不同糖種類及濃度對百子蓮‘Big Blue’體胚發生的影響
2021-05-25 07:58:50岳建華史亞冰李文星李文楊
西南農業學報 2021年2期
岳建華,史亞冰,李文星,董 艷,張 燕,李文楊*
(1.信陽農林學院園藝學院,河南 信陽 464100;2.信陽農林學院林學院,河南 信陽 464100;3.信陽市大別山區園藝植物遺傳改良重點實驗室,河南 信陽 464100)
【研究意義】碳源可為植物離體培養提供能量,合成細胞碳骨架,參與滲透壓、氧化脅迫平衡的調節[1]。不同種類及濃度碳源可顯著調節植物離體培養過程中的形態建成,影響植物器官發生途徑(Organogenesis)或體細胞胚胎發生(Somatic embryogenesis,SE)效果[2]。不同植物、同種植物在離體培養不同階段所需要的碳源種類及濃度不同[2]。蔗糖為最普遍應用的碳源,牡丹(Paeoniasuffruticosa)組織培養時,蔗糖效果優于可溶性淀粉、無水葡萄糖[3];麥芽糖利于棉花(Gossypiumhirsutum)胚性愈傷組織誘導,葡萄糖利于其胚狀體的形成和植株再生[4];3 %蔗糖利于布朗百合(Liliumbrownii)離體再生[5];2 %葡萄糖利于山核桃(Caryacathayensis)胚性愈傷組織的誘導,而5 %的蔗糖利于其體胚萌發[6]。綜上,碳源種類及濃度可顯著影響離體培養的效果[2,7],對特定物種特定發育階段進行精細的碳源調控可顯著提升離體培養的效果。【前人研究進展】百子蓮‘Big Blue’(Agapanthuspraecoxssp.orientalis‘Big Blue’)又稱藍百合,為藍色大花品種,花姿優雅,具有極高的觀賞價值,且抗逆性極強,在國內已逐步推廣,市場前景廣闊[8]。植物組織培養,尤其是SE技術成為百子蓮快繁的重要途徑[9]。雖然百子蓮的SE體系已經建立[9-10],但植物體胚發生的機制仍是未解之謎[11],導致體胚發生效率較低。離體條件下,植物SE過程是一個受激素[12]、碳源[2,13]、活性氧(Reactive oxygen species,ROS)及培養環境[14]共同調控的復雜發育過程[15],目前百子蓮SE體系的研究僅以PIC為激素、蔗糖作為碳源[9-10]。【本研究切入點】碳源對植物離體培養具有重要影響,但對百子蓮SE的調節效果未見系統研究,尤其是在成熟胚發育階段的調節效果及機理鮮有報道。【擬解決的關鍵問題】本研究擬利用胚性愈傷(Embryogenic callus,EC)為試材,通過蔗糖、葡萄糖、麥芽糖及濃度(2 %~4 %)[13]的比較,優化百子蓮‘Big Blue’體胚發生體系,為百子蓮SE快繁提供技術參考。同時,通過測定糖代謝、激素代謝、氧化脅迫等生理指標,初步揭示碳源種類及濃度對成熟胚發育的生理調節機制,豐富植物體胚發生理論。
1 材料與方法
1.1 試驗材料
百子蓮‘Big Blue’購自南京市彩虹花卉公司,2018年5月,選取5年生植株未開裂的小花苞,切取小花梗作為外植體進行愈傷組織的誘導,參照何葉[16]的方法。愈傷組織每月繼代1次,7~9個月后,得到單細胞起源的EC,繼代6個月后用于試驗研究。選取狀態一致、顏色鮮黃、結構疏松的EC。
1.2 試驗設計
按照表1糖種類及濃度進行培養基配制。
1.3 培養基配制
稱取MS干粉(Phytotech Lab M519)4.43 g·L-1、
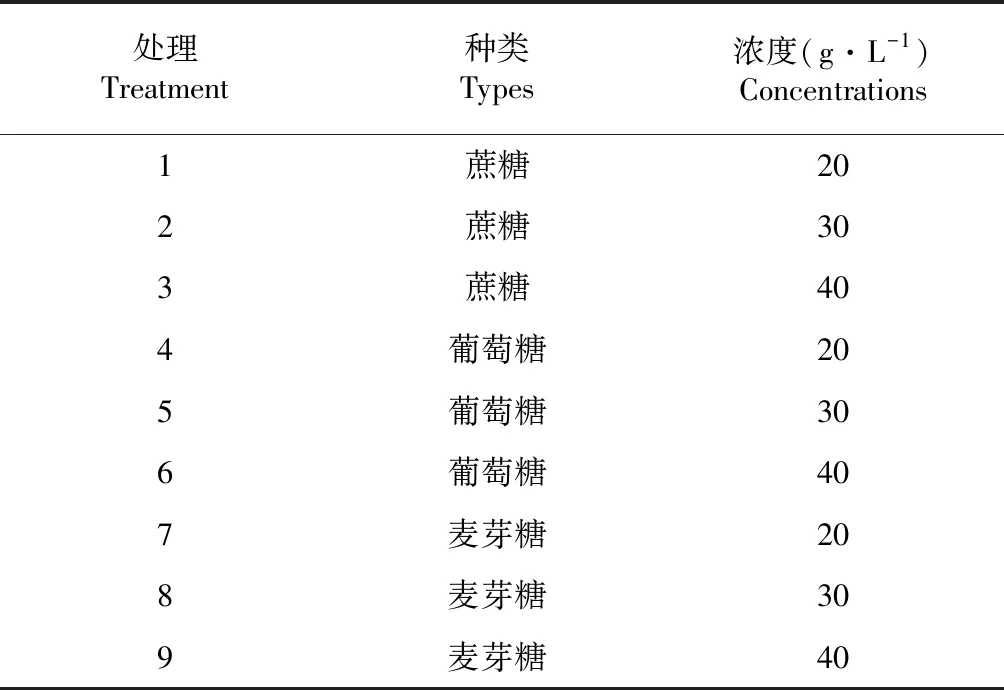
表1 培養基中糖種類及濃度
按2 %、3 %、4 %(w/v)濃度分別加入3種糖,用磁力攪拌器溶解混勻,1 mol/L NaOH溶液調pH至6.0,加瓊脂粉7.0 g·L-1,121 ℃高壓滅菌25 min,降溫至60 ℃置于超凈工作臺內,分裝至90 mm× 16 mm玻璃培養皿,每培養皿分裝25 mL培養基,冷卻至室溫后接種。
1.4 材料接種
體胚誘導階段,每個培養皿平板接種1.0 gEC,平分為大小基本相同的7個細胞團,每處理接種5個培養皿,重復3次。獲得均勻一致的幼胚用于成熟胚發育階段實驗,接種方法同體胚誘導。
1.5 培養條件
幼胚誘導、成熟胚發育過程設溫度(25±2) ℃,相對濕度65 %,暗培養1周后,轉至光下培養(14 h/10 h,光/暗),光照強度為50 μmol·m-2·s-1。光下繼續培養3周后統計數據。
1.6 數據記錄
以誘導獲得的幼胚及成熟胚數量作為數據統計指標,分別統計幼胚(≥1 mm,乳白色)和成熟胚(≥5 mm,具有完整植株結構,形態學上端轉為綠色)數量。
1.7 生理測定
采用酶聯免疫吸附法(ELISA)測定糖類、激素、活性氧與保護酶等生理指標,參照Pradko(2015)的方法[17],委托上海酶聯生物科技有限公司測定。
1.8 數據分析
用Excel 2016、GraphPad Prism 5、Adobe Acrobat DC、Adobe Photoshop軟件進行數據整理和制圖,用IBM SPSS Statistics 20數據分析軟件進行顯著性檢驗(Duncan法,P< 0.05)。
2 結果與分析
2.1 不同糖種類及濃度對百子蓮幼胚誘導的影響
由圖1可知,3種糖及其濃度對體胚誘導效果具有顯著影響,蔗糖的誘導效果相對較好,其次為葡萄糖、麥芽糖。其中3 %蔗糖處理成胚數量最多,平均為117.33個,顯著高于其他各處理(P< 0.05),幼胚呈現松散的顆粒狀,乳白色。3種濃度葡萄糖處理,對成胚數量影響不顯著,其中3 %葡萄糖處理成胚數量較多,其次為2 %和4 %葡萄糖處理;葡萄糖處理幼胚的一致性較好,尺寸略小于蔗糖處理。以麥芽糖作為碳源的成胚數量較少,同步化效果較差。
2.2 不同糖種類及濃度對百子蓮成熟胚發育的影響
不同種類和濃度糖對百子蓮成熟胚發育存在顯著影響,蔗糖相對較好,其次為葡萄糖、麥芽糖(圖2)。其中2 %蔗糖處理成熟胚數量最多,為51.75個,顯著高于其他各處理(P<0.05,圖2-B),多為鮮綠色(圖2-A),而4 %麥芽糖處理效果最差,平均為4.00個。隨蔗糖濃度的提高,成熟胚數量逐漸減少。3種葡萄糖濃度處理對百子蓮成熟胚數量影響不顯著,其中2 %葡萄糖處理成熟胚數量相對較多。麥芽糖作為碳源的成熟胚數量最少,處理間差異不顯著,3 %效果相對較好,多為黃綠色(圖2-A)。
2.3 不同糖濃度對百子蓮成熟胚發育糖代謝的影響
碳源對百子蓮成熟胚發育期糖代謝具有顯著影響(圖3)。總糖含量測定結果顯示,葡萄糖和麥芽糖處理整體上高于蔗糖處理,隨著蔗糖濃度的升高總糖含量逐漸降低,與成熟胚數量的變化規律基本一致。而葡萄糖處理下,百子蓮總糖含量差異不顯著。麥芽糖處理下百子蓮總糖含量最高(圖3-A);百子蓮淀粉含量的變化規律與總糖差異較大,蔗糖處理下淀粉含量相對較高,其中3 %蔗糖處理百子蓮淀粉含量最高(圖3-B);百子蓮體胚組織內蔗糖含量在3種糖類濃度處理下呈現相似的變化規律,3 %濃度處理下百子蓮內源蔗糖含量較低(圖3-C);葡萄糖處理百子蓮內源葡萄糖含量相對高,其中3 %葡萄糖處理顯著高于其他處理(圖3-D);百子蓮麥芽糖的變化規律和總糖呈現出相似的規律,蔗糖處理下呈現梯度變化規律(圖3-E);不同碳源處理下,百子蓮果糖含量的差異相對較小(圖3-F)。成熟胚發育階段糖代謝分析結果表明,總糖、淀粉、蔗糖與麥芽糖含量與成熟胚發育相關性較強。
2.4 不同糖濃度對百子蓮成熟胚發育內源激素含量的影響
碳源對成熟胚激素代謝存在顯著影響(圖4)。蔗糖處理下,IAA含量與成熟胚數量變化趨勢相反,葡萄糖和麥芽糖處理下,IAA含量與成熟胚數量變化趨勢相同,而IAA-氧化酶活性和IAA含量的變化趨勢相反(圖4-A,圖4-C)。隨著碳源濃度的升高,GAs總量出現升高的趨勢(圖4-D),GAs組分中GA3和GA4含量高于GA1;蔗糖處理下,GA1和GA4含量隨濃度的升高而升高,而GA3含量隨濃度升高而降低;葡萄糖處理下,GA4含量與成熟胚數量呈現相反的變化規律;麥芽糖處理下,GA1和GA3含量與成熟胚數量呈現相反的變化規律(圖4-E,圖4-G)。CTK和BR是除了生長素和GA之外調節植物生長的重要激素,蔗糖和麥芽糖處理下,兩者含量變化呈現相反的規律(圖4-H,圖4-I)。蔗糖處理下,ABA含量相對較低,而葡萄糖和麥芽糖處理下,ABA含量相對較高(圖4-J)。乙烯、JA含量的變化幅度相對較小,且趨勢相似(圖4-K,圖4-L)。激素定量結果表明,生長素、GA因具有梯度變化規律,在成熟胚發育過程中可能作用顯著。
2.5 不同糖濃度對百子蓮成熟胚發育活性氧含量及保護酶活性的影響
不同糖處理對活性氧及保護酶活性具有顯著影響(圖5)。ROS活性與成熟胚發育數量趨勢相似,在2 %蔗糖、2 %葡萄糖、3 %麥芽糖處理下相對較高(圖5-A)。蔗糖處理下,·OH含量整體上高于葡萄糖、麥芽糖,但不同濃度蔗糖處理·OH含量差異不顯著(圖5-B)。H2O2含量整體上隨著糖濃度增加而升高,葡萄糖處理下呈現梯度變化(圖5-C)。
保護酶活性變化幅度大于ROS及其組分,POD、CAT活性規律性相對較弱(圖5-D,圖5-E)。SOD活性隨著蔗糖濃度的升高而升高,但與成熟胚發育效果趨勢相反(圖5-F)。
3 討 論
除了光自養體系,碳源是植物離體培養中不可或缺的因素[15]。碳源可構成細胞碳骨架,并為細胞呼吸代謝提供底物與能量[1],植物對碳源的需求因種類、基因型、發育階段均有差異[15]。本研究發現百子蓮體胚誘導及發育成熟階段對碳源濃度的需求分別為3 %和2 %,蔗糖可作為百子蓮SE體系的優選碳源,葡萄糖利于體胚同步化,而麥芽糖效果較差,與山核桃(Caryacathayensis)基本一致[6]。蔗糖可通過改變培養基的滲透壓,使細胞失水,內含物濃度升高,影響體細胞胚的數量及成熟[18],例如內源蔗糖的積累對荔枝(Litchichinensis)SE效率至關重要[19]。而在成熟胚的發育及生根過程中,低濃度的碳源可促進淀粉向蔗糖的轉化,使體胚內滲透壓升高,造成細胞吸水,致使代謝增強,另一方面,成熟胚發育過程中,較低的滲透壓利于根系從培養基中吸收養分。
碳源在體胚發生中起著重要作用,不僅為細胞分裂、分化提供能量,參與信號轉導,物質運輸,還可調節細胞糖代謝,進而影響植物生長[15]。碳源種類和濃度在體胚的發生和成熟發育階段代謝復雜[13,15]。趙舒野[20]發現蔗糖可以顯著提高胡桃楸(Juglansmandshurica)體胚發生,挪威云杉(Piceaabies)體胚成熟過程中,總可溶性糖的含量降低,蔗糖相對含量升高,利于成熟胚發育[21],與本研究結果一致。也有研究表明麥芽糖由于分解較慢,可為體胚后期成熟階段提供更穩定的能量供給,從而利于體胚發生[22]。淀粉的積累通常作為體胚發生中的重要事件,可在體胚發育過程中快速分解,為細胞旺盛分裂活動提供能量[11,22],例如荔枝(Litchichinensis)SE過程中,在淀粉酶、轉化酶作用下,淀粉可轉化為蔗糖,滿足體胚發生物質能量需求[19]。本研究中2 %蔗糖處理的淀粉含量顯著低于3 %、4 %蔗糖處理,內源蔗糖含量顯著高于3 %蔗糖處理,而該處理成熟胚發育效果最佳,表明淀粉向蔗糖的轉化可能為體胚發育提供了充足的能量,而內源蔗糖的積累對體胚發育更為關鍵[19]。
植物激素是自身合成與代謝的產物,調控細胞分化、生長方向與進程[23]。生長素、GA、CTK、ABA、乙烯等對體胚誘導具有重要作用,但因物種而異[24]。本研究中生長素、GA組分隨蔗糖濃度呈梯度變化,且與成熟胚誘導效果關系密切,其作用可能相對重要。雙子葉植物,例如寧夏枸杞(Lyciumbarbarum),體胚誘導時需要加入2,4-D[25],而百子蓮等單子葉植物胚性誘導及維持則需加入PIC[9,16]。2,4-D和PIC均為外源激素類物質,通過調節細胞內源IAA的合成與信號調控體胚發生[23,26]。IAA可通過調節細胞極性誘導體胚形成,因此體胚誘導階段通常需要較高濃度的IAA,內源IAA含量是調節體胚發生的主導因素[27-28]。本研究中,IAA在蔗糖處理下呈現梯度變化,濃度與成熟胚數量趨勢相反,可能是因為體胚誘導階段積累的IAA較多,而在體胚成熟發育階段,高濃度IAA對成熟胚發育不利,因為高濃度的IAA會抑制頂端優勢的形成,不利于成熟胚發育成植株。GA對體胚發生的效果因物種而異,多數研究表明高濃度GA對SE誘導不利[29-30],本研究中,具有生物活性的GA1、GA4與成熟胚數量相反,與紫花苜蓿(Medicagosativa)[31]、天竺葵(PelargoniumhortorumBailey)[29]結果一致。
離體培養產生的氧化脅迫可導致代謝紊亂,甚至細胞死亡[32]。植物通過增強抗氧化酶活性清除ROS[33]。不適的糖種類及濃度會對植物造成脅迫,而ROS和保護酶活性存在動態平衡。在ROS類別中,H2O2可影響基因表達,是非生物脅迫重要的信號分子之一[34-35]。中等強度的H2O2對棉花、牡丹(Paeoniasuffruticosa)離體培養有促進作用[24,36]。蒔蘿(Anethumgraveolens)在愈傷組織誘導體細胞胚階段,H2O2含量持續上升,球形胚時期達到最高,而成熟胚時期含量下降[37]。在本研究中,H2O2含量隨糖濃度的升高而增加,SOD活性加強,而最終成熟胚發育效果隨糖濃度升高而降低,表明H2O2含量、SOD活性增強時脅迫相對嚴重,對成熟胚發育不利。
4 結 論
綜上,糖種類及濃度顯著影響百子蓮體胚發生,蔗糖效果優于葡萄糖和麥芽糖,其中3 %蔗糖利于體胚誘導,2 %蔗糖利于成熟胚發育。培養基碳源對體胚成熟發育期糖代謝、激素代謝及氧化脅迫具有顯著調節作用。淀粉與蔗糖之間的代謝平衡對百子蓮成熟胚發育具有決定作用;高濃度的IAA、GAs對成熟胚發育不利;H2O2、SOD不利于成熟胚發育。
