家蠶絲氨酸蛋白酶基因BmSPH33對BmNPV誘導的免疫響應
2021-05-25 08:52:12張永紅邵榆嵐蘇振國白興榮
西南農業學報 2021年1期
張永紅,邵榆嵐,蘇振國,白興榮
(云南省農業科學院蠶桑蜜蜂研究所,云南 蒙自 661101)
【研究意義】絲氨酸蛋白酶(Serine protease,SP)是一類以絲氨酸為催化中心的蛋白質水解酶基因家族,在昆蟲體內參與了食物消化、發育、先天免疫等[1-3]。家蠶絲氨酸蛋白酶基因家族包括BmSP(Bombyxmoriserine protease,BmSP)和同源體BmSPH(Bombyxmoriserine protease homolog,BmSPH)[4],它們是家蠶先天免疫通路中調控級聯的關鍵酶類[5],所以研究BmSPs基因在家蠶免疫應答方面的作用具有重要意義。【前人研究進展】絲氨酸蛋白酶(SPs)普遍存在于生物體中,是一類含有保守催化三聯殘基His、Asp和Ser的蛋白質水解酶[6],它們通常以非酶催化活性的前體存在,其前肽必須被修剪才能激活,趙萍等[7]在家蠶基因組中共鑒定到143個BmSPs基因,分別標記為BmSP1—BmSP143,其中包括51個BmSPs和94個BmSPHs。BmSPs基因在家蠶體內組織和時期表達量不同,進而發揮著不同的功能[8]。已有研究發現BmSPs基因家族成員中BmSP2作為中腸消化酶,對家蠶核型多角體病毒(Bombyxmorinucleopolyhedrovirus,BmNPV)具有免疫作用[9],且BmSP25基因在BmNPV的感染誘導下產生了免疫響應[10]、BmSP142在家蠶感染BmNPV與BmBDV(Bombyxmoribidensovirus,BmBDV)后均表現為上調[11],BmSP36與BmSP141參與家蠶的食物消化過程[12-13]、BmSP95在家蠶蛹皮蛻化過程中發揮著功能[14]。絲氨酸蛋白酶同源物(SPHs)在氨基酸序列上與SPs相似,但由于一個或多個催化殘基發生突變而缺乏酶催化活性[15],但它們參與了前酚氧化酶(proPO)激活系統,導致黑色素化。研究發現家蠶絲氨酸蛋白酶同源體BmSPH-1具有幼蟲-蛹轉化、蛹表皮黑色素化和家蠶天然免疫的雙重作用[16]。【本研究切入點】家蠶絲氨酸蛋白酶BmSPs在家蠶腸道內主要參與食物消化,還參與宿主的免疫應答過程,至今未見BmSPH33基因對BmNPV免疫響應機理方面的報道。【擬解決的關鍵問題】獲得BmSPH33基因序列并結合其轉錄情況,另外家蠶感染BmNPV后該基因表達模式明確誘導響應機理,為弄清BmSPH33基因在參與家蠶先天免疫方面發揮作用提供依據。
1 材料與方法
1.1 供試材料
供試家蠶品種為大造(P50),純化后的BmNPV病毒粒子由云南省農業科學院蠶桑蜜蜂研究所保存。TaKaRa寶生物工程(大連)有限公司產品:RNAiSo Plus Total RNA提取試劑盒、One Step RNA PCR Kit (AMV)與PrimeScriptTMRT reagent kit with gDNA Eraser反轉錄試劑盒、PCR Mixture、SYBR Primix ExTaqTMII (Tli RNaseH Plus) qRT-PCR試劑盒;生工生物工程(上海)股份有限公司產品:PCR引物、瓊脂糖凝膠回收試劑盒等試劑。
1.2 試驗方法
1.2.1 家蠶組織樣品收集 家蠶大造(P50)品種幼蟲在27 ℃條件下用新鮮桑葉飼養至5齡第3天,解剖幼蟲并收集頭部、血液、體壁、中腸、脂肪體、絲腺、精巢和卵巢;BmNPV添毒實驗中,正常飼育的5齡起蠶平均分為2組,其中一組按照BmNPV多角體懸浮液5.13×107PIB·mL-1的濃度喂食家蠶10 μl·頭-1;另外一組同樣喂食10 μl ddH2O。兩組家蠶添食后常規桑葉飼養。在不同時間4、8、12、24和48 h分別解剖家蠶并收集中腸樣品,其中添食ddH2O家蠶的中腸樣品為對照。
1.2.2 BmSPH33序列分析 登錄家蠶基因組數據庫SilkwormDB,獲取BmSPH33基因ORF序列,利用生物學在線軟件對BmSPH33蛋白的分子量、等電點、信號肽、蛋白質二級結構、三級結構進行預測,其中理論等電點與分子量預測軟件ExPASy(http://web.expasy.org/)、信號肽預測軟件SignalP v5.0(http://www.cbs.dtu.dk/services/SignalP/)、功能結構域軟件CDD(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)、二級結構軟件PSIPRED(http://bioinf.cs.ucl.ac.uk/psipred/)和三級結構預測軟件SWISS-MODLE(https://swissmodel.expasy.org/);利用MEGA5.0軟件對BmSPH33與其它物種SPH33序列生成.fasta與.mega文件,結合GeneDoc軟件進行多序列比對;根據MEGA5.0 中Neighbor-joining方法構建系統發生樹[17]。
1.2.3 家蠶組織樣品總RNA提取及cDNA合成 利用液氮將家蠶不同組織樣品和不同時期處理的中腸樣品充分研磨,按照RNAiSo Plus Total RNA提取試劑盒操作步驟抽提RNA,超微量核酸蛋白測定儀檢測RNA樣品濃度和純度,將抽提的RNA根據One Step RNA PCR Kit (AMV)說明反轉錄成cDNA。
1.2.4 家蠶不同組織BmSPH33基因轉錄情況 設計BmSPH33基因RT-PCR引物(表1)。以家蠶肌動蛋白基因Bmactin3為內參基因,家蠶不同組織cDNA為模板進行PCR擴增,反應體系為25 μl:10×PCR buffer(Mg2+plus) 2.5 μl,2.5 mmol·L-1dNTPs 2 μl,5 U·μl-1r-Taq聚合酶 0.25 μl,10 μmol·L-1上下游引物 0.5 μl,cDNA模板0.5 μl,ddH2O補齊25 μl;反應條件為:94 ℃ 4 min;(94 ℃ 30 s,60 ℃ 30 s,72 ℃ 30 s)× 30 cycles;72 ℃ 10 min;12 ℃ forever,擴增產物用1 %的瓊脂糖凝膠檢測。
1.2.5 qRT-PCR檢測BmSPH33基因對BmNPV感染的響應 利用primer5.0設計BmSPH33基因熒光定量特異引物(表 1),以不同時間段ddH2O與BmNPV處理的家蠶中腸組織cDNA為模板,Bmactin3為內參基因進行qRT-PCR,參照SYBR Primix ExTaqTMII (Tli RNaseH Plus)試劑盒說明進行配制,反應體系為20 μl(SYBR Premix ExTaqII 10 μl,ROX Reference Dye 50× 0.4 μl,F/R Primers 0.4 μl,cDNA 1 μl,ddH2O 補齊20 μl),擴增程序:95 ℃30 s;(95 ℃ 30s,60 ℃ 30 s)× 40 cycles。ABI StepOnePlus儀器記錄實驗結果,不同處理的樣品設置3個重復,收集基因的Ct值數據,最后根據2-△△Ct計算BmSPH33基因的相對表達量[18]。
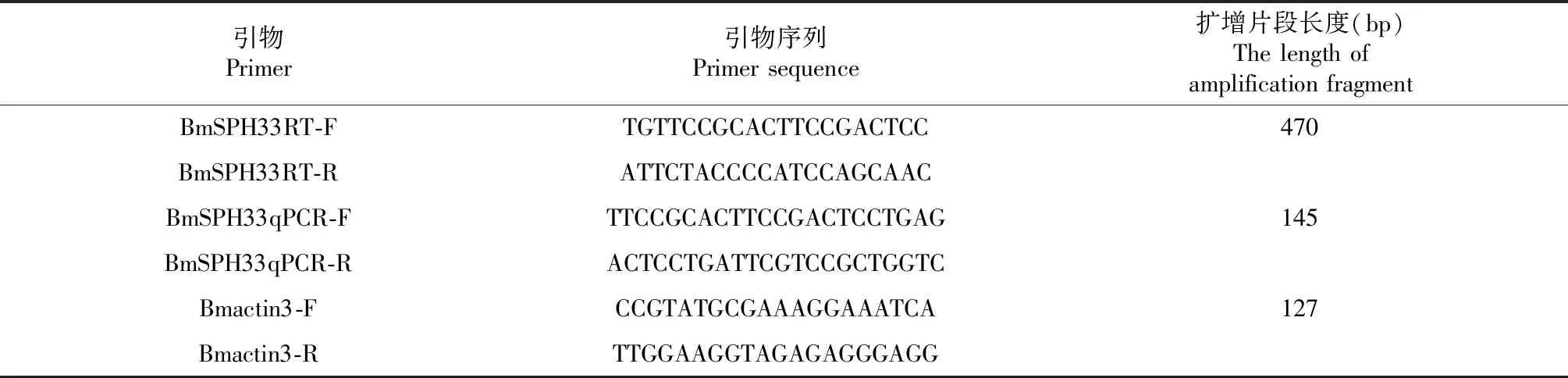
表1 試驗引物Table 1 Research primers
2 結果與分析
2.1 BmSPH33基因序列生物信息學分析
對SilkwormDB公布的BmSPH33基因ORF進行序列分析,BmSPH33ORF全長為840 bp,編碼279個氨基酸殘基,軟件預測無信號肽序列,為非分泌型蛋白,去除信號肽后理論分子量大小為22.95 kDa,等電點為5.98。NCBI CDD在線預測軟件發現,40~277位氨基酸為類胰蛋白酶絲氨酸蛋白酶結構域(Trysin-like serine protease domain,Tryp_SPc domain)。利用PSIPRED Server在線軟件預測BmSPH33蛋白質的二級結構(圖1),預測結果發現,BmSPH33蛋白由5個α螺旋、14個β折疊和一些無規則卷曲構成;利用SWISS-MODEL軟件預測三級結構(圖2),其二級結構及三級結構預測特征一致而且以β折疊構成為主。

圖1 預測的BmSPH33蛋白質二級結構Fig.1 Secondary structure for BmSPH33 protein by server
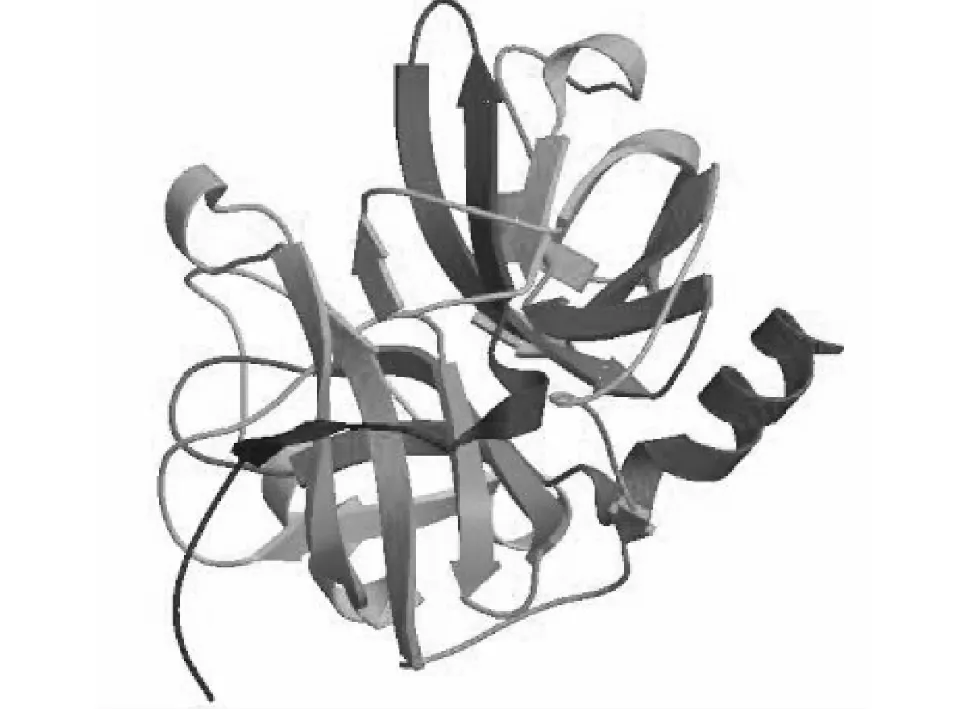
圖2 預測的BmSPH33蛋白質三級結構Fig.2 Tertiary structure of for BmSPH33 protein by server
BmSPH33的氨基酸序列(GenBank登錄號:AK378171.1)與NCBI上登錄的蓓帶夜蛾SPH33(GenBank登錄號:ACR15983.2)進行序列比對。結果顯示,各序列中均含有類胰蛋白酶絲氨酸蛋白酶結構域(圖3)。基于不同物種SPH33蛋白氨基酸序列,利用MEGA5.0軟件,采用鄰接法對構建系統發生樹(圖4),遺傳進化結果顯示,BmSPH33與蓓帶夜蛾(Mamestraconfigurata)和冬尺蠖蛾(Operophterabrumata)SPH33在親緣關系上較近,其中不同節肢動物的物種SPH33未聚在同一分支,而植物大葉杜鵑(Danausplexippusplexippus)SPH33與家蠶(Bombyxmori)、蓓帶夜蛾(Mamestraconfigurata)和冬尺蠖蛾(Operophterabrumata)遺傳距離相近。

BmSPH33:家蠶SPH33;MacoSPH33:蓓帶夜蛾SPH33BmSPH33:Bombyx mori SPH33;MacoSPH33:Mamestra configurata SPH33圖3 BmSPH33與MacoSPH33氨基酸序列比對分析Fig.3 Sequence alignment of amino acid between BmSPH33 and MacoSPH33
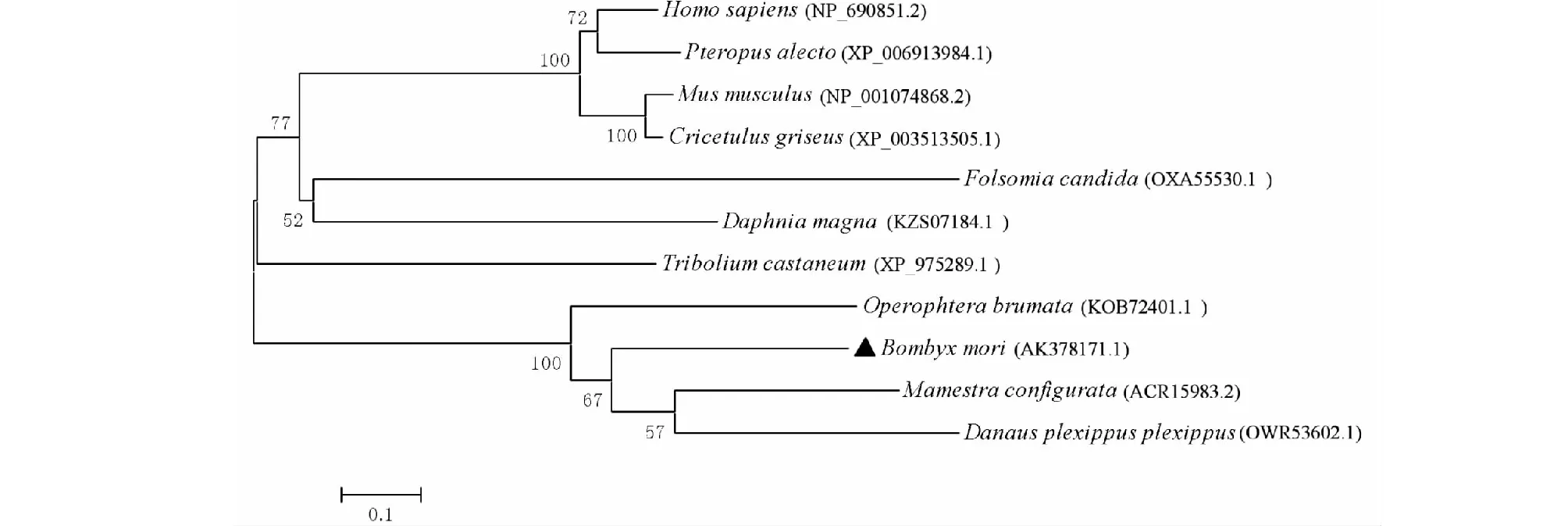
圖4 BmSPH33與其同源氨基酸序列進化分析Fig.4 Evolution analysis of amino acid between BmSPH33 with other homologues sequence
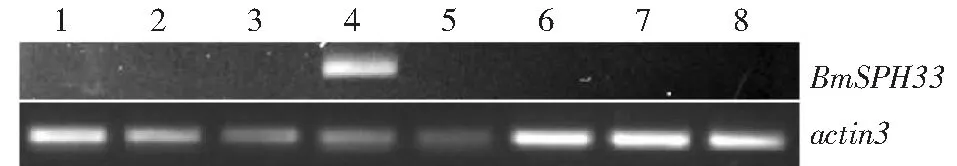
1:頭;2:血液;3:體壁;4:中腸;5:脂肪體;6:絲腺;7:精巢;8:卵巢1. Head;2: Hemocyte;3: Integument;4: Midgut;5: Fat body;6: Silkgland;7: Testis;8: Ovary圖5 家蠶不同組織BmSPH33基因轉錄分析Fig.5 Transcription analysis of BmSPH33 gene in different tissues of Bombyx mori
2.2 BmSPH33基因組織表達譜分析
利用RT-PCR對5齡第3天家蠶各組織BmSPH33基因轉錄情況進行檢測,檢測結果發現,BmSPH33在家蠶中腸組織中特異性表達(圖5)。
2.3 BmNPV感染家蠶后BmSPH33的轉錄分析
為探討BmSPH33基因在家蠶感染BmNPV病毒反應中可能的作用,利用實時熒光定量qRT-PCR對對照組(添食ddH2O)和實驗組(添食BmNPV)BmSPH33基因表達情況進行檢測,確定BmSPH33基因轉錄水平是否發生變化。定量結果發現家蠶在感染BmNPV后BmSPH33基因發生響應(圖6),在感染BmNPV 4 h后BmSPH33基因轉錄水平升高,而感染8、12、24、48 h時呈現下調,這說明家蠶在感染BmNPV后產生了應激響應,其中BmSPH33基因mRNA轉錄水平發生變化。BmSPH33基因mRNA轉錄反應受到BmNPV病毒的誘導,提示BmSPH33基因在大造(P50)家蠶品種的抗病毒作用中發揮功能。
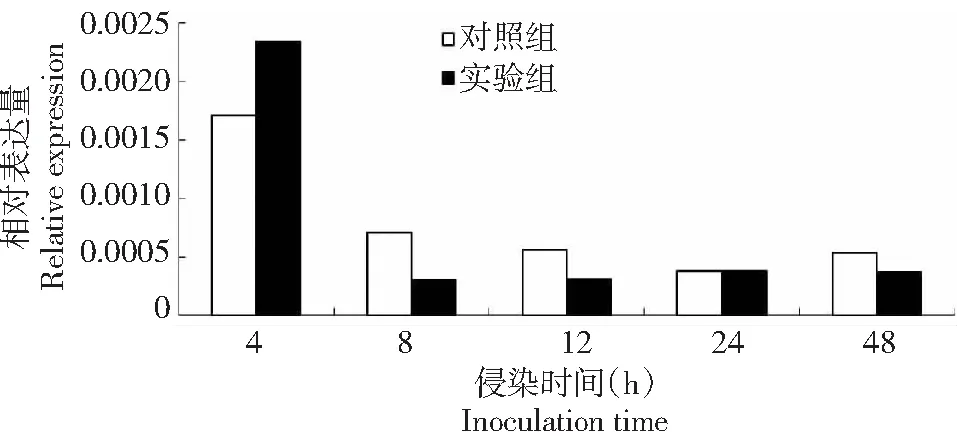
圖6 BmNPV對BmSPH33基因轉錄水平的影響Fig.6 Effects of BmNPV inoculation on transcriptional level of BmSPH33 gene
3 討 論
絲氨酸蛋白酶SPs參與了昆蟲的消化、發育和免疫應答,是一類重要的蛋白水解酶。研究BmSPH33與MacoSPH33氨基酸序列比對發現,它們含有保守的Trypsin-like serine protease的結構域,類胰蛋白酶絲氨酸蛋白酶是一種Clip結構域絲氨酸蛋白酶,被認為能參與昆蟲先天免疫的蛋白酶[19-20],其結構含有保守的TAAHC、DIAL和GDSGGP特征基序。昆蟲經過絲氨酸蛋白酶等一些酶級聯反應能將非活化前體酚氧化酶酶原(prophenoloxidase,proPO)催化形成活性的酚氧化酶原(Phenoloxidase,PO),PO能把酚類氧化成醌類物質,通過生成黑色素沉積在病原物表面,進而黑化和包裹作用消滅病原微生物[21]。SPs蛋白通過剪切羧基端的一些特殊氨基酸而活化proPO[5]。
DNA microarray研究發現64個表達的SPs和SPHs基因中有38個基因中腸轉錄,其中大部分在中腸表達,一些中腸SPHs基因可能由于缺少活性位點殘基而處于非活動狀態,但在其它鱗翅目昆蟲中也發現了表達[22],而且SPH1和SPH2還參與煙草天蛾的proPO活化過程[23];家蠶BmSPH1證實了還參與家蠶的免疫響應[16, 24]。家蠶中腸是消化桑葉并吸收營養的重要場所,BmNPV病毒粒子隨著食物進入中腸,它作為防御病毒入侵的第一道屏障,BmSPH33基因在家蠶中腸組織中特異表達(圖5),研究結果發現,BmSPH33基因轉錄水平在感染4 h時上調,其余時間點(8、12、24和48 h)下調,基因轉錄水平發生變化,表明BmSPH33基因表達受BmNPV誘導而參與家蠶幼蟲免疫應答過程。
絲氨酸蛋白酶在真核細胞和原核生物中是高度保守和普遍存在的,它們已經進化成一個豐富而功能多樣的酶群。絲氨酸蛋白酶是一類主要的消化蛋白酶,占鱗翅目昆蟲消化活性的95 %[25],由于BmSPH33與其BmSPs成員具有Tryp_SPc domain,昆蟲絲氨酸蛋白酶家族作為絲氨酸為活性中心的典型蛋白質水解酶,結合其中腸特異表達,說明該蛋白主要參與食物消化,家蠶感染BmNPV后基因轉錄水平發生變化,推測該基因在家蠶免疫方面起到了輔助作用,但具體功能還有待進一步驗證。
4 結 論
BmSPH33基因在家蠶中腸組織中特異表達,且在家蠶感染BmNPV后發生響應,推測該基因參與家蠶免疫應答過程,結合BmSPH33含有高度保守的胰蛋白酶樣的底物基序,為弄清絲氨酸蛋白酶同源體BmSPHs在消化及免疫應答方面的機制提供參考。
