開花前不同光周期對大豆主要農藝性狀的影響
2021-05-25 07:58:26楊玉花白志元衛保國張瑞軍
西南農業學報 2021年2期
關鍵詞:大豆
楊玉花,雷 陽,白志元,衛保國,張瑞軍
(1.山西農業大學農業基因資源研究中心,農業部黃土高原作物基因資源與種質創制重點實驗室,雜糧種質資源發掘與遺傳改良山西省重點實驗室,山西 太原 030031;2.山西農業大學園藝學院,山西 太原 030031)
【研究意義】日照長度波動會影響大多數生物的發育進程和行為,如哺乳動物和鳥類在長光照的春季適宜繁殖,而昆蟲在短光照的秋季化蛹,這些現象統一稱為光周期現象,可以感知季節的變化和預測環境條件(如溫度和濕度等)[1]。光周期現象最早是Garner和Allard[2]在1920年提出,研究發現一些植物受日照長于臨界日長條件下促進開花的命名為長日植物,反則為短日植物。大豆是典型的短日照植物[2],對光周期反應較為敏感,在不同生態區開花期和成熟期差異較大[3-4],且同一生態區大豆品種對光周期的反應也有差異。大豆育種專家王金陵[5]在1956年就提出開花前的不同光照長度對大豆開花期、株高和其它性狀均有影響。開花后不同品種的大豆對日照長度的反應也有一定差別[6]。韓天富等[7]研究表明大豆從播種至成熟整個生命過程中均存在光周期反應現象。【前人研究進展】前人研究發現,中國大豆不同生態區品種類型開花后光周期敏感性為南方秋豆>南方夏豆>黃淮夏豆>南方春豆、黃淮春豆>北方春豆,不同性狀對光周期處理的敏感性也不相同,如株高>干重>節數>莢數>單株粒數>單株產量[8-9]。何言章等[10-11]對國內各地有代表性的大豆品種進行遮光處理,發現短光敏感性強弱為南方夏大豆>北方夏大豆>南方春大豆>北方春大豆,大豆開花前和開花后不同光周期對大豆農藝性狀均有一定影響,如株高、主莖節數、莖粗、分枝數、莢數、秕莢率、株粒數和百粒重等。費志宏等[12]人利用盆栽暗室和加光等措施研究發現,大豆光周期反應敏感性強弱依次為南方夏大豆>南方春大豆>黃淮海夏大豆>北方春大豆,說明大豆品種的光周期反應敏感性隨著主產地緯度的降低而增強,同一地區品種的光周期反應敏感性具有很大相似性。王英等[13]研究發現,開花后光照處理對大豆農藝性狀和品質性狀均有影響,且不同性狀的光周期敏感度差異明顯,如株高>主莖節數>蛋白質含量、脂肪含量>百粒重>單株莢數>蛋白質和脂肪總量。基于對大豆光周期敏感性的鑒定同時,一些學者對大豆受光周期影響較大的花期進行深入研究,現已經克隆E1~E99個調控大豆花期基因[14-20],這些基因顯性位點能不同程度延長大豆的營養生長或者生殖生長(E6和E9除外),其中E1基因對于抑制大豆開花功能最強。除此之外還有一些基因可以調控大豆開花期,如GmPhyA基因可以調控光周期,影響大豆開花[17-18],GmCRY1a和GmCRY2a基因通過調控生物鐘或者葉片的衰老最終調節大豆光周期開花[21-22],敲除GmRAV2轉錄因子可以使大豆提前開花[23],超表達GmSOC1基因可以促進大豆提前開花[24],而超表達microRNAGmiR156b可以延遲大豆開花[25]等。目前對受大豆光周期影響較大的花期性狀研究較多且機理比較明確,但實際應用于生產相對較少,對受大豆光周期影響的其它性狀,尤其是產量和品質性狀研究較少。因此對于大豆光敏感這個復雜性狀,應該多角度來解析和闡明其機制,為大豆廣適育種提供理論基礎。【本研究切入點】本研究主要利用3份不同大豆資源為研究對象,在開花前不同光周期條件下研究株高,有效分枝數,無效分枝數,主莖節數,一粒莢,二粒莢,三粒莢,四粒莢,空莢,單株有效莢數,單株總莢數,單株粒數,每莢粒數,百粒重,單株粒重,蛋白含量,水分含量,脂肪含量18個農藝性狀,利用方差分析3份大豆資源分別在不同光周期條件下農藝性狀的差異,進行各農藝性狀在不同光周期下的相關性和主成分分析,同時比較各農藝性狀光相對敏感度,揭示不同大豆資源對光周期敏感性存在差異,并且能夠精確了解大豆光敏感的農藝性狀。【擬解決的關鍵問題】此研究結果為后續開展大豆開花前遮光對農藝性狀光敏感度的遺傳分析和相關農藝性狀的QTL定位以及候選基因的分析奠定基礎。
1 材料與方法
1.1 供試材料
本研究利用的3份大豆供試材料均由山西省農業科學院農作物品種資源研究所收集并提供。2017年5-10月分別在山西省農業科學院東陽基地和祁縣基地種植,土壤肥力均勻,按照標準田間管理。
1.2 試驗方法
1.2.1 田間試驗設計 大豆于5月9日統一播種,播種出苗后統一利用覆蓋雙層黑色遮光布的遮光架進行遮光處理,遮光處理為12 h遮光(19:00-7:00),12 h不遮光(7:00-19:00),對照處理為不遮光(自然光)。田間試驗按照完全隨機區組設計3個重復,每個小區3行,行距×株距為50 cm×13.5 cm。
1.2.2 性狀考察 大豆成熟時,每個小區選擇10個代表單株,測定株高,有效分枝數,無效分枝數,主莖節數,一粒莢,二粒莢,三粒莢,四粒莢,空莢,百粒重等性狀,通過測定性狀再計算單株有效莢數,單株總莢數,單株粒數,每莢粒數性狀,然后取均值作為該性狀的最終值。蛋白含量,水分含量,脂肪含量3個性狀由近紅外測定[傅立葉變換近紅外光譜儀(MPA)],最后取均值。具體調查標準參照《中國大豆品種志》[26]。
1.2.3 數據分析 利用SAS V8軟件中的PROC ANOVA程序對2個地點的3份大豆資源2個不同遮光處理下的18個性狀進行方差分析,從而確定大豆的不同性狀是否在不同光周期間存在顯著差異。利用SAS V8軟件中的PROC ANOVA程序進行相關性分析,PROC ANOVA程序進行主成分分析。采用長短日照條件下某一性狀的相對差值RD% =(L-S) /S×100 來評價18個性狀的光周期敏感度[27]。
2 結果與分析
2.1 不同光周期下大豆各農藝性狀的方差分析
通過對3份大豆材料2個不同的光周期調查的18個性狀(表1)進行方差分析,可以看出不同光周期對大豆大多數農藝性狀均有影響,主要表現在株高,有效分枝數,主莖節數,三粒莢,單株有效莢數,單株總莢數,單株粒數和單株粒重等性狀,遮光后的短光周期顯著低于正常光照(P<0.01),然而短光周期條件下一粒莢明顯高于對照(P<0.01)。無效分枝數,四粒莢,空莢,每莢粒數,百粒重,蛋白含量,水分含量和脂肪含量幾乎不受光周期影響,差異不顯著。以上結果說明,短光周期顯著影響大豆部分產量相關性狀,但對本研究采用3份大豆資源的品質性狀影響甚微。
同時發現不同大豆材料不同性狀對光周期影響不同,如晉豆25在短光周期的條件下二粒莢,單株有效莢數,單株總莢數3個性狀反而顯著高于長光照,中黃35在短光周期的條件下的百粒重,蛋白含量,水分含量和脂肪含量明顯高于長光照。說明不同大豆材料對光周期影響不同,同時同一大豆材料不同性狀對光周期影響也不相同。
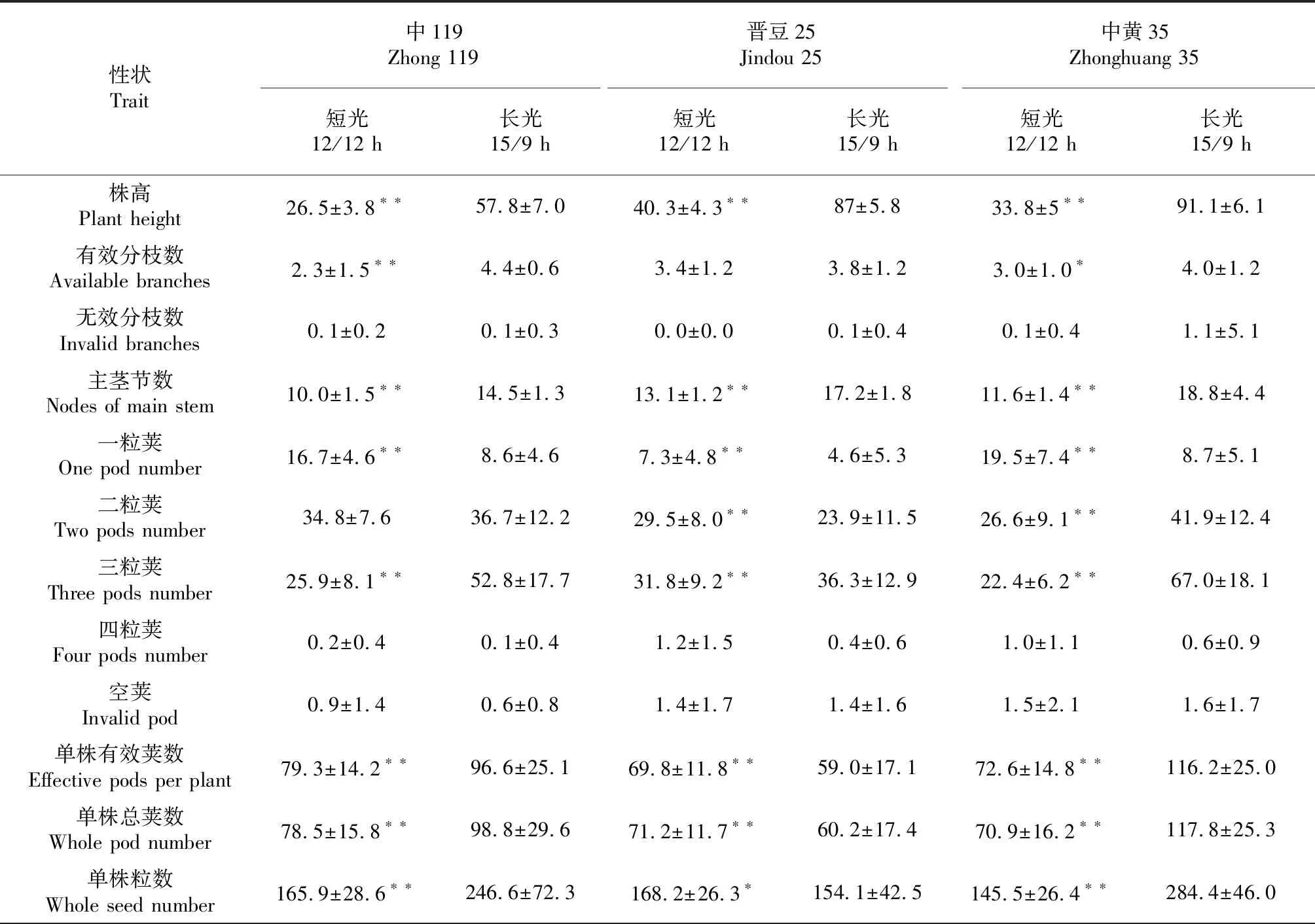
表1 不同光周期對3份大豆材料各性狀的影響
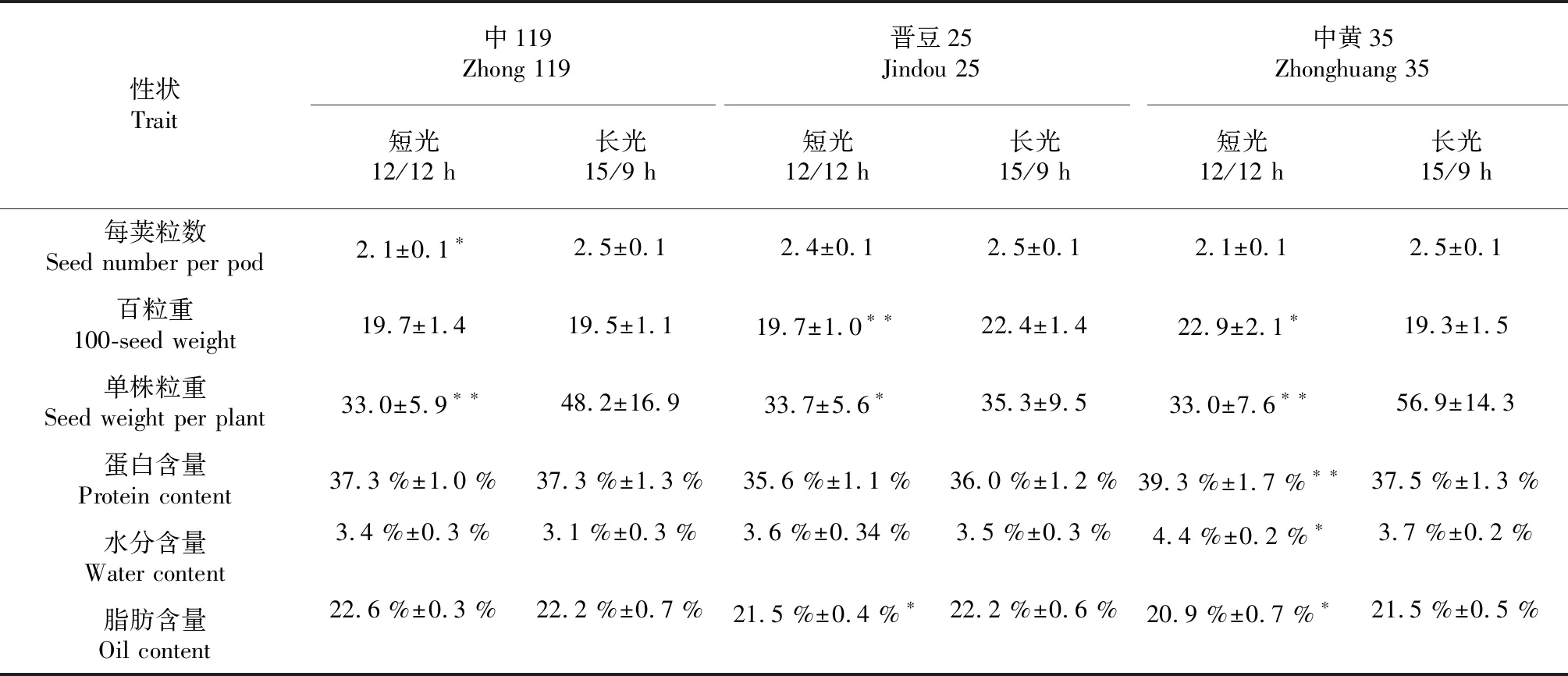
續表1 Continued table 1
2.2 不同光周期條件下大豆各農藝性狀的比較
從表2可以看出,在遮光后短光周期條件下和正常長光周期條件下大多數性狀間呈現顯著或者極顯著相關性,同一性狀在遮光后短光周期和正常長光周期條件下也存在一定的顯著或者極顯著相關性。比如在正常長光周期條件下的株高與遮光后在短光周期條件下的有效分枝數,主莖節數,二粒莢,三粒莢,單株有效莢數,單株粒數,單株粒重和蛋白含量呈顯著或者極顯著正相關,與一粒莢,四粒莢,空莢,單株總莢數,每莢粒數,百粒重,水分含量和脂肪含量呈顯著或者極顯著負相關,與無效分枝數不相關。而在正常長光周期條件下株高與同樣長光周期條件下的有效分枝數,主莖節數,二粒莢,三粒莢,單株有效莢數,單株粒數,單株粒重和蛋白含量呈顯著或者極顯著正相關,與一粒莢,四粒莢,空莢,單株總莢數,每莢粒數,百粒重,水分含量和脂肪含量呈顯著或者極顯著負相關,與無效分枝數不相關。遮光后短光周期和正常長光周期間的株高,有效分枝數,主莖節數,三粒莢,單株粒數,單株粒重和蛋白含量呈顯著或者極顯著正相關,一粒莢,二粒莢,四粒莢,空莢,單株有效莢數,單株總莢數,每莢粒數,百粒重,水分含量和脂肪含量呈顯著或者極顯著負相關,同樣2種處理之間的無效分枝數不相關。由此可以說明,大豆大多數性狀在遮光后短光周期和正常長光周期條件下相互之間的相關性表現基本一致,可見改變光周期后大豆的一些性狀之間的相互關系不受影響。
2.3 大豆各農藝性狀主成分分析
由表3可以看出,遮光后短光周期條件下前6個主成分的貢獻率分別為31.66 %、19.96 %、13.28 % 、7.92 %、6.19 %和5.78 %,累積貢獻率為84.79 %;正常長光周期條件下前6個主成分的貢獻率分別為39.52 %,14.42 %,10.35 %,7.80 %,6.78 %和5.86 %,累積貢獻率為84.72 %,其余12個主成分貢獻率均較小。因此遮光后短光周期條件下可以用前6個主成分包含84.79 %信息很好地概括了大豆18個農藝性狀對光周期的反應,同樣在正常長光周期條件下,大豆18個農藝性狀也可以轉換成前6個相對獨立的主成分,可以代表原來18個性狀84.72 %信息。
2.4 大豆各農藝性狀光周期相對敏感度比較
根據光相對敏感度計算公式分別計算出3個大豆材料(中119,晉豆25和中黃35)18個農藝性狀的平均光敏感度值。從圖1可以看出,中119和中黃35的18個農藝性狀的光相對敏感度強弱相對較一致,而晉豆25的部分農藝性狀的光相對敏感度相對較弱且與中119和中黃35的短光照和長光照敏感度相反。如中119的18個農藝性狀的光相對敏感度強弱順序為株高,三粒莢,有效分枝數,單株粒數,一粒莢,主莖節數,單株粒重,空莢,四粒莢,單株總莢數,單株有效莢數,每莢粒數,水分含量,二粒莢,百粒重,脂肪含量,蛋白含量,和無效分枝數;中黃35的18個農藝性狀光相對敏感度強弱順序為三粒莢,株高,單株粒數,單株粒重,單株總莢數,主莖節數,單株有效莢數,一粒莢,二粒莢,四粒莢,有效分枝數,每莢粒數,空莢,水分含量,百粒重,蛋白含量,脂肪含量和無效分枝數;而晉豆25的18個農藝性狀的光相對敏感度強弱順序為株高,四粒莢,一粒莢,主莖節數,二粒莢,單株有效莢數,單株總莢數,三粒莢,百粒重,有效分枝數,單株粒數,每莢粒數,單株粒重,水分含量,脂肪含量,空莢,蛋白含量和無效分枝數。可以看出大豆株高是對光敏感度最強的一個農藝性狀,其次是三粒莢,有效分枝數,主莖節數,單株粒重,一粒莢,四粒莢等農藝性狀,其中一粒莢和四粒莢受光周期影響與其它性狀相反,也就是說株高,三粒莢,有效分枝數,主莖節數,單株粒重在長光周期條件下比短光周期條件下相對值較高,而一粒莢和四粒莢則相反,在短光周期條件下的相對值較高,該結果與表1中對不同的光周期方差分析結果一致。
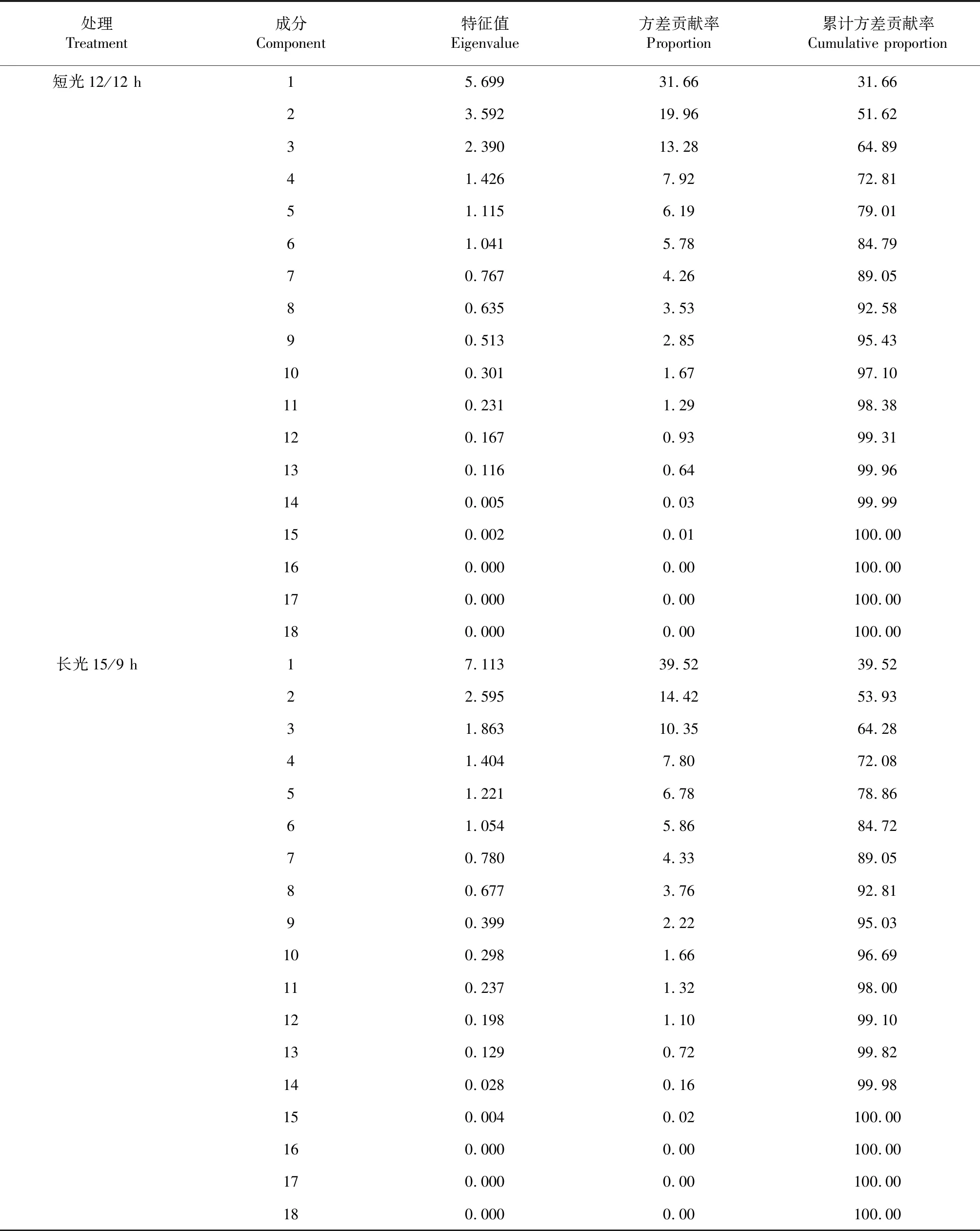
表3 大豆各農藝性狀在不同光周期下的主成分分析
3 討 論
3.1 大豆是一個受光周期影響較大的作物
大豆是一個短日照作物,如果日照長度高于臨界日照將會出現營養生長不能正常轉變生殖生長,導致不能正常開花結實。由于大豆對光周期反應較為敏感且隨著地理緯度的降低而增強,因此在大豆生產上有明顯的地域性限制。以前大多數科研工作者以開花期或者成熟期的變化來研究大豆光周期敏感性,本研究則側重以大豆農藝性狀和品質性狀受光周期的影響來探討大豆光周期敏感性。
通過研究發現大豆株高,有效分枝數,無效分枝數,主莖節數,一粒莢,二粒莢,三粒莢,四粒莢,空莢,單株有效莢數,單株總莢數,單株粒數,每莢粒數,百粒重,單株粒重,蛋白含量,水分含量,脂肪含量18個農藝性狀中大多數性狀受光周期的影響,尤其是株高,有效分枝數,主莖節數,一粒莢,三粒莢,單株粒重等性狀在長光照和短光照不同光周期條件下差異極顯著,這與前人研究結果較一致[8,10-11],進而說明大豆是一個極易受光周期影響的農作物。
3.2 光周期對大豆各農藝性狀的影響
農藝性狀是作物在長期適應環境的過程中,通過進化而產生的,同時也受外界生存環境變化影響。本研究中大豆株高,有效分枝數,主莖節數,百粒重等性狀在不同光周期條件下表現有一定的差異。如在短光周期條件下性狀值明顯低于長光周期,說明開花前短光周期條件會導致大豆營養生長時間縮短,從而促進大豆提前進入生殖生長,這也是作物在特定環境條件下的自身調節反映[28]。同樣在特定條件下各農藝性狀間也具有相關性,如正常長光周期條件下,株高與有效分枝數,主莖節數等農藝性狀存在一定程度的相關,同樣在短光周期條件下,株高與有效分枝數,主莖節數等農藝性狀也存在一定程度的相關,說明作物各農藝性狀間不同程度的相互關聯,同時也顯示作物會隨著不同的生態條件進行自身調節。作物農藝性狀在受到自然因素(不同經緯度、地區氣候)等影響,形成對地理環境因子適應性的響應性狀特征[29-30]。
本研究對3個大豆材料的各農藝性狀的光相對敏感度進行分析發現株高,有效分枝數,主莖節數,一粒莢,三粒莢和單株粒重表現出對光較強的敏感性,說明這6個指標適合作為大豆光敏感評價指標。其次發現不同大豆材料間的農藝性狀光相對敏感度有一定的差別,如中119和中黃35農藝性狀的光相對敏感度較一致,而晉豆25的部分農藝性狀的光相對敏感度則相反,如二粒莢,百粒重等性狀。說明不同大豆材料間對光敏感度有差異,這與前人研究的不同大豆生態區不同品種類型光周期敏感性不同結果一致[8],說明本研究采用的晉豆25與中119和中黃35屬于來源不同生態區大豆品種有關。
3.3 大豆光敏感育種應用
結合前人研究和本研究發現,大豆光周期反應敏感性不僅體現在開花期和成熟期等性狀,同時也體現在一些農藝性狀上。因此可以借鑒小麥育種經驗,通過培育矮化和光不敏感性農藝性狀等品種,推動小麥育種的 “綠色革命”[31]。在大豆育種過程同樣選擇光不敏感性農藝性狀以及對光周期遲鈍型的品種,不斷改善大豆品種的廣生態適應性,或者選擇光敏感性大豆品種,推進大豆“兩系”育種進程。
在育種過程中,結合大豆的開花期,成熟期,農藝性狀,品質性狀以及結合目前已經克隆的調控大豆光周期基因(雖然現在報道的已經克隆的調控大豆光周期基因很多,但是實際應用的很少)等綜合指標,配制大量組合,多年多點跨生態區選擇,最終將大豆廣適性優良基因聚合在一起,選育出大豆廣適性且高產優良品種。以此來突破國產大豆供不應求的局面[32],應對全球大豆貿易格局變化對我國大豆產業所造成的風險。
4 結 論
本研究利用3份來源不同的大豆材料,對其開花前不同光周期處理,通過分析18個不同農藝性狀方差分析、相關性分析、主成分分析和光相對敏感度比較分析,結果表明3份大豆材料各農藝性狀對光周期敏感性有差異,其中株高,株高,有效分枝數,主莖節數,一粒莢,三粒莢和單株粒重表型較強,可以作為大豆光敏感性育種評價指標,同時也可以為大豆廣適性和“兩系”育種提供鑒定依據。
猜你喜歡
農業科技通訊(2023年1期)2023-02-12 07:09:18
今日農業(2022年16期)2022-11-09 23:18:44
中國化肥信息(2022年7期)2022-08-31 01:29:28
中國化肥信息(2022年5期)2022-08-30 01:58:26
今日農業(2021年20期)2021-11-26 01:23:56
今日農業(2021年14期)2021-10-14 08:35:34
下一代英才(酷炫少年)(2018年6期)2018-07-09 03:17:44
農產品市場周刊(2017年4期)2017-03-03 19:40:05
兒童故事畫報·智力大王(2015年10期)2016-01-27 01:01:35
讀寫算(中)(2015年10期)2015-11-07 07:24:12
