硼對根瘤菌胞外多糖和吲哚乙酸分泌的調控研究
2021-05-21 05:22:00陳永崗康文娟吳芳阿蕓師尚禮張翠梅李自立
草業學報 2021年5期
關鍵詞:差異
陳永崗,康文娟,吳芳,阿蕓,師尚禮,張翠梅,李自立
(甘肅農業大學草業學院,甘肅 蘭州 730070)
根瘤菌能與豆科植物共生結瘤固氮,將大氣中分子態氮轉化為氨供宿主植物吸收利用,進而促進宿主植物生長發育[1]。現代農業生產中,根瘤菌拌種提高豆科植物生產力已經成為一種常見的農業措施[2]。然而由于宿主種類、根瘤菌種以及其他因素的影響,使根瘤菌結瘤能力千差萬別,部分根瘤菌難以高效地與宿主植物結瘤固氮,達到應有的增產效果[3]。因此如何提高根瘤菌接種效率,一直是豆科植物固氮體系構建研究的熱點。
研究發現,根瘤菌可以內生于植物體內,并運移至種子器官定殖,隨種子的萌發而優先共生結瘤[4]。但根瘤菌在侵染宿主過程中,會誘發植物的防御反應,減弱根瘤菌的侵染能力[5]。根瘤菌侵染宿主并侵入體內的過程中,根瘤菌分泌的次生代謝產物胞外多糖和吲哚乙酸(indole-3-acetic acid,IAA)扮演著重要角色,它們不僅可以減弱宿主的防御能力,促進根瘤菌的侵染及侵入[5?6],還可以促進根瘤菌在植物根皮層細胞內運移及根與根瘤菌的有效互作[7]。據報道,硼是根瘤菌固氮所必需的微量元素[8],其在根瘤菌與宿主信號傳遞、根瘤細胞壁和膜結構的形成等過程中發揮著重要作用[9],它不僅對宿主植物生長有影響,還對根瘤菌侵染宿主以及侵染成功后在宿主體內運移定殖有重要作用[10]。苗陽陽等[11]發現,添加外源硼可以促進根瘤菌侵染宿主以及侵入后在宿主體內運移定殖的數量。缺硼會減弱根瘤菌定殖能力、減少根瘤菌定殖數量并削弱固氮能力,同時也會降低根瘤菌表面多糖的形成,進而影響其功能的發揮[12]。硼還可以促進宿主植株及根瘤菌IAA 合成,有利于其在植物組織內擴散和運輸[13?14],而IAA 能增大宿主植物細胞壁膜透性,促進養分釋放及根系根瘤菌侵染,同時可以降低宿主植物防衛系統胞壁降解酶活性,使入侵根瘤菌易定殖于植物組織內[5,15]。因此,闡明硼對根瘤菌分泌胞外多糖及IAA 的調控機制,對于研究硼促進根瘤菌侵染宿主的機制,提高根瘤菌接種效率具有指導意義。
1 材料與方法
1.1 試驗材料
根瘤菌:熒光標記根瘤菌[Ensifer meliloti,gn5f,分離自甘農 5 號紫花苜蓿(Medicago sativa)種子]。硼酸:購自無錫市晶科化工有限公司,含量大于99.5%。YMA 液體培養基:準確稱取10 g 甘露醇,0.5 g 磷酸氫二鉀,0.2 g 七水硫酸鎂,0.1 g 氯化鈉,1 g 酵母粉,并調節 pH 為 6.8~7.0,定容至 1 L。YMA 固體培養基加瓊脂:16~20 g?L?1。
1.2 最適硼濃度確定
設置硼濃度梯度為 0(CK)、0.05、1、5、10 和 100 mg?L?1,然后按濃度梯度加至 40 mL YMA 液體培養基,每個處理3 次重復,以上操作均在無菌操作臺內進行;將活化后的根瘤菌gn5f 接種于50 mL YMA 液體培養基,28 ℃、180 r?min?1培養至OD600值為0.5~0.8。取5 mL 該菌液加入上述含有不同硼濃度的40 mL YMA 液體培養基中,再在 28 ℃、180 r?min?1培養至 OD600值為 0.5~0.8,在不同時間段測定 OD600值,確定根瘤菌 gn5f 最適硼濃度。
1.3 硼處理根瘤菌gn5f 胞外多糖和IAA 的測定
1.3.1 胞外多糖的測定 采用苯酚?硫酸法測定胞外多糖[16]。步驟如下:配制上述所選最適硼濃度,添加至40 mL YMA 液體培養基中,對照不加硼液,每個處理3 個重復。將已培養至OD600值為0.5~0.8 的根瘤菌gn5f 取5 mL 加入 40 mL 液體培養基中,在 28 ℃、180 r?min?1培養 72 h 后,在 4 ℃下,15000 r?min?1離心 10 min,棄沉淀,收集上清液;將上清液轉入新的離心管,加入Sevag 試劑(氯仿?正丁醇體積比為5∶1,樣液與試劑體積比為2∶1),振蕩 30 min,然后 6000 r?min?1,離心 10 min,收集上清液,反復處理 4 次;將上清液移到另一 50 mL 離心管中,加入 3倍體積 95% 乙醇,4 ℃冰箱中浸提 12 h,4 ℃,15000 r·min?1,離心 45 min,并收集沉淀,棄上清液;在超凈工作臺內風干,稱重,即得粗胞外多糖含量。測定粗胞外多糖吸光度值,代入曲線方程:y=54.4858x?0.01024,R2=0.9951,計算根瘤菌gn5f 胞外多糖含量。
1.3.2 吲哚乙酸(IAA)的測定 采用高效液相色譜(high performance liquid chromatography,HPLC)測定IAA[17]。步驟如下:根瘤菌培養方法同上,培養 72 h 后,在 4 ℃下,15000 r·min?1,離心 10 min,棄沉淀,收集上清液;將上清液低溫干燥至 10~15 mL,并用鹽酸(1 mol?L?1)將 pH 值調至 2.8;轉入分液漏斗,加入 20 mL 乙酸乙酯,振蕩5 min,待分層后;轉移上層溶液于另一分液漏斗中,向下層溶液再加入乙酸乙酯提取一次,合并上層溶液于50 mL 燒瓶中,于低溫干燥濃縮至近干;用2 mL 無水乙醇溶解后盛于小試管中。于高效液相色譜儀測定IAA含量,測定條件為:進樣量 10 uL,流動相∶甲醇∶水∶乙酸(45∶54∶1),流速0.8 mL?min?1,Eclipse Plus C18色譜柱(4.6 mm×250 mm,美國Thermo fisher 公司)。
1.4 硼處理根瘤菌gn5f 蛋白質測定及純化
根據1.3.1 的方法收集最適硼處理后的根瘤菌gn5f 的菌體,低溫保存備用;冷凍狀態下取出樣品,轉移至MP振蕩管中,加入適量抽提 buffer(1% SDS,200 mmol?L?1DTT,50 mmol?L?1Tris?HCl,pH 8.8 含蛋白酶抑制劑),渦旋混勻并用高通量組織研磨儀振蕩3 次,每次40 s,裂解30 min。100 ℃下孵化10 min,置冰上冷卻。在4 ℃ 12000 r·min?1條件下,離心 20 min,取上清,加入預冷丙酮(1∶4),在?20 ℃下過夜沉淀。次日在 4 ℃、12000 r·min?1條件下,離心20 min,棄上清,向沉淀中加入90%預冷丙酮,混勻后離心棄上清,重復2 次。向沉淀中加入蛋白裂解液(8 mol?L?1尿素,含蛋白酶抑制劑),在 4 ℃ 12000 r·min?1條件下,離心 20 min,棄沉淀,留上清[18]。然后根據BCA 試劑盒(蘇州達麥迪生物醫學科技有限公司)操作測定蛋白質濃度,并進行SDS?PAGE 電泳[19];取樣品各100 μg,向各樣品中加入終濃度為100 mmol·L?1三乙基碳酸氫銨緩沖液(tetraethylammonium bromide,TEAB)和 10 mmol·L?1TCEP[tris(2-carboxyethyl)phosphine],在 37 ℃下反應 60 min。加入終濃度 40 mmol?L?1碘乙酰胺,室溫避光反應 40 min。加入 7 倍體積 100 mmol?L?1TEAB 和胰蛋白酶,37 ℃酶解過夜。酶切后用HLB(hydrophile lipophile balance)、陽離子固相萃取進行脫鹽純化。
1.5 液相串聯質譜分析
使用 Q-Exactive HF-X 質譜儀(Thermo fisher 公司,美國)鑒定蛋白質序列,掃描范圍為 350~1300 m?z?1,采集模式為DDA(discontinous deformational analysis);一級質譜分辨率為70000,最大注入時間為20 ms,AGC tar?get 為3e6,采用高能碰撞解離(high energy collision dissociation,HCD)碎裂方式;選擇母離子中信號最強的20 個進行二級碎裂,二級分辨率 17500,AGC target 為 1e5,最大注入時間 50 ms,起始質荷比:100 m?z?1;最小 AGC tar?get 8e3,臨界強度為1.6e5,動態排除時間18 s。液相條件為:采用C18 column 色譜柱(Thermo fisher 公司,美國),A 液:0.1%甲酸和 2% 乙腈,B 液:0.1%甲酸和 80% 乙腈,流速為 300 nL?min?1,分離時間為90 min。
1.6 數據處理
用SPSS 19.0 和Excel 2010 軟件進行數據分析處理。用PEAKS Studio 8.5 進行質譜數據搜索,選擇已經建立好的數據庫。將搜庫得到的蛋白質數據重新分析,在蛋白質層面進行t配對檢驗,計算相應的P-value 及差異倍數(fold change,FC),以此作為顯著性指標。根據FC 以及P-value 進行差異蛋白質篩選。當FC≥1.5 且P-val?ue<0.05 時,表示該蛋白質表達量顯著上調;當FC≤1 且P-value<0.05 時,表示該蛋白表達量顯著下調。并對差異蛋白進行GO(gene ontology)功能注釋以KEGG(kyoto encyclopedia of genes and genomes)通路富集分析。
2 結果與分析
2.1 適宜硼濃度的篩選
在 20~180 min 時,適宜的硼濃度可以促進根瘤菌 gn5f 的生長。20 min 時,硼濃度為 100 mg?L?1,根瘤菌生長速度最快,顯著大于對照(P<0.05),其余處理與對照差異不顯著(P>0.05);60 和140 min 時,當硼濃度小于10 mg?L?1,根瘤菌gn5f 生長速度與對照相比差異均不顯著(P>0.05),其余處理生長速度較對照均差異顯著(P<0.05),且硼濃度為 100 mg?L?1時生長速度最快;100 min 時,硼濃度為 100 mg?L?1生長速度最快,除添加 1 mg?L?1硼生長速度較對照差異不顯著(P>0.05)外,其余處理較對照均差異顯著(P<0.05);180 min 時,除100 mg?L?1硼濃度處理較對照差異顯著(P<0.05)外,其余處理較對照差異均不顯著(P>0.05)(圖1)。
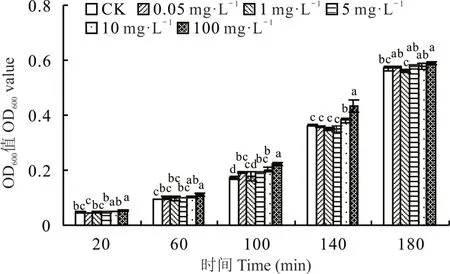
圖1 不同硼濃度對根瘤菌gn5f OD600的影響Fig.1 Effects of different boron concentrations on gn5f OD600
結合前期試驗結果[11](100 mg?L?1硼處理根瘤菌gn5f 后接種紫花苜蓿時,根瘤菌gn5f 在紫花苜蓿體內運移數量最多),確定根瘤菌gn5f 的最適硼濃度為100 mg?L?1。
2.2 根瘤菌gn5f 的生長曲線
對照和添加最適硼時的生長曲線均呈現S 型,分為延遲期、對數生長期、穩定期和衰亡期。在根瘤菌的對數期,細胞數量呈幾何級數增長,細胞內各種酶的合成也最活躍、代謝旺盛,各種物質成分和生理特性較為一致,結合圖1 和圖2 確定后續蛋白組學取樣時間為180 min,即對數期時進行菌體收集,以進行后續蛋白組學的研究。添加硼與對照相比,達到相同OD600值的培養時間以及進入對數期的培養時間均明顯縮短。說明最適硼處理顯著促進了根瘤菌生長。由于胞外多糖以及IAA 是根瘤菌的次生代謝產物,根據細菌的特性,次生代謝產物分泌主要在穩定期,結合最適硼處理下的生長曲線,確定后續試驗最適硼濃度處理下,根瘤菌gn5f 培養時間為72 h(圖2)。
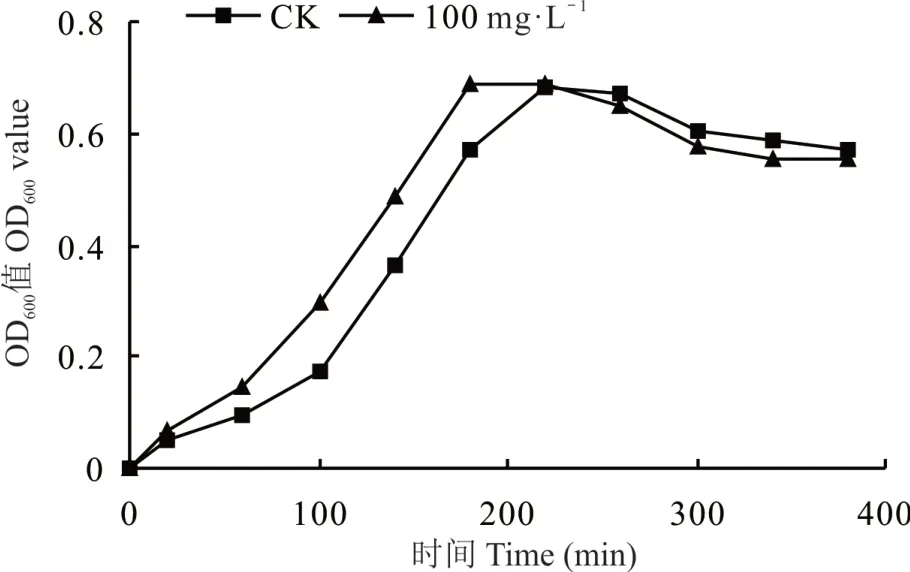
圖2 硼處理前后根瘤菌gn5f 生長曲線Fig. 2 Growth curve of strain gn5f treated by boron before and after
2.3 硼對根瘤菌gn5f 產胞外多糖和IAA 的影響
相比對照,100 mg?L?1硼處理根瘤菌 gn5f 顯著提高了其胞外多糖和IAA 產量(P<0.05),其中胞外多糖提高33.0%,IAA 提高65.9%。表明,最適硼處理根瘤菌gn5f 可顯著提高胞外多糖和IAA 含量(圖3)。
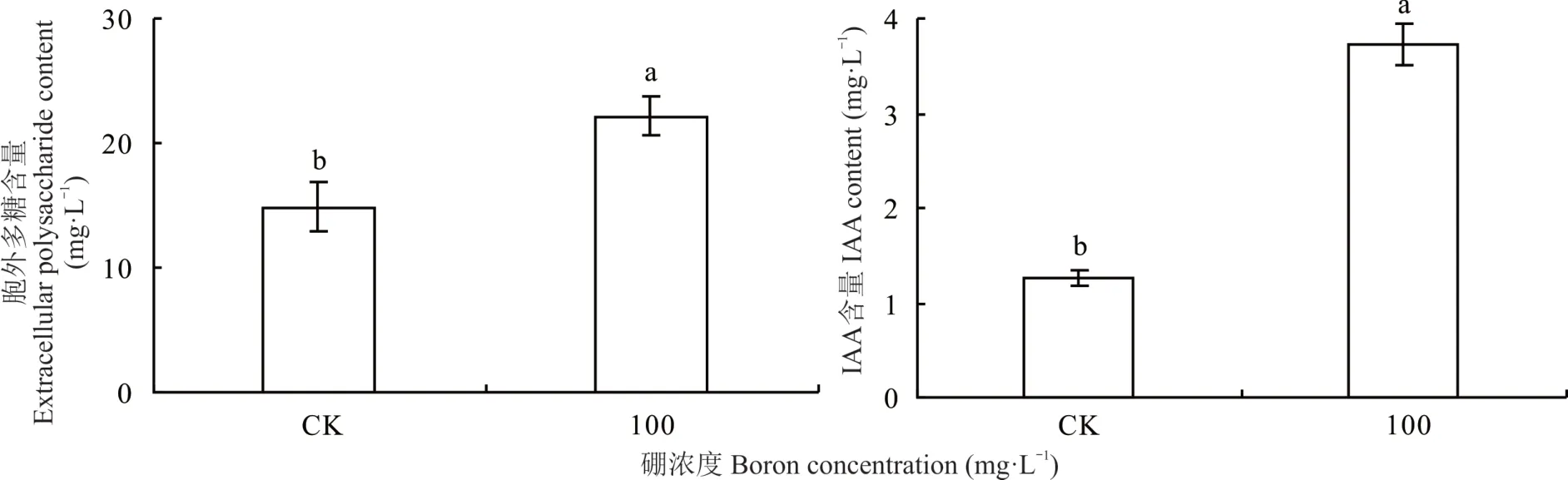
圖3 硼對根瘤菌gn5f 產胞外多糖和IAA 的影響Fig.3 Effect of boron on extracellular polysaccharides and IAA production of gn5f
2.4 硼處理根瘤菌gn5f 蛋白質SDS?PAGE 電泳
各樣本電泳條帶均清晰,樣本間平行性較好。蛋白預測質譜顯示酶解正常,色譜質譜行為正常,可以進行后續試驗(圖4)。
圖4 SDS-PAGE 電泳結果Fig. 4 SDS-PAGE electrophoresis results
2.5 硼處理根瘤菌gn5f 差異蛋白質生物信息學分析
采用 Label free 技術,對 100 mg?L?1硼處理根瘤菌gn5f 進行定量蛋白組學分析,共鑒定出160 個蛋白點,54 個差異表達蛋白,其中7 個蛋白表達上調,47 個蛋白表達下調。表明最適硼濃度處理根瘤菌,可引起部分蛋白表達上調和下調。
2.5.1 硼處理根瘤菌gn5f 差異蛋白GO 富集分析
根據分子功能(molecular function,MF)、細胞組分(cellular component,CC)、生物過程(biological pro?cess,BP)對差異表達蛋白進行GO 功能注釋,統計對應差異蛋白所占的百分比(圖5)。結果表明,100 mg?L?1硼處理根瘤菌gn5f 后,差異蛋白參與的分子功能主要包括催化活性(36 個),結合(20 個)及轉運活性(7個)等;而參與的細胞組分主要有細胞組分(11 個),生物膜(5 個)及蛋白質復合物(5 個)等;主要參與的生物過程為代謝過程(30 個),細胞過程(23 個)及定位(7個)等。結果表明,100 mg?L?1硼處理根瘤菌 gn5f 后差異表達蛋白具有多種分子功能,并參與多個生物過程(圖5)。
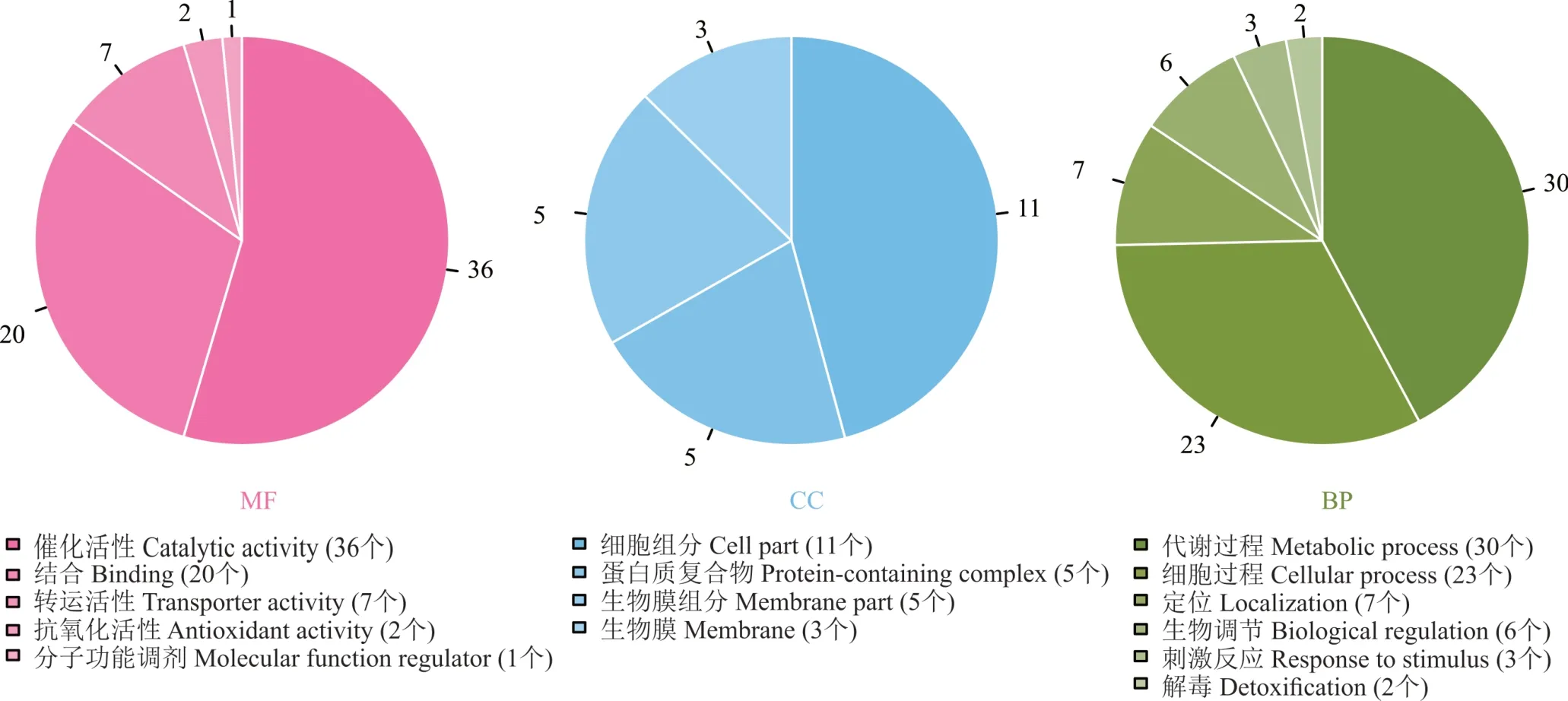
圖5 硼處理根瘤菌gn5f 差異表達蛋白GO 功能富集分析Fig.5 Functional enrichment analysis of boron-treated rhizobium gn5f differentially expressed protein
2.5.2 硼處理根瘤菌gn5f 差異蛋白KEGG pathway 富集分析 對差異蛋白進行KEGG pathway 富集分析,發現差異蛋白分布于4 個功能區,有28 個蛋白參與代謝(全局和概覽通路),7 個參與環境信息處理(膜運輸),6 個參與細胞過程(運輸和分解代謝)、3 個參與遺傳信息處理(轉錄)及13 個未知功能(圖6)。已知功能的差異蛋白主要富集在抗生素合成、碳代謝、賴氨酸降解、丁酸脂代謝、酪氨酸代謝(tyrosine metabolism)及色氨酸代謝(trypto?phan metabolism)等代謝通路中(圖7)。其中已知功能的上調蛋白乙酰輔酶A 合成酶(A0A4R3PRU1)共參與11條代謝通路,琥珀酸半醛脫氫酶(NADP+)(A0A1Q9AU32)和煙酰胺腺嘌呤二核苷酸(nicotibnamide adenine di?nucleotide,NAD)依賴性琥珀酸半醛脫氫酶(A0A0Q6ZU08)各參與7 條代謝通路,且共同參與γ-氨基丁酸(γ-aminobutyric acid,GABA)代謝、丙氨酸代謝及谷氨酸代謝等與三羧酸循環有關的代謝通路。4 種上調蛋白未找到KEGG 相對應通路。下調蛋白乙酰輔酶 A 乙酰轉移酶(A0A0N0JNE5)、烯酰輔酶 A 水合酶(A0A2M8UJS2)、脂酰輔酶 A 脫氫酶(J2L150)、煙酰胺腺嘌呤二核苷酸(NAD)依賴性琥珀酸半醛脫氫酶(A0A0Q7XRQ0)、磷酸烯醇丙酮酸合酶(A0A4Q8YPJ5)、色氨酸合酶 β 鏈(A0A2T0P7K1)等共同參與抗生素合成、碳代謝及次生代謝產物生物合成等通路(圖7)。
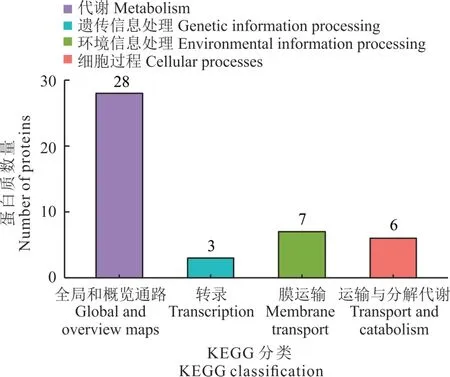
圖6 硼處理根瘤菌gn5f 差異表達蛋白KEGG 注釋統計Fig. 6 KEGG annotation statistics of gn5f differentially ex?pressed protein of boron-treated rhizobium
圖7 根瘤菌gn5f 差異表達蛋白數量最多的前20 個通路Fig. 7 Top 20 pathways with the most differentially ex?pressed proteins from rhizobia gn5f
綜上,硼處理根瘤菌gn5f 會引起多種蛋白表達上調和下調,且多個蛋白協同調節多個代謝通路,繼而促進根瘤菌gn5f 生長以及其他生理代謝過程。
3 討論
3.1 根瘤菌gn5f 適宜硼濃度篩選以及生長曲線的測定
OD600值表示細菌吸收的光密度值,用來反映細菌相對生長量[20]。細菌的生長主要是指細胞數目的增多以及胞內物質的倍增[21],本研究發現,與對照相比,硼濃度為 100 mg?L?1時,根瘤菌 gn5f OD600值增加最顯著,說明適宜的硼可以促進該根瘤菌生長[11]。Mon?tanari 等[22]發現,細菌在受到環境脅迫時,會誘發體內生理應激反應,包括蛋白質復合物的重組以及代謝物質變化,以此來適應微生態環境的變化,本研究表明,硼脅迫誘發了根瘤菌的響應機制,使體內某些代謝途徑或酶發生變化,繼而使生長加快,對數期縮短。根瘤菌的繁殖和個體的生長依賴于胞內物質的合成和加倍[23],研究發現原核生物體內的類鈣調蛋白與細胞分裂以及細胞內多種酶的活性有關[24],硼可能是通過激活該蛋白,促進細胞分裂以及與生長相關酶的活性,間接促進根瘤菌的繁殖和生長。
3.2 硼對根瘤菌gn5f 產胞外多糖和IAA 的影響
硼是固氮細菌固氮所必需的微量元素,在根瘤菌與宿主共生關系建立、表面多糖形成以及根瘤菌固氮等過程中發揮著重要作用[6,8]。缺硼可以減弱根瘤菌定殖能力,影響根瘤菌表面多糖形成及功能發揮[11,25],同時減少IAA 含量和根瘤菌侵染宿主時的侵染位點[26]。本研究發現,最適硼可以顯著促進根瘤菌gn5f 胞外多糖和IAA 含量,Martin 等[27]也發現,適宜的硼酸會影響白鏈球菌碳水化合物的能量代謝,且可以導致根瘤菌Botrytiscinerea胞質內溶物:可溶性糖的泄露[28],同時硼還可以影響多種酶的活性,其中大多數與能量底物代謝有關[29]。因此,推測硼對根瘤菌胞外多糖和IAA 產量的影響,可能主要是通過影響碳水化合物代謝過程及各種關鍵酶活性,繼而影響其合成以及分泌。
3.3 硼對根瘤菌gn5f 差異蛋白組學的影響
硼是固氮細菌固氮所必需的微量元素,在根瘤菌與宿主共生關系建立,促進根瘤菌侵染宿主以及促進根瘤菌次生產物胞外多糖和IAA 合成等過程中扮演著重要作用,硼處理根瘤菌接種苜蓿后,根瘤菌對苜蓿的影響已有研究[11]。本研究發現,硼處理下根瘤菌gn5f 蛋白組學發生變化,其中一些重要的參與γ-氨基丁酸(GABA)代謝、丙氨酸代謝,天冬氨酸和谷氨酸代謝的蛋白表達上調,包括乙酰輔酶A 合成酶、煙酰胺腺嘌呤二核苷酸(NAD)依賴性琥珀酸半醛脫氫酶及琥珀酸半醛脫氫酶(NADP+)。乙酰輔酶A 合成酶與生物體內物質和能量的生成有關,主要參與能量代謝及物質合成相關的代謝通路[30]。丙氨酸可以轉化為丙酮酸,丙酮酸可以進入三羧酸循環(tri?carboxylic acid cycle,TCA)。天冬氨酸和谷氨酸在特定條件下可以相互轉換,谷氨酸主要參與γ-氨基丁酸、蛋白質以及葡萄糖等的合成;γ-氨基丁酸與丙酮酸在特定條件下可以生成琥珀酸半醛和丙氨酸,琥珀酸半醛在琥珀酸半醛脫氫酶(NADP+)的作用下生成琥珀酸,進入三羧酸循環,該過程稱為γ-氨基丁酸(GABA)旁路[31]。有研究發現,在許多細菌體內存在兩種類型的琥珀酸半醛脫氫酶,其活性分別依賴于煙酰胺腺嘌呤二核苷酸磷酸(nico?tinamide adenine dinucleotide phosphate,NADP)或煙酰胺腺嘌呤二核苷酸(NAD)[32]。表明硼處理根瘤菌 gn5f 后煙酰胺腺嘌呤二核苷酸(NAD)依賴性琥珀酸半醛脫氫酶、乙酰輔酶A 合成酶、琥珀酸半醛脫氫酶(NADP+)蛋白表達上調,琥珀酸半醛脫氫酶(NADP+)Gab D(W6S1U0)蛋白表達下調均與上述代謝通路有關。上述代謝過程可以為根瘤菌生長及胞外多糖和IAA 的合成提供各種前體物質和能量。
烯酰輔酶A 水合酶可以催化脂肪酸β 氧化中水分子可逆地順式加成到a,β 不飽和硫酯雙鍵上[33?34],脂酰輔酶A 脫氫酶主要參與脂肪酸β 氧化,是脂肪酸β 氧化中一種重要的酶[35],本研究中烯酰輔酶A 水合酶和脂酰輔酶A脫氫酶表達均顯著下調,表明硼可以減弱或抑制根瘤菌gn5f 的脂肪酸β 氧化。
磷酸烯醇丙酮酸合酶與細菌糖異生路徑中磷酸烯醇式丙酮酸的合成有關[36]。乙酰輔酶A 乙酰轉移酶在多種酶存在條件下可以將乙酰輔酶A 轉化成多聚3-羥丁酸[Poly(3-hydroxybutyrate),PHB],而PHB 是根瘤菌的儲能物質[37]。色氨酸合酶β 鏈可以催化吲哚乙酸合成色氨酸[38]。這些蛋白表達下調,表明硼處理會抑制該根瘤菌的糖異生途徑、儲能物質合成以及色氨酸的分解。
可見,100 mg?L?1硼處理根瘤菌gn5f,會引起與三羧酸代謝有關的蛋白表達上調,進而促進三羧酸循環,同時會引起與糖異生、脂肪酸β 氧化以及氨基酸代謝有關的蛋白表達下調,進而抑制這些代謝通路。表明硼對根瘤菌gn5f 胞外多糖及IAA 的調控是一個復雜的反應過程,是多個蛋白協同作用的結果。
3.4 硼對根瘤菌gn5f 產胞外多糖和IAA 的調控探究
根據100 mg?L?1硼處理根瘤菌gn5f 后上調差異蛋白主要富集的代謝通路以及已知根瘤菌胞外多糖合成的調控[39],推測硼可能是通過促進GABA 旁路代謝、丙酮酸代謝以及谷氨酸代謝等與三羧酸循環有關的代謝通路,為胞外多糖合成提供能量以及所需要的各種糖類物質(D-半乳糖殘基,D-葡萄糖殘基以及D-葡萄糖醛酸等),繼而促進胞外多糖的合成。具體機制有待進一步研究證明。
100 mg?L?1硼處理根瘤菌gn5f 后,上調表達蛋白促進了色氨酸代謝通路,推測硼促進色氨酸合成,然后色氨酸經吲哚-3-乙酰胺(indole-3-acetamide,IAM)或吲哚-3-丙酮酸途徑合成吲哚乙酸(IAA),繼而促進IAA 合成。
4 結論
篩選的根瘤菌 gn5f 最適硼濃度為 100 mg?L?1;100 mg?L?1硼可顯著促進根瘤菌 gn5f 胞外多糖和 IAA 的合成;最適硼處理根瘤菌gn5f 后共鑒定到54 個差異表達蛋白,7 個上調蛋白,47 個下調蛋白。推測硼可能是通過促進丙酮酸代謝、GABA 旁路代謝,谷氨酸代謝等代謝通路,為胞外多糖合成提供能量以及所需要的各種單糖(D-半乳糖殘基,D-葡萄糖殘基以及D-葡萄糖醛酸等),繼而促進胞外多糖的合成;IAA 合成機制為:硼通過促進色氨酸的合成,然后色氨酸在相關酶的作用下經吲哚-3-丙酮酸或吲哚-3-乙酰胺(IAM)途徑合成吲哚乙酸(IAA),繼而促進IAA 合成。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:19:16
汽車實用技術(2022年10期)2022-06-09 11:16:58
音樂探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
中國非營利評論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現代語文(2016年21期)2016-05-25 13:13:50
