基于線粒體Cytb基因序列的華南沿海黃鰭棘鯛種群遺傳結(jié)構(gòu)分析
2021-05-25 07:59:08朱威霖楊春玲陳慧芳何震晗曾地剛陳秀荔
西南農(nóng)業(yè)學(xué)報(bào) 2021年2期
關(guān)鍵詞:分析
朱威霖,肖 珊,楊春玲,陳慧芳,3,何震晗,3,曾地剛,陳秀荔*,彭 敏*
(1.廣西水產(chǎn)科學(xué)研究院/廣西水產(chǎn)遺傳育種與健康養(yǎng)殖重點(diǎn)實(shí)驗(yàn)室,廣西 南寧 530021;2.廣西水產(chǎn)引育種中心,廣西 南寧 530031;3.廣西大學(xué)動(dòng)物科學(xué)技術(shù)學(xué)院,廣西 南寧 530004)
【研究意義】黃鰭棘鯛(Acanthopagruslatus)又名闊黒鯛,隸屬于鱸形目(Percoiformes)鯛科(Sparide)棘鯛屬(Acanthopagrus),為淺海暖水性底層魚(yú)類,原產(chǎn)于太平洋和北印度洋,廣泛分布在我國(guó)、日本、朝鮮、菲律賓和科威特等國(guó)家,在我國(guó)主要分布于東南沿海地區(qū),在廣東俗稱黃腳魚(yú)立、黃絲魚(yú)立、黃立魚(yú)或黃墻,在福建俗稱黃翅,在臺(tái)灣俗稱烏鯮或赤鰭仔[1-2]。黃鰭棘鯛肉質(zhì)鮮美、營(yíng)養(yǎng)價(jià)值較高,口感極佳,是一種名貴的海洋經(jīng)濟(jì)魚(yú)類,也是我國(guó)海水魚(yú)類養(yǎng)殖的重要對(duì)象[3]。近年來(lái),由于環(huán)境污染、過(guò)度捕撈、近親繁殖、人工放流和養(yǎng)殖逃逸等引起黃鰭棘鯛種質(zhì)資源混亂,自然種群嚴(yán)重衰退,導(dǎo)致種質(zhì)資源嚴(yán)重退化、抗病力減弱及養(yǎng)殖性能下降[4]。線粒體DNA(mtDNA)是細(xì)胞核外唯一具有自我復(fù)制功能的DNA,其進(jìn)化效率遠(yuǎn)高于單拷貝核基因,較少的mtDNA樣本即可代表1個(gè)群體[5]。mtDNA由于具有進(jìn)化速度快、母系遺傳、基因組較小、易于測(cè)序分析等特點(diǎn),已成為分子進(jìn)化研究中的首選材料[6-9],其中線粒體細(xì)胞色素b基因(Cytb)序列是mtDNA中最常用的分子標(biāo)記之一,已在魚(yú)類種群遺傳學(xué)和系統(tǒng)發(fā)育學(xué)中廣泛應(yīng)用[10-14],但未見(jiàn)在華南沿海黃鰭棘鯛遺傳多樣性方面的研究報(bào)道。因此,基于線粒體Cytb基因序列分析華南沿海黃鰭棘鯛種群遺傳結(jié)構(gòu),對(duì)促進(jìn)華南沿海黃鰭棘鯛資源的可持續(xù)利用及遺傳多樣性保護(hù)具有重要意義。【前人研究進(jìn)展】胡靜等[15]通過(guò)mtDNACOⅠ和Cytb基因序列分析發(fā)現(xiàn),我國(guó)南海波紋唇魚(yú)的遺傳多樣性處于較低水平,存在遺傳分化現(xiàn)象但分化不顯著。李敏等[16]基于線粒體Cytb基因序列分析發(fā)現(xiàn),我國(guó)近海和陸架的花斑蛇鯔遺傳分化不顯著,在漁業(yè)上可作為一個(gè)單元來(lái)管理。朱傳坤等[15]基于線粒體Cytb基因和D-loop序列分析發(fā)現(xiàn),洪澤湖、千島湖和西湖3個(gè)華鳈群體存在明顯的遺傳分化現(xiàn)象。沈朕等[18]研究顯示,線粒體Cytb基因和D-loop區(qū)均可作為檢測(cè)大瀧六線魚(yú)遺傳多樣性的有效分子標(biāo)記。李大命等[19]分析太湖新銀魚(yú)Cytb和COⅠ基因序列的多態(tài)性發(fā)現(xiàn),太湖新銀魚(yú)的遺傳多樣性處于較低水平,種群遺傳結(jié)構(gòu)不穩(wěn)定。近年來(lái),國(guó)內(nèi)外關(guān)于黃鰭棘鯛線粒體遺傳學(xué)的研究主要有:劉紅艷等[20]對(duì)海南海口市、福建廈門市和廣東珠海市3個(gè)地理群體海捕黃鰭鯛線粒體D-loop基因進(jìn)行多態(tài)性分析:Xia等[21]對(duì)我國(guó)水域黃鰭棘鯛mtDNA控制區(qū)序列變異遺傳進(jìn)行分析;江世貴等[22]對(duì)4種鯛科魚(yú)類Cytb基因序列及分子系統(tǒng)學(xué)進(jìn)行分析;徐田軍等[23]對(duì)舟山海域4種鯛科魚(yú)類線粒體Cytb基因全序列進(jìn)行克隆分析;黃元佳等[24]基于線粒體COⅠ基因的海陵島鯛科魚(yú)類系統(tǒng)發(fā)育分析;陳詠霞等[25]基于線粒體COⅠ基因序列的中國(guó)鯛科魚(yú)類系統(tǒng)進(jìn)化關(guān)系分析;Hsu等[26]基于線粒體和擴(kuò)增片段長(zhǎng)度多態(tài)性的黃鰭棘鯛、黑棘鯛(Acanthopagrusschlegelii)、桔鰭棘鯛(A.sivicolus)、臺(tái)灣棘鯛(A.taiwanensis)、琉球黃鰭棘鯛(A.chinshira)和太平洋棘鯛(A.pacificus)區(qū)分。【本研究切入點(diǎn)】為了保護(hù)和恢復(fù)黃鰭棘鯛海洋資源,我國(guó)華南沿海各地常年大量放流黃鰭棘鯛魚(yú)苗,但目前未見(jiàn)針對(duì)黃鰭棘鯛遺傳結(jié)構(gòu)變化長(zhǎng)期跟蹤及進(jìn)行遺傳多樣性分析的相關(guān)研究報(bào)道。【擬解決的關(guān)鍵問(wèn)題】比較我國(guó)華南沿海黃鰭棘鯛8個(gè)地理群體的線粒體Cytb基因序列,分析其特征和遺傳多樣性背景,了解我國(guó)黃鰭棘鯛群體的遺傳差異和分化情況,并對(duì)其遺傳多樣性進(jìn)行評(píng)價(jià),為黃鰭棘鯛種質(zhì)資源的保護(hù)和利用提供參考依據(jù)。
1 材料與方法
1.1 試驗(yàn)材料
供試黃鰭棘鯛樣本于2017年12月至2018年12月分別采自我國(guó)華南沿海的福建廈門(以下簡(jiǎn)稱XM)、廣東汕尾(以下簡(jiǎn)稱SW)、廣東陽(yáng)江(以下簡(jiǎn)稱YJ)、海南海口(以下簡(jiǎn)稱HK)、海南三亞(以下簡(jiǎn)稱SY)、廣西北海(以下簡(jiǎn)稱BH)、廣西欽州(以下簡(jiǎn)稱QZ)和廣西防城港東興(以下簡(jiǎn)稱FC),共320尾。每尾約取5.0 g肌肉組織存放于95 %乙醇中,-20 ℃保存?zhèn)溆谩R原傊莺{為界,又將黃鰭棘鯛群體劃分為東海域(EAST)組群和西海域(WEST)組群。
1.2 DNA提取、PCR擴(kuò)增與測(cè)序
每尾黃鰭棘鯛分別剪取肌肉組織50.0 mg,采用醋酸銨法[27]提取總DNA,用微量核酸蛋白分析儀測(cè)定樣品DNA濃度和純度,然后以1 %瓊脂糖凝膠電泳檢測(cè)樣品DNA質(zhì)量,檢測(cè)合格的樣品DNA置于-20 ℃保存?zhèn)溆谩R院细竦臉悠稤NA為模板進(jìn)行PCR擴(kuò)增,PCR引物F(5′-GACTTGAAAAACCACCGTTG-3′)和R(5′-GGCGTTCGGCTTACAAAA-3′)委托生工生物工程(上海)股份有限公司合成。PCR反應(yīng)體系50.0 μl:2×ExTaqMastermix Bufffer 25.0 μl,10 μmol/L上、下游引物各2.0 μl,100 ng/μl DNA模板2.0 μl,ddH2O補(bǔ)足至50.0 μl;擴(kuò)增程序:95 ℃預(yù)變性5 min;95 ℃ 30 s,62 ℃ 30 s,72 ℃ 1 min,進(jìn)行40個(gè)循環(huán);72 ℃延伸10 min,4 ℃保存。擴(kuò)增產(chǎn)物經(jīng)1 %瓊脂糖凝膠電泳檢測(cè)合格后送至深圳華大基因科技有限公司測(cè)序。
1.3 統(tǒng)計(jì)分析
利用Chromas 2.33(http://www.technelysium com.au/chromas.html)對(duì)測(cè)序結(jié)果進(jìn)行編輯、校對(duì)和剪切;利用SeqMan 7.1.0(https://www.dnastar.com/)進(jìn)行拼接,以獲得Cytb基因全長(zhǎng)序列;采用MEGA X(https://www.megasoftware.net/)分析序列的堿基組成及變異位點(diǎn)。運(yùn)用DnaSP 5.10(http://www.ub.edu/dnasp/)分析黃鰭棘鯛的單倍型(h)、單倍型多樣性指數(shù)(Hd)、核苷酸遺傳多樣性指數(shù)(π);利用MEGA X分析序列計(jì)算Kimura 2-parameter遺傳距離和分析遺傳結(jié)構(gòu),然后采用遺傳距離、基因流(Nm)、遺傳分化指數(shù)(Fst)評(píng)價(jià)兩兩群體間的遺傳差異,重復(fù)抽樣10000次以檢驗(yàn)Fst結(jié)果的差異顯著性,并構(gòu)建非加權(quán)組平均法(UPGMA)系統(tǒng)發(fā)育進(jìn)化樹(shù),以及進(jìn)行群體間聚類分析。通過(guò)種群分化測(cè)試(Exact test of population differentiation)進(jìn)行隨機(jī)交配群(Population panmixia)假設(shè)即單倍型在群體間隨機(jī)分布的檢驗(yàn),以AMOVA分析估算遺傳變異的分布,10 000次隨機(jī)重復(fù)抽樣后進(jìn)行差異顯著性分析。采用Tajima′sD檢驗(yàn)[28]和Fu′sFs檢驗(yàn)[29]檢測(cè)種群進(jìn)化是否嚴(yán)格遵循中性理論。根據(jù)公式t=Tτ/2u[30]計(jì)算群體自擴(kuò)張開(kāi)始至今的時(shí)間(t),其中T為世代時(shí)間,τ為擴(kuò)張時(shí)間參數(shù)(在Arlequin軟件中根據(jù)核苷酸錯(cuò)配分布計(jì)算獲得),u為序列的突變速率[u=2μk,式中,k為序列長(zhǎng)度,μ為每個(gè)核苷酸位點(diǎn)的突變速率(每百萬(wàn)年突變1.0 %~2.5 %[31])]。
2 結(jié)果與分析
2.1 線粒體Cytb基因序列分析結(jié)果
經(jīng)PCR擴(kuò)增、測(cè)序和校對(duì),獲得8個(gè)黃鰭棘鯛群體的320條線粒體Cytb基因全長(zhǎng)序列(1141 bp),Cytb基因序列中堿基T、C、A和G的平均含量分別為30.3 %、29.4 %、24.8 %和15.5 %,且A+T平均含量(55.1 %)高于C+G平均含量(44.9 %),表現(xiàn)出較強(qiáng)的少G偏倚性,符合脊椎動(dòng)物線粒體DNA序列特征。在Cytb基因序列片段使用的密碼子中,T、C、A和G在第1、2和3位點(diǎn)的平均含量存在明顯差異,其中G平均含量變化最明顯,在第1、2和3位點(diǎn)分別為26.4 %、13.9 %和6.0 %,說(shuō)明密碼子的堿基使用頻率存在明顯偏向性。
在黃鰭棘鯛Cytb基因全長(zhǎng)序列中未檢測(cè)到插入或缺失位點(diǎn),但檢測(cè)到63個(gè)變異位點(diǎn),占總位點(diǎn)數(shù)的5.52 %,其中簡(jiǎn)約信息位點(diǎn)25個(gè)、單一變異位點(diǎn)38個(gè);在XM群體中發(fā)現(xiàn)變異位點(diǎn)最多,為23個(gè),在FC群體中發(fā)現(xiàn)變異位點(diǎn)最少,僅10個(gè)。在63個(gè)變異位點(diǎn)中,59個(gè)為轉(zhuǎn)換、4個(gè)為顛換,轉(zhuǎn)換與顛換比(R值)為14.75,說(shuō)明線粒體Cytb基因序列突變未達(dá)飽和狀態(tài),受進(jìn)化噪音影響的可能性較小。其中,52個(gè)變異位點(diǎn)發(fā)生在密碼子第3位上,10個(gè)變異位點(diǎn)發(fā)生在密碼子第1位上,1個(gè)變異位點(diǎn)發(fā)生在密碼子第2位上。
黃鰭棘鯛Cytb基因部分序列編碼380個(gè)氨基酸殘基,其中亮氨酸平均含量最高,半胱氨酸平均含量最低。4種堿基(T、C、A和G)在氨基酸密碼子3個(gè)位點(diǎn)出現(xiàn)頻率差異的趨勢(shì)一致,均為在密碼子第1位點(diǎn)無(wú)明顯的堿基偏倚性,在第2位點(diǎn)存在明顯的T偏倚和反G偏倚,在第3位點(diǎn)存在反G偏倚。將黃鰭棘鯛群體Cytb基因的核苷酸序列翻譯成氨基酸序列,檢測(cè)到8個(gè)氨基酸變異位點(diǎn),其中7個(gè)由密碼子第1位突變引起,另1個(gè)由密碼子第2位突變引起,其他核苷酸突變未引起氨基酸變異,屬同義突變,據(jù)此定義9個(gè)氨基酸為單倍型,說(shuō)明黃鰭棘鯛的Cytb氨基酸序列具有較低的多態(tài)性。
2.2 單倍型及遺傳多樣性分析結(jié)果
8個(gè)黃鰭棘鯛群體的320條線粒體Cytb基因序列定義62個(gè)單倍型,其中,42個(gè)單倍型僅在一個(gè)個(gè)體中檢測(cè)到,在其余20個(gè)單倍型中,有18個(gè)是群體間共享單倍型,另外2個(gè)單倍型在不止1個(gè)個(gè)體中發(fā)現(xiàn),但這些個(gè)體僅分布在同1個(gè)群體。Hap1為優(yōu)勢(shì)單倍型,在8個(gè)群體中均有出現(xiàn);單倍型Hap4和Hap2分別由8和7個(gè)群體共享;單倍型Hap5、Hap6、Hap8、Hap11、Hap16和Hap37均由3個(gè)以上群體共享;單倍型Hap3、Hap7、Hap9、Hap15、Hap23、Hap26、Hap34、Hap40和Hap54均由2個(gè)以上群體共享;而Hap10為BH群體獨(dú)有,Hap12~Hap14為FC群體獨(dú)有,Hap17~Hap22、Hap24、Hap25和Hap27為HK群體獨(dú)有,Hap28~Hap32為QZ群體獨(dú)有,Hap33、Hap35和Hap36為SW群體獨(dú)有,Hap38、Hap39和Hap41~Hap44為SY群體獨(dú)有,Hap46~Hap53和Hap55~Hap59為XM群體獨(dú)有,Hap60~Hap62為YJ群體獨(dú)有。在62個(gè)單倍型中,7個(gè)單倍型為2個(gè)組群共享,13個(gè)單倍型為WEST組群獨(dú)享,42個(gè)單倍型為EAST組群獨(dú)享。62個(gè)單倍型的出現(xiàn)頻率存在較明顯差異,其中以Hap1出現(xiàn)的頻率最高,占總個(gè)體數(shù)的38.75 %,其次為Hap4和Hap2,分別占總個(gè)體數(shù)的23.75 %和8.44 %。說(shuō)明2個(gè)組群共享、WEST組群獨(dú)享和EAST組群獨(dú)享的單倍型是華南沿海黃鰭棘鯛在長(zhǎng)期進(jìn)化過(guò)程中形成的較穩(wěn)定的優(yōu)勢(shì)基因型。
利用DnaSP 5.10分析8個(gè)黃鰭棘鯛群體的單倍型多樣性和核苷酸多樣性,結(jié)果(表1)表明,8個(gè)黃鰭棘鯛地理群體的Hd在0.57179~0.83718,其中以XM群體最高,為0.83718,其次為BH群體和HK群體,分別為0.74872和0.72821,QZ群體最低,為0.57179;π在0.00089~0.00162,其中BH群體最高,為0.00162,其次為XM群體和HK群體,分別為0.00152和0.00148,SW群體最低,為0.00089。WEST組群和EAST組群的Hd分別為0.66471和0.69714,π分別為0.00128和0.00120,二者差異不明顯。將所有樣本作為一個(gè)整體進(jìn)行分析,發(fā)現(xiàn)黃鰭棘鯛群體的Hd為0.78638,π為0.00164。
2.3 群體系統(tǒng)進(jìn)化關(guān)系分析結(jié)果
根據(jù)黃鰭棘鯛Cytb基因全長(zhǎng)序列,以黑棘鯛線粒體Cytb基因序列(GenBank登錄號(hào)NC_018553.1)作為外群,利用MEGA X分析序列計(jì)算Kimura 2-parameter的遺傳距離并構(gòu)建單倍型非加權(quán)組平均法(UPGMA)系統(tǒng)發(fā)育進(jìn)化樹(shù)及進(jìn)行群體間聚類分析,結(jié)果(圖1)顯示,62個(gè)單倍型系統(tǒng)發(fā)育進(jìn)化樹(shù)可分為三大分支:Group A、Group B和Group C,各節(jié)點(diǎn)的支持率均較低(<30 %),只有極少數(shù)兩兩單倍型分支間的支持率超過(guò)50 %;不同群體的單倍型未按取樣地點(diǎn)有規(guī)律地分布在系統(tǒng)發(fā)育進(jìn)化樹(shù)上,但大部分單倍型按瓊州海峽東、西海域分布在系統(tǒng)發(fā)育進(jìn)化樹(shù)上,即分為EAST和WEST兩個(gè)組群。聚類分析結(jié)果(圖2)也表明,8個(gè)黃鰭棘鯛地理群體以瓊州海峽分隔為瓊州海峽東和瓊州海峽西兩大部分。Hap1和Hap4是高頻單倍型,二者在所有群體中均有出現(xiàn),以Hap4為主體的Group B包含了WEST組群(FC、QZ和BH群體)絕大部分個(gè)體(87.74 %),以Hap1為中心的Group A則涵蓋了EAST組群(HK、SY、YJ、SW和XM群體)絕大部分個(gè)體(87.38 %)。
2.4 群體遺傳結(jié)構(gòu)分析結(jié)果
利用MEGA X分析測(cè)序獲得320尾黃鰭棘鯛樣品的線粒體Cytb基因序列,得到8個(gè)黃鰭棘鯛地理群體間的遺傳距離(表2)。由表2可知,群體內(nèi)遺傳距離為0.00089~0.00162,各地理群體內(nèi)的遺傳距離排序?yàn)锽H>XM>HK>FC>SY>YJ>QZ>SW;從群體間遺傳距離來(lái)看,QZ群體與XM群體的遺傳距離最遠(yuǎn),為0.00232,而遺傳距離最近的為SW群體與YJ群體(0.00093);8個(gè)黃鰭棘鯛地理群體間的Nm為0.39049~無(wú)窮大(inf),WEST組群內(nèi)兩兩間的Nm為8.77670~inf,EAST組群內(nèi)兩兩群體間的Nm均為inf,WEST組群與EAST組群兩兩群體間的Nm為0.39049~1.36673,BH群體與EAST組群間的Nm為1.00378~1.36673,WEST組群與EAST組群的Nm以BH群體為過(guò)渡。說(shuō)明EAST組群和WEST組群內(nèi)基因交流頻繁,組群間由于地理隔離而限制了群體的基因交流。
由表3可知,8個(gè)黃鰭棘鯛地理群體間的Fst在-0.01300~0.56149,其中,SW群體與QZ群體的Fst最大(0.56149),且差異極顯著(P<0.01,下同),說(shuō)明其遺傳分化程度最大,而SY群體與YJ群體間

表1 黃鰭棘鯛的單倍型及遺傳多樣性信息
的Fst(最小-0.01300),說(shuō)明其遺傳分化程度最小。在EAST組群內(nèi),F(xiàn)st均為負(fù)值,但相互間差異不顯著(P>0.05,下同);而在WEST組群內(nèi),最大分化出現(xiàn)在BH群體與QZ群體間(Fst=0.05390,P=0.00901),且差異顯著(P<0.05,下同);而WEST組群與EAST組群兩兩群體間的Fst為0.26785~0.56149,且差異極顯著。相對(duì)于組群內(nèi),WEST組群與EAST組群的兩兩群體間遺傳距離和遺傳分化指數(shù)較大,基因流較小,說(shuō)明WEST組群與EAST組群存在明顯的遺傳分化。綜合8個(gè)黃鰭棘鯛地理群體的聚類分析結(jié)果及組群間的Fst分析結(jié)果可知,瓊州海峽與海南島形成的天然地理屏障限制了8個(gè)黃鰭棘鯛地理群體的基因交流,且WEST組群與EAST組群存在明顯的遺傳分化,在漁業(yè)管理上可將瓊州海峽以東和瓊州海峽以西的黃鰭棘鯛群體作為2個(gè)管理單位(MUs)進(jìn)行管理。
對(duì)以樣品來(lái)源劃分的EAST組群(HK、SY、YJ、SW和XM群體)和WEST組群(FC、QZ和BH群體)黃鰭棘鯛群體,通過(guò)AMOVA計(jì)算其遺傳變異分布,結(jié)果(表4)顯示組群間、組群內(nèi)群體間和群體內(nèi)個(gè)體間的變異分別為41.26 %、0.09 %和58.65 %,經(jīng)檢驗(yàn)均達(dá)顯著或極顯著差異水平,進(jìn)一步說(shuō)明華南沿海黃鰭棘鯛EAST組群和WEST組群內(nèi)群體間分化較小,組群間由于地理隔離限制了組群間的基因交流,而WEST組群和EAST組群內(nèi)的群體基因交流因不受限制而無(wú)群體分化。將所有黃鰭棘鯛個(gè)體按單倍型類群可分成三部分(Group A、Group B和Group C)進(jìn)行AMOVA分析,結(jié)果(表4)顯示70.98 %變異來(lái)自單倍型類群間。
Exact檢驗(yàn)結(jié)果(表3)表明,單倍型在群體間的隨機(jī)交配假設(shè)檢驗(yàn)水平不均勻,其在EAST組群與WEST組群間表現(xiàn)為顯著(P=0.00000),不符合單倍型在群體間群體是隨機(jī)分布的假設(shè),而在WEST組群內(nèi)(P=0.49038)和EAST組群內(nèi)(P=0.70135)表現(xiàn)不顯著,符合單倍型分別在WEST組群和EAST組群內(nèi)群體是隨機(jī)分布的假設(shè)。
2.5 群體動(dòng)態(tài)分析結(jié)果
由表5可知,單倍型類群Group C的Tajima’sD和Fu’sFs檢驗(yàn)結(jié)果均為負(fù)值,但未達(dá)顯著差異水平,而個(gè)體數(shù)占絕大部分的單倍型類群Group A和Group B的Tajima’sD和Fu’sFs檢驗(yàn)結(jié)果為負(fù)值,但均存在顯著差異,單倍型類群總體的Tajima’sD和Fu’sFs檢驗(yàn)結(jié)果也存在顯著差異,說(shuō)明序列中含有比中性進(jìn)化模型更多的核苷酸位點(diǎn)變化,預(yù)示被研究群體可能經(jīng)歷了一個(gè)擴(kuò)張歷史。核苷酸錯(cuò)配分析結(jié)果(圖3)表明,單倍型類群Group A和Group B均為單峰分布,Group C為雙峰分布,但單倍型類群總體上呈雙峰分布,可能是多個(gè)單倍型類群疊加的結(jié)果。可見(jiàn),Tajima’sD和Fu’sFs檢驗(yàn)及核苷酸錯(cuò)配分析結(jié)果均表明華南沿海黃鰭棘鯛群體可能經(jīng)歷了種群擴(kuò)張事件。采用Arlequin分析獲得擴(kuò)張時(shí)間參數(shù)τ=2.42773,以每百萬(wàn)年突變2.0 %為線粒體Cytb基因的核苷酸進(jìn)化速率,根據(jù)黃鰭棘鯛存在1~2齡雄性性腺發(fā)育成熟后2~3齡轉(zhuǎn)變?yōu)榇菩孕阅孓D(zhuǎn)現(xiàn)象,本研究參考施曉峰等[32]、洪萬(wàn)樹(shù)等[33]的方法以生物學(xué)最小齡2年齡作為一個(gè)世代的時(shí)間跨度,并根據(jù)公式τ=2ut,計(jì)算得到華南沿海黃鰭棘鯛群體擴(kuò)張時(shí)間約為5.319萬(wàn)年前的更新世晚期。綜上所述,采用無(wú)限突變位點(diǎn)模型Tajima’sD和Fu’sFs、線粒體DNA單倍型錯(cuò)配分布等方法對(duì)8個(gè)黃鰭棘鯛地理種群的歷史變動(dòng)情況進(jìn)行調(diào)查,可判斷華南沿海黃鰭棘鯛群體可能經(jīng)歷了種群擴(kuò)張事件,擴(kuò)張時(shí)間約在5.319萬(wàn)年前。
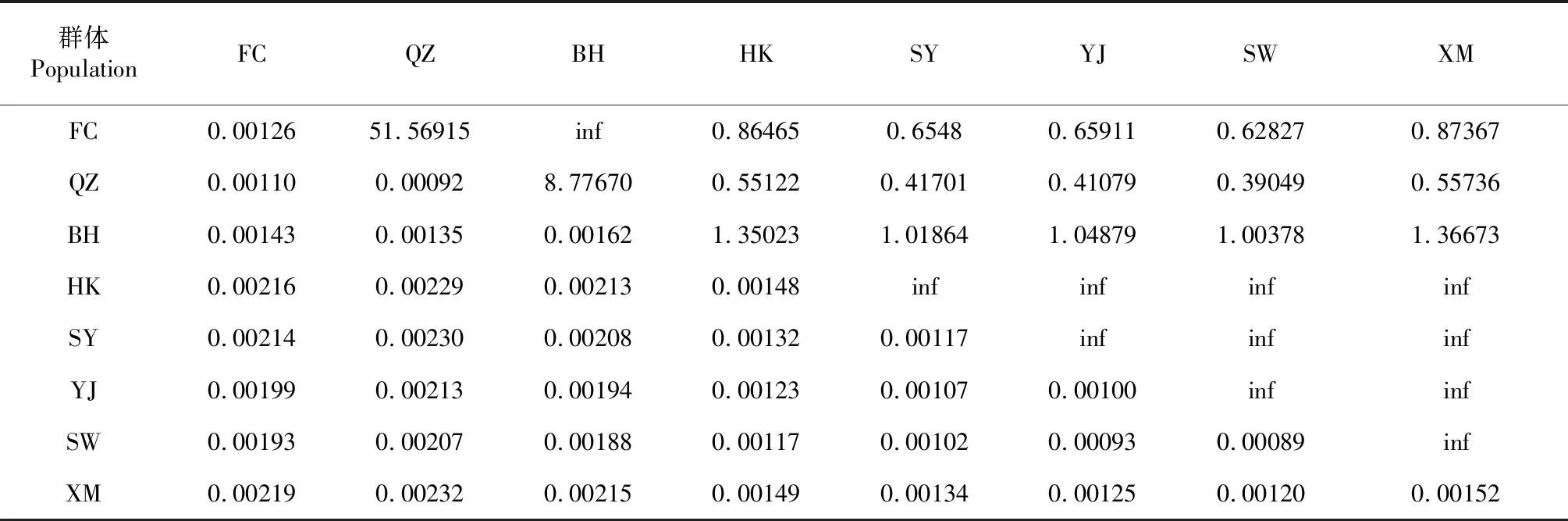
表2 8個(gè)黃鰭棘鯛地理群體間的遺傳距離(對(duì)角線下方)、群體內(nèi)遺傳距離(對(duì)角線上)和Nm(對(duì)角線上方)比較
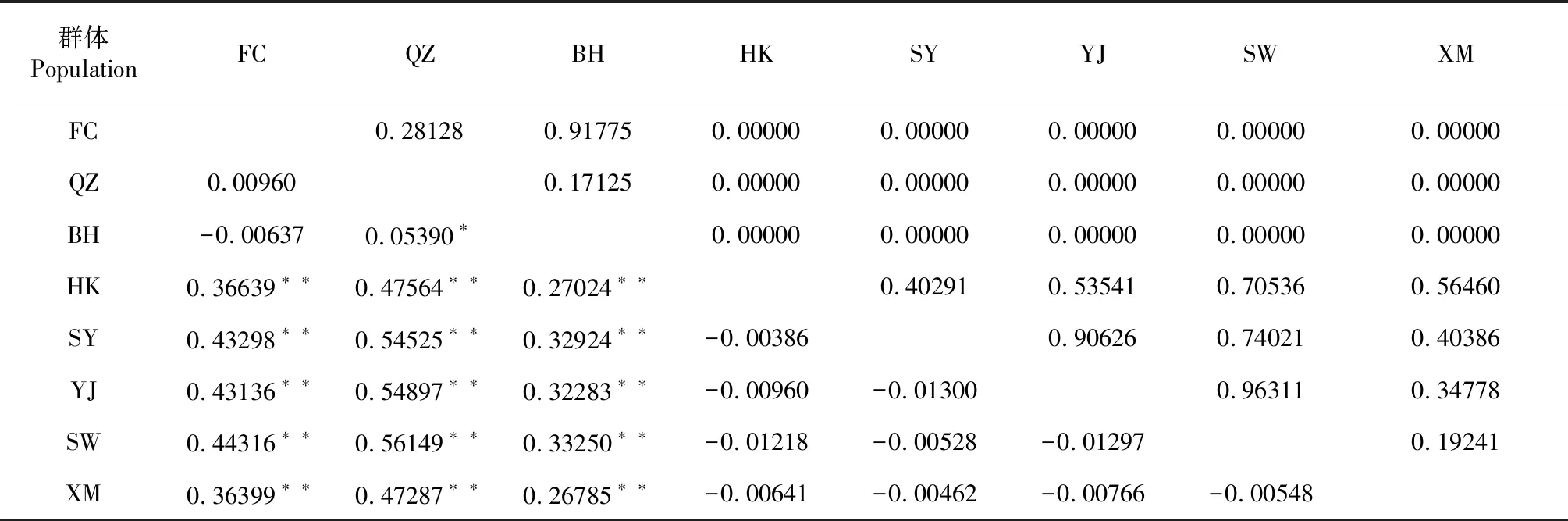
表3 8個(gè)黃鰭棘鯛地理群體間的Fst(對(duì)角線下方)及其Exact 檢驗(yàn)(對(duì)角線上方)結(jié)果
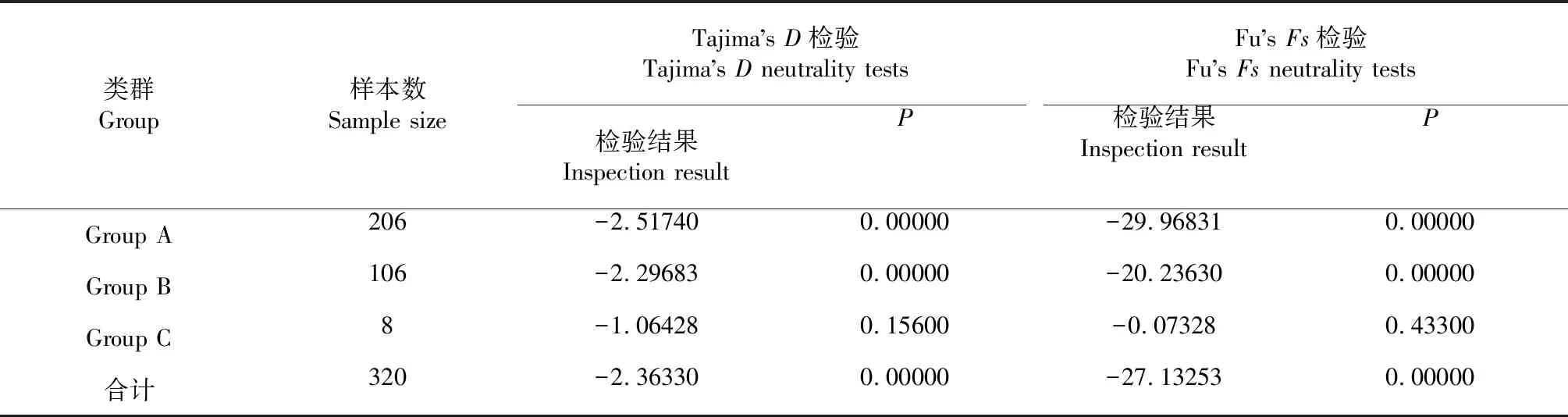
表5 Tajima’s D和Fu’s Fs檢驗(yàn)結(jié)果
3 討 論
遺傳多樣性是物種長(zhǎng)期進(jìn)化或群體持續(xù)生存并適應(yīng)不斷變化環(huán)境而進(jìn)化的結(jié)果,物種遺傳多樣性或變異性越豐富,表明物種的進(jìn)化潛力越大,對(duì)環(huán)境改變響應(yīng)的進(jìn)化能力越強(qiáng)[34-35]。核苷酸多樣性指數(shù)和線粒體DNA的單倍型多樣性指數(shù)是衡量DNA多態(tài)程度的兩個(gè)重要指標(biāo)[36]。本研究中,8個(gè)黃鰭棘鯛地理種群基于線粒體Cytb基因序列分析的單倍型多樣性指數(shù)在0.57179~0.83718,群體的單倍型多樣性指數(shù)為0.78638,核苷酸多樣性指數(shù)在0.00089~0.00162,總體為0.00164。相對(duì)于Grant和Bowen[37]的分類結(jié)果,本研究中8個(gè)黃鰭棘鯛群體基于線粒體Cytb基因全長(zhǎng)序列的遺傳多樣性符合高Hd低π類型,具有較高的Hd(>0.5)和較低的π(<0.005)可能是種群歷史上經(jīng)歷瓶頸效應(yīng)后快速擴(kuò)張的表現(xiàn);與同樣基于線粒體Cytb基因序列標(biāo)記分析的同海域其他魚(yú)類相比,8個(gè)黃鰭棘鯛群體的多樣性低于花斑蛇鯔[16]、短尾大眼鯛[38]、大黃魚(yú)[39]和黃鰭?cǎi)R面鲀[40],高于龍頭魚(yú)[41],與波紋唇魚(yú)[15]和藍(lán)圓鲹[42]接近。可見(jiàn),基于線粒體Cytb基因的黃鰭棘鯛多樣性處于中等偏低水平。魚(yú)類的遺傳多樣性受捕撈壓力影響,過(guò)度捕撈會(huì)顯著降低其遺傳多樣性[43]。因此,在漁業(yè)資源開(kāi)發(fā)利用中。正確認(rèn)識(shí)和評(píng)價(jià)魚(yú)類的遺傳多樣性水平,是可持續(xù)開(kāi)發(fā)利用該漁業(yè)資源的基礎(chǔ)。
Tajima[28]研究認(rèn)為,應(yīng)用Tajima’sD和Fu’sFs檢驗(yàn)推測(cè)種群歷史時(shí),若Tajima’sD與Fu’sFs檢驗(yàn)結(jié)果呈負(fù)值,且在統(tǒng)計(jì)學(xué)上達(dá)顯著水平,說(shuō)明序列中含有比中性進(jìn)化模型更多的核苷酸位點(diǎn)變化,預(yù)示著種群可能曾經(jīng)經(jīng)歷一個(gè)擴(kuò)張歷史。本研究中,單倍型類群總體的Tajima’sD和Fu’sFs檢驗(yàn)結(jié)果均為顯著負(fù)值,且個(gè)體數(shù)占絕大部分的單倍型類群Group A和Group B的核苷酸錯(cuò)配分布圖為單峰,說(shuō)明華南沿海黃鰭棘鯛群體發(fā)生了群體擴(kuò)張,并依據(jù)Arlequin分析所得擴(kuò)張時(shí)間參數(shù)推算擴(kuò)張時(shí)間約在5.319萬(wàn)年前的更新世晚期。
單倍型間的遺傳距離是衡量一個(gè)物種或群體線粒體變異程度的重要指標(biāo),也是衡量群體間遺傳變異程度的可靠參數(shù),遺傳距離越大表明群體間親緣關(guān)系越遠(yuǎn)。本研究利用MEGA X根據(jù)線粒體Cytb基因序列計(jì)算8個(gè)黃鰭棘鯛群體間的遺傳距離為0.00093~0.00232,群體內(nèi)的遺傳距離為0.00089~0.00162,群體間和群體內(nèi)的遺傳距離較小,均遠(yuǎn)低于0.05000,且群體間遺傳距離與群體內(nèi)遺傳距離無(wú)明顯差異,說(shuō)明8個(gè)黃鰭棘鯛群體間存在較近的親緣關(guān)系,與Shaklee[44]對(duì)魚(yú)類種群、種和屬水平上的分類依據(jù)吻合;UPGMA聚類分析結(jié)果表明,大部分單倍型按瓊州海峽東、西海域分布在系統(tǒng)發(fā)育進(jìn)化樹(shù)上,Group A涵蓋了EAST組群絕大部分個(gè)體(87.38 %),Group B包含了WEST組群絕大部分個(gè)體(87.74 %);群體聚類分析結(jié)果也表明,8個(gè)黃鰭棘鯛地理群體以瓊州海峽分隔為兩部分。一般認(rèn)為,群體間的Nm<1.00000表示群體可能由于遺傳漂變而發(fā)生分化,而Nm>1.00000表示群體間的Nm水平較高,群體間遺傳分化較小;當(dāng)Nm>4.00000時(shí),種群間的基因交流更充分,遺傳分化更小[45]。本研究中EAST組群和WEST組群內(nèi)的Nm均>4.00000,表明組群內(nèi)的基因交流充分,遺傳分化更小;而EAST組群與WEST組群間,以BH群體為過(guò)渡,其余Nm均小于1.00000,表明組群間可能發(fā)生遺傳分化。Wright[46]提出的遺傳分化標(biāo)準(zhǔn)為:Fst≤0.05,無(wú)遺傳分化;0.05 以瓊州海峽為界分隔的東海域和西海域兩個(gè)華南沿海黃鰭棘鯛組群(EAST和WEST)間的Fst為0.26785~0.56149,遺傳分化明顯。因此,可將我國(guó)華南沿海黃鰭棘鯛作為2個(gè)管理單位(MUs)進(jìn)行漁業(yè)管理。4 結(jié) 論
猜你喜歡
現(xiàn)代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機(jī)設(shè)計(jì)與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
當(dāng)代經(jīng)濟(jì)研究(2016年5期)2016-12-01 03:12:05
現(xiàn)代農(nóng)業(yè)(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國(guó)中醫(yī)藥現(xiàn)代遠(yuǎn)程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學(xué)學(xué)報(bào)(社會(huì)科學(xué)版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
