍。PHA還具有優良的生物相容性和生物可降解性。PHA良好的生物相容性表現在它能夠為多種組織器官細胞提供生長環境,不會導致癌癥且大多可轉化為動物體內本身存在的物質。因此,將PHA材料用作醫療植入性材料和用于藥物的可降解載體具有極大的潛力[9]。PHA的生物降解性表現為不同的微生物會產生不同的PHA解聚酶來降解PHA[10],在自然條件下可被微生物完全降解。
2 甲烷作為電子供體用于PHA合成
2.1 甲烷氧化菌
甲烷氧化菌是一類利用甲烷作為碳源和能源的革蘭氏陰性菌,在好氧條件下可將甲烷轉化為生物質、二氧化碳和生物聚合物[11]。甲烷氧化菌廣泛分布在濕地、森林、海洋、湖泊、稻田、煤礦、地下水、垃圾填埋場、污水污泥等各種環境中。甲烷氧化菌根據其生理和形態特征分為三類:Ⅰ型、Ⅱ型和Ⅹ型甲烷氧化菌[11-12]。Ⅰ型甲烷氧化菌通過核糖磷酸(RuMP)途徑進行碳同化;Ⅱ型甲烷氧化菌利用絲氨酸途徑進行碳同化;而Ⅹ型甲烷氧化菌具有Ⅰ型和Ⅱ型甲烷氧化菌的特征,通過RuMP途徑吸收甲醛,并利用絲氨酸途徑固定CO2[13]。
當前已知的甲烷氧化菌可基于16 S rRNA分為兩個不同的門,即變形菌門(細分為α-和γ-Proteobacteria)和疣微菌門[14]。α-Proteobacteria門中最有代表性的屬是Methylocystis、Methylosinus、Methylocella和Methylocapsa,它們屬于Methylocystaceae和Beijerinckiaceae科,為Ⅱ型甲烷氧化菌;而γ-Proteobacteria門中的甲烷氧化菌有Methylomonas、Methylobacter、Methylosarcina和Methylococcus等屬,它們屬于Methylococcaceae科,為Ⅰ型甲烷氧化菌。
2.2 純種或混合甲烷氧化菌合成PHA的能力
早在1970年,Whittenburry等研究發現Ⅱ型甲烷氧化菌具有將甲烷轉化為PHB的能力[12]。此后,眾多研究者對利用甲烷氧化菌將甲烷轉化為PHB展開了大量的研究。不同實驗條件下Ⅱ型甲烷氧化菌積累PHB的能力見表1。
Methylocystis、Methylosinus和Methylobacterium是目前研究較多的3個屬。 其中,Methylocystis屬在不同環境下的混合菌群中均占主導地位,對其的研究最為廣泛[15-17]。MethylocystisparvusOBBP的最大PHB積累量可達到36%~70%,而Methylocystishirsuta的PHB積累量變化范圍較大,為7%~85%[4,15,18-19]。Methylosinus和Methylobacterium是另外兩個PHB積累量較高的屬[18,20-21]。從不同環境樣品分離到的Methylosinustrichosporium可獲得25%~51%的PHB積累[21-22],在MethylobacteriumorganophilumCZ 2中最多可獲得57%的PHB積累[21]。

表1 甲烷氧化菌的PHB積累能力
除了對純甲烷氧化菌進行培養,人們還可考慮利用混合甲烷氧化菌群合成PHA,從而降低純菌操作的成本,并可獲得相似的PHA積累。在營養限制的條件下,對Methylocystissp.GB25(占混合菌群的86%)進行混合培養,PHB積累可在約24 h內達到最大值(33.6%)[26]。在開放條件下以甲烷為底物培養甲烷氧化菌群,PHB積累量可達到35.5%,且開放培養過程中菌群結構穩定[27]。PHA的積累與甲烷氧化菌的接種種齡直接相關,通常在穩定生長期PHA的積累量達到最高值[16],純種和混合甲烷氧化菌獲得最大PHA積累的時間往往有所不同。研究發現純培養Methylocystisparvus可在170~180 h達到28%的PHB積累[4],而混合菌群可在培養144 h內達到類似的積累效果(27.5%)[28]。同樣,以MethylocystishirsutaCSC1為優勢種的混合菌群中,PHB積累量在接種后80 h內達到了42.5%[16],而純培養MethylocystishirsutaCSC1僅積累了7%的PHB[18]。這表明,相較于純甲烷氧化菌培養,混合甲烷氧化菌群可以在較短的時間內達到較高的PHB積累量。
2.3 甲烷氧化途徑和甲烷氧化菌中PHA的合成
甲烷氧化菌氧化甲烷與合成PHA的途徑見圖1(見后)。甲烷氧化菌通過逐步反應將甲烷氧化為CO2,生成甲醇、甲醛、甲酸作為中間體[29]。甲烷單加氧酶(MMO)可以催化甲烷轉化為甲醇,通常以細胞內膜上的顆粒形式(pMMO)或細胞質內的可溶性形式(sMMO)出現。sMMO一般由Ⅱ型甲烷氧化菌合成,很少有Ⅰ型甲烷氧化菌能夠合成sMMO[13]。首先MMO催化甲烷氧化為甲醇,接著由甲醇生成具有細胞毒性的甲醛。在此階段甲烷氧化菌需要高濃度的還原當量,將甲醛轉化為甲酸并進一步同化為細胞生物質。在Ⅰ型甲烷氧化菌中,3-己糖磷酸合酶催化甲醛合成(D-阿拉伯糖)-3-己糖-6-磷酸,再轉化為不同的中間產物進入RuMP途徑,通過不完全的TCA循環轉化為CO2。而在Ⅱ型甲烷氧化菌中,四氫葉酸催化甲醛氧化進入絲氨酸途徑,隨后在營養充足的條件下進行TCA循環產生CO2。一旦在培養液中出現了營養限制的情況,乙酰CoA進入TCA循環受到限制,Ⅱ型甲烷氧化菌就會將過量的乙酰CoA轉移到PHA/PHB循環中,合成PHA/PHB。首先通過由PhaA基因編碼的3-酮硫解酶催化乙酰CoA轉化為乙酰乙酰CoA。隨后,通過由PhaB基因編碼的乙酰乙酰CoA還原酶將其轉化為3HB-CoA。最后,通過由PhaC基因編碼的PHA合酶將3HB-CoA聚合成P(3HB)[30]。
甲烷的生物降解可通過RuMP途徑和絲氨酸途徑實現,兩個途徑的化學計量如公式(1)和(2)[29]:
RuMP途徑:
0.118(C4H3O2N)+0.529CO2+1.71H2O+0.118H+
(1)
絲氨酸途徑:
0.102(C4H3O2N)+0.593CO2+1.75H2O+0.102H+
(2)
Ⅰ型甲烷氧化菌的RuMP途徑能夠更高效地轉化甲烷,在甲烷氧化過程中形成3個甲醛分子僅需要1個ATP分子。雖然Ⅱ型甲烷氧化菌可以更好地將甲烷儲存為磷脂脂肪酸和生物聚合物(即PHA)[11,14],但它們需要3個ATP和2個NADH分子才能通過絲氨酸途徑固定2個甲醛和1個CO2分子。這表明在混合培養條件下Ⅰ型甲烷氧化菌的生長比Ⅱ型甲烷氧化菌具有優勢,因此在進行PHA積累階段之前,需要選擇適當的培養條件對混合菌群中的Ⅱ型甲烷氧化菌進行富集培養,以排除Ⅰ型甲烷氧化菌。
3 影響甲烷氧化菌利用甲烷合成PHA的因素
甲烷氧化菌中的PHA含量和性質受到多個因素的影響,例如甲烷、O2、氮、磷、微量營養元素、pH、溫度等。為實現PHA的最大化積累,必須對這些相互聯系的影響因素進行解析,使其保持在最佳范圍內。
3.1 甲烷和O2的含量
甲烷氧化菌會受到甲烷濃度變化的影響。相比于Ⅰ型甲烷氧化菌,Ⅱ型甲烷氧化菌對甲烷含量變化的耐受性更高。低濃度的甲烷會使Ⅱ型甲烷氧化菌中的關鍵酶sMMO失活,因此需要其他能源來維持細胞活性并提供還原當量[31],而甲烷的濃度過高也會對PHA的合成產生抑制作用。研究發現在20 g 和2 g CH4/m3下PHB的最大積累量分別達到1.0%和12.6%(w/w)[32]。因此,需要控制甲烷濃度在合適的范圍內,既保證甲烷營養菌與其關鍵酶的活性,又要盡可能提高PHA的積累量。
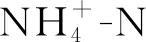
3.2 營養物氮磷含量
同樣,磷源限制可以促進PHA的積累。Methlocystissp.GB25 DSMZ 7674在磷源限制時,其PHA的最大積累量為0.55 g PHB/g CH4[23]。在磷源限制條件下,Methylocystis和Methylobacterium成為優勢屬,且在25 ℃時PHB含量高達(13.6±5.6)%[34]。維持sMMO和甲醇脫氫酶活性所需的最佳P濃度范圍為2~25 mmol/L,而當P濃度>40 mmol/L時會抑制Ⅱ型甲烷氧化菌MethylosinustrichosporiumOB3b的生長[35]。因此,在利用甲烷合成生物聚合物的過程中提供適量的P也至關重要。
3.3 微量營養元素
微量營養元素含量的變化也會對甲烷氧化菌的生長產生影響。研究表明K和Mg的濃度對Ⅱ型甲烷氧化菌積累PHA會產生影響,對K和Mg進行限制可以促進PHA的積累。當培養液中的K濃度控制在17 mg/L時,以Methlocystissp.GB25為優勢菌的混合菌群積累了33.6%的PHB,且在K限制的條件下生成了具有3.1 MDa的超高平均分子量的聚合物[26]。當培養液中的K濃度降至25 mg/L以下時,Methylobacteriumorganophilum進行PHB的積累,培養70 h后獲得的PHB積累量為52%~56%[25]。Mg的添加對sMMO活性、細胞生長和甲烷轉化為甲醇的效率均有積極影響。而在Mg限制條件下,以Methlocystis sp.GB25為優勢菌的混合菌群可以積累28.3%的PHB且生成的PHB分子量高達2.5 MDa[23]。 這表明K和Mg限制不僅有利于PHA的積累,還能產生高分子量的PHA。
Cu和Fe等金屬離子會通過對甲烷氧化菌MMO活性的調節影響其合成PHA。Cu可以控制編碼MMO的基因表達,并對pMMO和sMMO的活性有積極作用[14]。添加的Cu含量需要控制在一定的范圍內。研究表明過高的Cu含量會抑制sMMO的活性和甲烷氧化菌的生長,而一定的初始Cu濃度有利于pMMO活性的表達,有助于細胞中PHB的合成[35]。研究發現加入了30 mol/L Cu后MethylosinustrichosporiumIMV 3011中的pMMO活性提高到未添加Cu的4倍,細胞密度為未添加Cu的2倍,PHB含量增加到8.3%[36]。而Fe含量也會對MMO的活性產生影響,維持sMMO活性的最佳Fe濃度范圍為40~80 μmol/L,然而超過最佳Fe濃度范圍并未觀察到會對MethylosinustrichosporiumOB3b的生長產生顯著影響[35]。較高的Fe濃度可以減輕銅胺絡合物的抑制作用,因此可就Cu和Fe對PHA積累的聯合影響進行研究,以實現穩定且高效的培養。
3.4 溫度和pH
利用純甲烷氧化菌或混合菌群進行大規模培養時,pH值會對PHA的積累產生影響。通常用于富集Ⅰ型和Ⅱ型甲烷氧化菌的培養基pH值介于6.8~7.0。由于Ⅱ型甲烷氧化菌需要CO2作為絲氨酸循環的輸入,而較低的pH值有助于溶解CO2。因此,Ⅱ型甲烷氧化菌對低pH值的耐受性比Ⅰ型甲烷氧化菌更高[18],略酸性的pH對于Ⅱ型甲烷氧化菌的生長和PHA積累量的增加更為有利。在pH值分別為3.6~4.5,5.0~6.0和6.0~6.2的情況下,Methylocella、Methylocapsa和Methylocystis屬的一些菌種的生長得到了促進,這表明這些菌種最適于偏酸性的培養條件[18,31]。
溫度是影響甲烷氧化菌積累PHA的另一個關鍵參數。甲烷氧化菌通常在常溫條件下表現出較好的PHA積累能力。在較高溫度(30,37 ℃)時,PHA的積累量最多可增加30%,且Methylocystis始終為優勢菌屬[37]。此外,在反應器55 ℃和58 ℃培養嗜熱型甲烷氧化菌時發現將溫度提高到58 ℃時PHA積累率未受顯著影響。這表明甲烷氧化菌具有在高溫條件下利用甲烷生產PHA的潛力[38]。因此,可以考慮研究在較廣泛的溫度范圍下甲烷氧化菌大規模合成PHA的情況。
4 數學模型在甲烷氧化菌生物合成PHA方面的應用
數學模型的建立可以評估甲烷氧化菌利用甲烷生產PHA的經濟性。通過數學模型,我們可以獲得較高的比生長速率、較高的底物利用率和較高水平的PHB生產,從而獲取最大化收益。O2和氮源的變化會對MethylosinustrichosporiumOB3b和MethylocystisparvusOBBP的生長和PHB的積累產生影響,我們可以通過建立化學計量學和動力學模型對其進行描述,從而為生物反應器設計、大規模生產和生命周期評估(LCA)提供更有價值的信息[15]。通過批量實驗數據建立Methylocystishirsuta的化學計量學和動力學的模型,所獲得的模型和參數不僅有利于生物反應器的設計和操作,而且還能夠從不同環境中篩選出Ⅱ型甲烷氧化菌進行具體研究[39]。建立MethylocystisparvusOBBP的基因組規模代謝模型(GSMM)與PHB和甲烷共同消耗的模型,并將其與Methylocystishirsuta、Methylocystissp.SC2的模型進行比較,可闡明了MMO的甲烷氧化機理,準確預測生物量的產量和耗氧率,并揭示PHB降解的回補作用[40]。
5 利用甲烷氧化菌如何高效合成PHA
5.1 生物反應器的設計
目前,進行甲烷氧化菌生物合成PHA的研究大部分是小試實驗。雖然其中的關鍵參數已經得到了優化,但仍然無法用于實際應用中。與小試實驗相比,生物反應器或生物過濾器是可對大多數工藝參數進行良好控制的培養系統。已有一些研究對同時降解甲烷和生產PHB的生物反應器進行了設計。在序批式反應器(SBR)和鼓泡塔生物反應器(BCB)中Methylocystis分別達到20%~25%和40%~50%的積累量[16,41]。BCB中獲得的較高PHB含量可能是由于安裝了內部氣體再循環系統,與SBR相比該系統可達到更高的甲烷傳質。對生物反應器的運行參數進行優化,可在最佳條件下獲得最大的PHA積累量。使用全因子設計對BCB合成PHB進行優化,發現在甲烷:空氣比例為50∶50、初始氮源為50時,PHB積累量最大為細胞干重的42.5% (w/w)[16]。對BCB的5個工藝變量(氮源含量、磷酸氫二鈉、甲烷與空氣的比例、種齡和pH)進行優化,在最佳操作條件和工程參數下,干燥生物質中的PHB含量達到25%(w/w)[42]。于BCB中高細胞密度培養Methylocystishirsuta,在最佳條件下PHB含量可到達73.4%(w/w)[19]。
另一方面,大多數由Ⅱ型甲烷氧化菌生產PHA的實驗主要集中于使用純甲烷,很少將沼氣或天然氣等原料直接用作甲烷的來源合成PHA。研究發現從合成沼氣中將甲烷和CO2生物轉化為PHB和脂質,MethylocystisparvusOBBP可獲得最大PHB含量約為30%[33]。利用Methylocystishirsuta進一步評估了沼氣作為甲烷來源進行PHA積累的可行性,獲得的PHB含量約為45%[43]。研究還證實了Methylocystis為優勢菌的混合菌群利用含有痕量乙烷、丙烷和少量丁烷的管道天然氣生產PHB(高達42%)的可行性[44]。可先通過合成沼氣對甲烷氧化菌利用其合成PHA進行研究與優化,隨后直接使用沼氣或天然氣等原料合成PHA,并就原料中其余成分對甲烷氧化菌生長和PHA積累的影響進行探索。
5.2 共底物的添加
不同的碳源作為共底物會對甲烷氧化菌生長性能和PHA積累量產生不同的影響。在Methylocystishirsuta的培養過程中以VFA的形式添加額外碳源,研究發現添加VFA不會妨礙該菌株的生長,且最大PHA產量和最終PHA含量分別增加至0.45~0.63 g PHA/g 底物和48%~54%[43]。添加甲醇可以提高PHA的積累量和分子量。研究不同碳源對Methylocystishirsuta合成PHB的影響,結果表明該菌株具有在各種底物上生長的能力,且使用1∶1 的甲醇∶乙醇時PHB含量最大可達到85% (w/w)[19]。適量的甲醇與甲烷共同添加可以提高MethylosinustrichosporiumIMW3011中積累的PHB的分子量[45]。添加甲酸可以作為替代的電子供體,有助于在甲烷缺乏的條件下維持PHA的積累水平并延緩PHA的消耗[17]。因此,可以通過提供不同濃度的不同碳源來優化甲烷氧化菌的PHA積累能力。
不同的碳源作為共底物會對PHA的種類產生影響。目前PHB是生產最多的PHA,但工業規模的廣泛應用要求生物聚合物具有多樣性。在過去的研究中,已發現可以通過在甲烷氧化菌培養物中添加共底物的措施來合成特定生物聚合物。添加戊酸、丙酸、戊醇等可以實現PHBV的合成,但其中的3-羥基戊酸酯(HV)含量有所不同。通過在培養液中添加戊酸使Methylocystishirsuta、MethylocystisparvusOBBP和MethylosinustrichosporiumOB3b菌株產生PHBV,HV的含量(20%~40%)取決于共底物的濃度[43,46-47]。丙酸和正戊醇也已成功地用于Methylocystis的PHBV生產中,但HV含量遠低于戊酸作為共底物時的含量[47-48]。通過調節戊酸濃度和限制Cu可以增加Methylocystissp.WRRC1中的PHA和特定HV含量,最大PHA和HV含量分別為78%和>50%[48]。通過添加檸檬酸和丙酸,MethylobacteriumorganophilumCZ-2生成的聚合物包含6種不同的單體:3HB、3HV、4HV、4HH、3HO和4HO,表明了該菌合成PHA的多功能性[49]。最近研究發現使用ω-羥基鏈烷酸酯可由MethylocystisparvusOBBP生產新的特定共聚物,例如聚(3-羥基丁酸酯-co-4-羥基丁酸酯)(P3HB4HB)、聚(3-羥基丁酸酯-co-5-羥基戊酸酯-co-3-羥基戊酸酯)(P(3HB-co-5HV-co-3HV))或聚(3-羥基丁酸酯-co-6-羥基己酸酯-co-4-羥基丁酸酯)(P(3HB-co-6HH-co-4HB)),其表現出明顯增強的機械性能,分子量4.5×105~1.5×106Da[50]。 雖然這方面的研究已經取得了上述成果,但未來的研究仍應繼續評估新的共底物對甲烷氧化菌生長和PHA生產的影響。
6 結論與展望
利用甲烷氧化菌將甲烷氧化合成PHA既可以有效地減少甲烷的排放,又可以合成具有優良特性、可廣泛應用的生物聚合物PHA。從目前的研究來看,甲烷氧化菌將甲烷作為電子供體可獲得一定量的PHA積累。許多因素會對甲烷氧化菌的PHA積累能力產生影響,但影響效果會因菌種的不同而發生變化。大多數研究報道仍在針對小試實驗進行優化改進,對更大的生物反應器系統性能和運行狀況研究較少。因此,盡管甲烷氧化菌利用甲烷合成PHA具有較大潛力,但進一步研究仍需做到:(1)開發新型生物反應器配制和運行策略,以同時增加甲烷的去除率和生物量生產率;(2)在營養限制條件下,通過添加共底物來合成特定生物聚合物,以優化PHA的生產;(3)了解不同環境因素產生作用的機理和動力學,建立模型進一步優化生產條件,以獲得PHA積累最大化。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
科技傳播(2019年22期)2020-01-14 03:06:54
