氧化還原介體強化厭氧活性污泥發酵產氫特征
2021-05-29 03:55:14張立國艾冰凌李建政班巧英
中國環境科學 2021年5期
張立國,艾冰凌,李建政,班巧英*
氧化還原介體強化厭氧活性污泥發酵產氫特征
張立國1,2,艾冰凌3,李建政4,班巧英1,2*
(1.山西大學環境與資源學院,山西 太原 030006;2.山西省黃河實驗室,山西 太原 030006;3.中國熱帶農業科學院海口實驗站,海南 海口 571101;4.哈爾濱工業大學環境學院,黑龍江 哈爾濱 150090)
為提高厭氧污泥的發酵產氫能力,采用間歇培養方式考察了氧化還原介體(ROMs)對厭氧污泥發酵葡萄糖產氫效能的影響,并通過Illumina MiSeq測序揭示了ROMs對微生物群落的影響.結果表明,在發酵液體積為100mL及葡萄糖初始濃度500mg/L條件下,對照的累計產氫量和最大產氫速率(max)分別為11.0mL和0.28mL/h.當腐殖酸和蒽醌-2-磺酸鈉(AQS)作為ROMs加入發酵體系后,厭氧污泥的產氫能力顯著提高,其累計產氫量和max分別比對照高出56.4%、13.6%和 53.6%、10.7%.相反,氧化石墨烯(GO)、指甲花醌和蒽醌-2,6-二磺酸鈉(AQDS)導致厭氧污泥的產氫能力受到不同程度的抑制.Illumina MiSeq測序揭示了發酵系統的優勢菌群存在顯著差異.對照系統中的優勢菌群主要來自5個屬(、、、、),腐殖酸、AQS和指甲花醌發酵系統的主要優勢菌群為、、,而、、、為AQDS和GO發酵系統的主要菌群.冗余分析的結果表明,產氫量與和呈正相關,而與和呈負相關.
厭氧活性污泥;氧化還原介體;發酵產氫;微生物群落
隨著化石能源的日益減少,可再生能源的開發與應用迫在眉睫.氫氣因其具有熱值高、無污染、可再生等優點而成為一種理想的替代燃料.與物理、化學方法相比,生物制氫可以利用各種有機廢水、固體廢棄物為原料產氫,因此受到研究者的廣泛關注[1-2].目前,厭氧細菌暗發酵制氫是研究較多的生物制氫方法之一,該法能夠利用廉價的有機廢棄物為原料進行低成本制氫,但產氫效率有待提高[3-4].因此,強化厭氧細菌暗發酵產氫效能對于提高有機廢棄物生物制氫效能具有重要意義.
氧化還原介體(ROMs)可以加速氧化還原反應過程中的電子傳遞速度,使反應速率提高1個到幾個數量級[5].近年來,ROMs被廣泛用于強化污染物的去除[5-7].有機物厭氧發酵產氫是通過一系列氧化還原反應實現的.可見,ROMs的生物催化功能有望提高厭氧污泥的產氫能力,進而改善有機物暗發酵產氫效能.因此,本研究通過間歇試驗考察了蒽醌-2,6-二磺酸鈉(AQDS)、蒽醌-2-磺酸鈉(AQS)、指甲花醌、氧化石墨烯(GO)、腐殖酸對厭氧污泥發酵葡萄糖產氫的強化效果,并通過高通量測序揭示了ROMs對產酸發酵相關微生物菌群的影響,為ROMs強化有機物厭氧發酵制氫提供新的技術參考.
1 材料與方法
1.1 接種污泥和試驗廢水
接種污泥取自太原市某城市污水處理廠的缺氧池,將污泥混合液置于燒杯中靜置6h,棄上清后用于試驗. 污泥濃度為2.4g MLVSS/L.本研究中所用有機廢水為人工合成廢水,以5000mg/L葡萄糖作為唯一碳源,并添加微生物生長繁殖所需要的其他營養元素[8].
1.2 ROMs固定化
選AQDS、AQS、指甲花醌、GO、腐殖酸,作為強化葡萄糖厭氧生物制氫的ROMs.其中,AQDS、AQS和指甲花醌采用海藻酸鈣包埋法進行固定[8].
1.3 ROMs強化有機廢水厭氧發酵產氫試驗
試驗采用間歇培養方式進行,實驗容器為300mL厭氧瓶.每個厭氧瓶中加入10mL接種污泥和10mL試驗廢水,然后向厭氧瓶中加入一定量空白海藻酸鈣珠子(對照),AQDS、AQS、指甲花醌海藻酸鈣珠子,GO和腐殖酸,使ROMs的終濃度均為80mg/L,加入適量蒸餾水使液相總體積達到100mL.用1mol/L HCl調培養液pH值至6.5.連續通入5min N2后立即密封.隨后用注射器向厭氧瓶中加入0.02%氯仿抑制產甲烷菌的活性. 將所有厭氧瓶置于35℃、120r/min條件下恒溫震蕩培養.每組試驗設置3個平行樣,數據分析取其平均值.每12h測定氣體組成,并于發酵結束時測定揮發酸(VFAs)濃度.
1.4 Illumina MiSeq測序
稱取0.15g污泥(濕重),采用DNA提取試劑盒(E.Z.N.ATMMag-BindSoilDNAKit,,Inc., USA)提取污泥總DNA.以樣品DNA為模板,用通用引物27F,5′-AGAGTTTGATCMTGGCTCAG-3′, 515R,5′-CACGTATTACCGCGGCTGCTGGCAC- 3′對細菌的16S rRNA基因V1~V3區進行PCR擴增.構建Illumina平臺文庫并測序. Illumina MiSeq測序原始序列提交NCBI,登錄號為PRJNA670924.使用Mothur軟件(version v.1.30.1)對測序獲得的原始序列進行質控篩選和過濾,按照97%相似性進行操作分類單元(OTU)聚類分析,在聚類過程中去除嵌合體,得到各OTU的代表序列,在Silva數據庫(http://www.arb-silva.de)進行比對、物種注釋,最后在門水平和屬水平進行群落結構分析.冗余分析采用軟件Canoco 5.0進行.
1.5 分析項目及方法
生物量(揮發性懸浮固體總量MLVSS)采用標準方法測定[9],產氣量通過10~50mL的玻璃注射器排氣計量.發酵氣體的組分和揮發酸濃度分別采用山東滕州瑞普分析儀器有限公司的RP-6800A型(TCD檢測器)和RP-6890型(FID檢測器)氣相色譜測定.累計氫氣產量參照Owen法進行計算[10].
1.6 動力學分析
不同ROMs作用下的產氫動力學分析參照Gompertz模型(式1).

式中:為反應(h)累計氫氣產量,mL;max為最大產氫量,mL;max為最大產氫速率,mL/h;為延遲時間,h; e為常數.
將累計甲烷產量和相應反應時間代入式(1),用統計軟件Origin 9.0計算出max、max、λ[10].
2 結果與討論
2.1 累計氫氣產量
ROMs可以提高電子從初級電子供體傳遞到最終電子受體的速度,從而提高氧化還原反應速率[5].在已有文獻中,醌類物質和腐殖質是研究較多的ROMs[5].因此,本研究考察了AQDS、AQS、指甲花醌、GO和腐殖酸對厭氧活性污泥發酵產氫的影響.如圖1所示,在培養12h后,對照的累計產氫量為2.7mL,隨著培養時間延長,累計產氫量逐漸提高,經過84h連續培養后,達到平臺期,累計產氫量為11.0mL.當加入ROMs后,厭氧污泥發酵產氫能力受到不同程度的影響.其中,腐殖酸和AQS可提高厭氧污泥發酵產氫的效能.腐殖酸試驗組在培養初期就表現出較高的活性,培養12h后的累計氫氣產量達到了4.0mL,比同期對照組高出48.1%,隨著培養時間延長,累積氫氣產量逐漸增加,最終在72h達到穩定,為17.2mL,比對照高出56.4%.研究表明,腐殖酸的電子傳遞能力主要依賴醌、酚等官能團的存在[11].盡管AQS也能提高厭氧活性污泥的產氫能力,但其強化效果低于腐殖酸.當AQS作為ROMs時,培養前期表現出與對照相似的產氫活性,但在培養24h后,AQS試驗組的產氫能力高于對照,并在84h達到最大值,比對照高出13.6%.腐殖酸和AQS能強化厭氧污泥發酵產氫效能主要是由于它們能夠降低葡萄糖氧化還原反應的活化能、提高產氫微生物的能量利用效率[5].然而,當GO、指甲花醌和AQDS作為ROMs時,厭氧活性污泥的產氫能力卻受到不同程度的抑制.在接種初期,它們的發酵產氫速率就低于對照,隨著培養時間延長,累積產氫量逐漸增加并最終分別穩定在5.6,2.1和10.1mL.除此之外,從圖1可知,指甲花醌試驗組存在明顯的耗氫現象,可能是由于指甲花醌的加入促進了一些耗氫菌的生長.
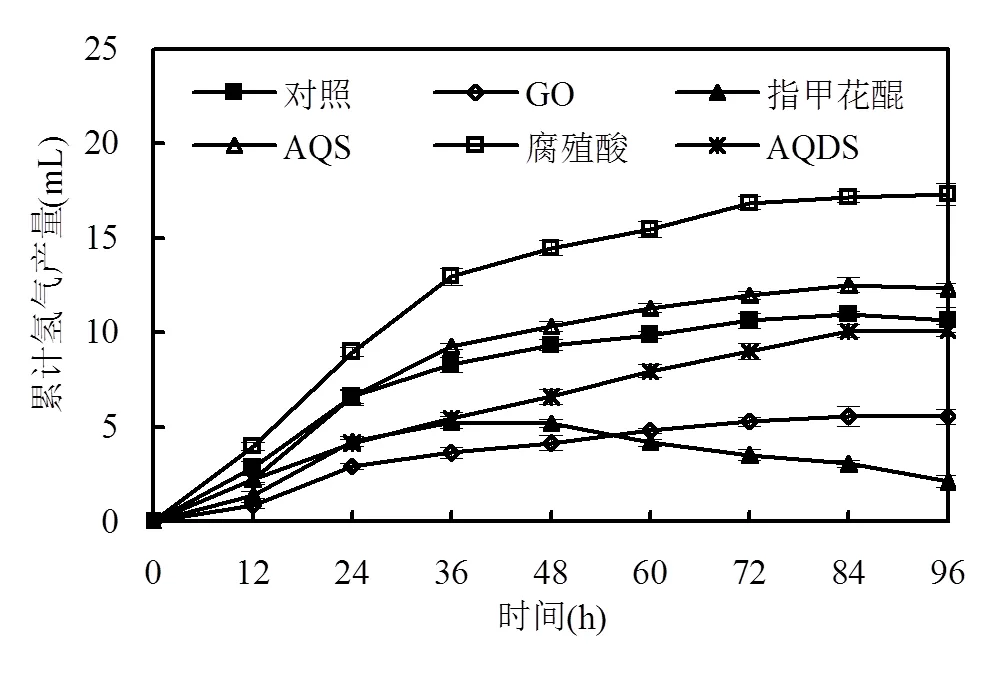
圖1 不同ROMs條件下累積產氫量
2.2 動力學分析
為了解各發酵系統的動力學特征,采用改進的Gompertz模型(式(1))對累計氫氣產量曲線進行了非線性擬合(表1).如表1所示,除指甲花醌發酵系統外,其余發酵系統的擬合系數(2)均大于0.99,表明該模型用來描述累計產氫量的進程是合理的.在對照組中,λ為3.19h,當系統中加入AQS和腐殖酸后,λ分別比對照縮短了11.3%和31.3%.與之相反,加入AQDS和GO之后,明顯延長,分別為3.65 和3.75h.由表1的擬合結果可以發現,腐殖酸發酵系統中的最大產氫速率(max)和最大氫氣產量(max)分別為0.43mL/h和17.1mL,顯著高于對照組,分別比對照提高了53.6%和58.3%.其次,AQS發酵系統的max和max也高于對照組,分別比對照高出10.7%和14.8%.然而,GO發酵系統的max和max卻顯著低于對照組.盡管AQDS發酵系統的max與對照相似,但max卻比對照減少了42.9%.產氫動力學的結果表明,腐殖酸可以顯著強化厭氧活性污泥的產氫能力,而AQDS和GO卻抑制了產氫菌的活性.
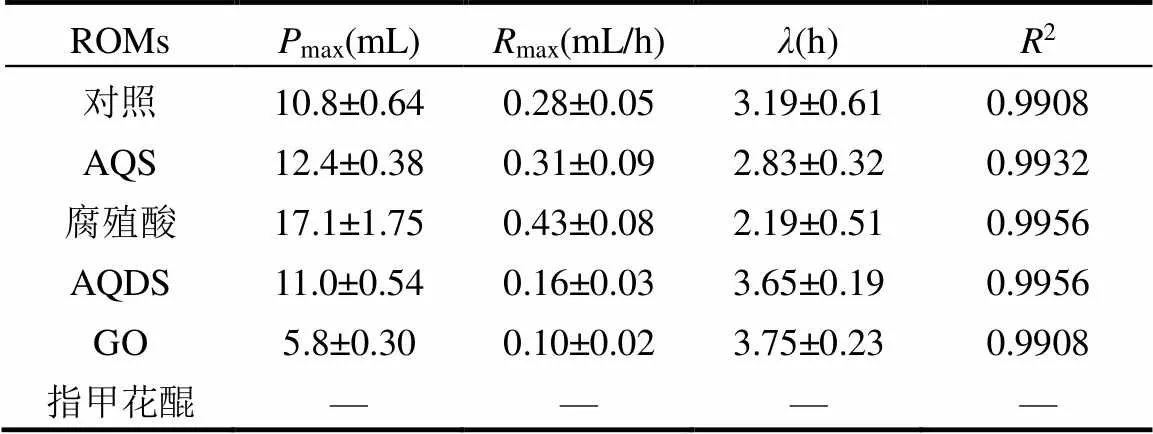
表1 不同ROMs條件下產氫動力學分析
注:“-”表示該系統不符合Gompertz模型.
2.3 液相末端產物
有機物厭氧發酵產氫過程通常伴隨著揮發酸的產生[12].如圖2所示,對照組中的液相末端產物以乙醇、乙酸和丙酸為主,含量分別為71.2,174.4和92.6mg/L.而丁酸含量顯著低于其他揮發酸,僅占揮發酸總量的6.1%.類似地,當AQDS、GO和指甲花醌作為ROMs時,乙醇、乙酸和丙酸仍然為主要液相末端產物,濃度分別為33.7~67.3,89.8~184.4和82.7~ 134.3mg/L.然而,當AQS和腐殖酸作為ROMs時,丁酸濃度顯著高于其他發酵系統,分別為129.5和83.6mg/L,成為主要液相末端產物之一.此外,AQS和腐殖酸發酵系統的揮發酸總量也高于其他發酵系統,表明AQS和腐殖酸能夠提高產酸發酵菌群的活性.相反,GO和指甲花醌發酵系統的揮發酸總量比分別對照低10.0%和24.9%,表明GO和指甲花醌不僅抑制了產氫菌的活性(圖1),同時對其他發酵菌也有一定的抑制作用.

圖2 ROMs對發酵液相末端產物的影響
結合圖1和圖2可以發現,AQS和腐殖酸的加入能夠在一定程度上提高產丁酸菌的活性,從而提高氫氣產量.另外,腐殖酸發酵系統中丙酸含量僅為49.6mg/L,有利于后續降解的進行.
2.4 微生物群落結構
2.4.1 多樣性和豐富度 厭氧活性污泥的產氫效能與其群落結構密切相關[13].本研究采用Illumina MiSeq測序技術對不同發酵系統中厭氧污泥進行了群落結構解析.如表2所示,6個樣品包含的有效序列數目為30599~31860.基于97%相似性對序列進行OTUs聚類分析,在對照、AQS、腐殖酸、AQDS、GO和指甲花醌污泥樣品中分別獲得3423,3574, 3766,3644,3638和3099個OTUs.多樣性指數(Shannon和Simpson)表明,6個污泥樣品中微生物具有相似的多樣性,而對照和AQS組的豐富度指數(Chao1和Ace)低于其他發酵系統. 盡管Chao1和Ace估計的OTUs數目高于實際檢測值,然而文庫覆蓋率(Coverage)達到了93%以上,表明樣品中主要微生物已被檢測到.
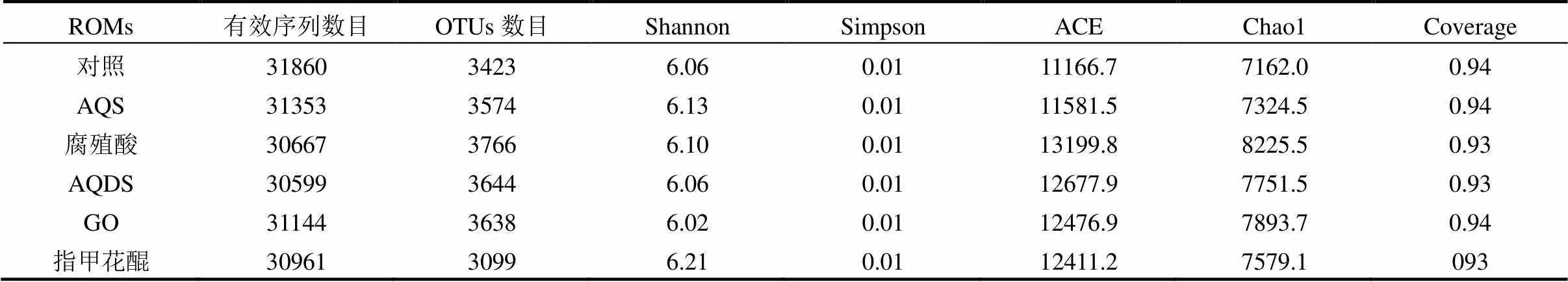
表2 不同ROMs條件下微生物群落多樣性和豐富度分析
2.4.2 微生物群落結構相似性 為揭示不同ROMs發酵系統中微生物群落結構的差異性,對所有樣品進行主坐標分析(PCoA),并通過Venn圖揭示了OTUs分布特征.如圖3(a)所示,PCoA1和PCoA2的貢獻率分別為54.0%和24.0%. PCoA分析結果表明,對照和GO及AQDS和AQS發酵系統中主要微生物相似性較高,而指甲花醌和腐殖酸發酵系統中的主要微生物與其他樣品差異較大. Venn圖進一步揭示了6個樣品中共有和獨有的OTUs數目.由圖3(b)可知,790個OTUs存在于所有樣品中.而對照、AQS、腐殖酸、AQDS、GO和指甲花醌樣品中獨有的OTUs數目分別為819,938, 1042,1023,980和1017.
2.4.3 主要微生物種群的相對豐度 如圖4(a)所示,6個樣品中OTUs所代表的序列在系統發育上涉及8個門,即:變形菌門(Proteobacteria)、綠彎菌門(Chloroflexi)、浮霉菌門(Planctomycetes)、擬桿菌門(Bacteroidetes)、厚壁菌門(Firmicutes)、放線菌門(Actinobacteria)、酸桿菌門(Acidobacteria)和綠菌門(Ignavibacteriae).其中,變形菌門、綠彎菌門、擬桿菌門、厚壁菌門和放線菌門為中溫厭氧反應器中常見的水解發酵菌群[14].與已有研究結果相似,本研究發現變形菌門和綠彎菌門為所有樣品的優勢細菌門.在對照和AQDS中,變形菌門的相對豐度分別為33.3%和33.5%,而AQS、腐殖酸、GO和指甲花醌的加入刺激了變形菌門微生物的生長,使其相對豐度比對照高出6.3%~19.2%.相反,綠彎菌門的相對豐度在對照、AQDS和GO中較高,而在AQS、腐殖酸和指甲花醌中的相對豐度較低.另外,在對照中浮霉菌門的相對豐度為11.5%,ROMs的加入抑制了它們的生長,而ROMs(GO除外)使厚壁菌門的相對豐度比對照顯著提高了0.3~2.2倍.其余細菌門的相對豐度在各樣品之間沒有顯著差異.從圖4b可以看出,在屬水平上未分類和其他序列的相對豐度高達45.5%~52.4%和20.4%~30.4%.由此可見,這些未知功能的微生物種屬在葡萄糖發酵產氫過程中起著重要作用.此外,高通量測序共檢測到11個主要菌屬.在對照組中,(10.9%)、長繩菌屬(2.8%)、(2.2%)、(2.6%)、動性桿菌屬(2.2%)為主要菌群,它們能夠利用一些碳水化合物、酵母粉、蛋白胨等進行生長繁殖[15-18].當加入ROMs后,、和的生長受到不同程度的抑制,其相對豐度較對照降低了15.9%~56.0%.相反,ROMs促進了的生長繁殖,腐殖酸使得的相對豐度比對照高出1.7倍.除了以上主要菌群外,在對照組中還存在少量其他菌群.可以小分子有機酸(如延胡索酸、乳酸)為碳源,其相對豐度為1.7%,加入ROMs后,其數量顯著增加[19].(產酸發酵菌)、磂黃果菌屬(紫色硫細菌)和小梨形菌屬(專性好氧菌)在6個樣品中的相對豐度相似,為0~1.4%之間[20-21].另外,由圖4b可知,產酸發酵菌屠場桿狀菌屬和束毛球菌屬只存在于部分樣品中.在AQS、腐殖酸和AQDS樣品中的相對分度分別為0.7%、1.0%和0.7%,而(1.1%)僅存在于腐殖酸樣品中.


圖4 不同ROMs條件下細菌群落的相對豐度

2.5 優勢功能菌群與產氫效能的關系
為了解發酵系統中優勢微生物種群與厭氧污泥產氫效能之間的關系,對優勢微生物和產量、VFAs進行冗余分析(RDA)(圖5).結果表明,產氫量與、、、和呈正相關,且這些菌群在腐殖酸發酵系統中的相對含量較高,使得腐殖酸系統的累計氫氣產量最大(圖1).從圖5還可以看出,產氣量與、、和呈負相關,它們主要分布在指甲花醌系統中,導致指甲花醌系統的累計產氫量僅為2.1mL,且存在明顯的耗氫現象.另外,RDA結果顯示,氫氣產量與丁酸、乙醇含量正相關,而與丙酸呈負相關,表明丁酸型、乙醇型發酵有利于氫氣產生,而丙酸型發酵不利于氫氣產生,這與以前研究結果一致[22].

圖5 優勢菌群、產氫量和VFAs之間的冗余分析
3 結論
3.1 腐殖酸能夠顯著提高厭氧污泥的產氫能力,其累計氫氣產量達到了17.2mL,比對照高出56.4%,且max比對照提高了53.6%;然而,AQDS、GO和指甲花醌對厭氧污泥發酵產氫有一定的抑制性.
3.2 對照、AQDS、GO和指甲花醌發酵系統的液相末端產物以乙醇、乙酸和丙酸為主,而在AQS和腐殖酸系統中,丁酸也成為主要液相末端產物之一,且揮發酸總量顯著高于其他組.
3.3 腐殖酸使優勢菌群從、、、和演替為、和,冗余分析表明產氫量與、、、和呈正相關.
[1] 王海燕,郝瑞霞,趙雅琪,等.蠟樣芽孢桿菌中甲酸脫氫酶基因產氫研究[J]. 中國環境科學, 2018,38(2):729-736. Wang H Y, Hao R X, Zhao Y Q, et al. Studies on hydrogen production via formate dehydrogenase fromstrains [J]. China Environmental Science, 2018,38(2):729-736.
[2] 杜金宇,任學勇,青春耀,等.厭氧混合菌群生物共發酵產氫動力學研究[J]. 農業機械學報, 2019,50(3):294-299. Du J Y, Ren X Y, Qing C Y, et al. Dynamics characteristics of co-fermentative hydrogen production by mixed photosynthentic and anaerobic consortium [J]. Transactions of the Chinese Society for agricultural Machinery, 2019,50(3):294-299.
[3] Zhang Q, Wang Y, Zhang Z, et al. Photo fermentative hydrogen production from crop residue: a mini review [J]. Bioresource Technology, 2017,229:222-230.
[4] Arimi M M, Knodel J, Kiprop A, et al. Strategies for improvement of biohydrogen production from organic rich wastewater: a review [J]. Biomass and Bioenergy, 2015,75:101-118.
[5] 班巧英,劉 琦,余 敏,等.氧化還原介體催化強化污染物厭氧降解研究進展[J]. 科技導報, 2019,37(21):88-96. Ban Q Y, Liu Q, Yu M, et al. Review on catalytic effects of redox mediator in anaerobic degradation of pollutants [J]. Science and Technology Review, 2019,37(21):88-96.
[6] 丁惠君,吳亦瀟,鐘家有,等.兩種介體物質在漆酶降解磺胺類抗生素中的作用 [J]. 中國環境科學, 2016,36(5):1469-1475. Ding H J, Wu Y X, Zhong J Y, et al. Role of two mediators in sulfonamide antibiotics degradation by laccase oxidation system [J]. China Environmental Science, 2016,36(5):1469-1475.
[7] Zhang D, Zhang N, Yu X et al. Effect of humins from different sediments on microbial degradation of 2,2′,4,4′,5,5′- hexachlorobiphenyl (PCB153), and their polyphasic characterization [J]. Rsc Advances, 2017,7(12):6849-6855.
[8] 班巧英,岳立峰,李建政,等.萘厭氧降解菌群的富集及氧化還原介體的強化[J]. 中國環境科學, 2020,40(7):3150-3155.Ban Q Y, Yue L F, Li J Z, et al. Enrichment of naphthalene anaerobic degrading bacterial consortium and enhancement by redox mediators [J]. China Environmental Science, 2020,40(7):3150-3155.
[9] 國家環保局《水和廢水監測分析方法》編委會.水和廢水監測分析方法 [M]. 北京:中國環境科學出版社, 2002:27-30.State Environmental Protection Administration. Water and wastewater monitoring and analysis methods [M]. Beijing: China Environmental Science Press, 2002:27-30.
[10] 張立國,班巧英,李建政.UASB反應器中產甲烷菌對溫度脅迫的響應[J]. 中國環境科學, 2016,36(4):1082-1086. Zhang L G, Ban Q Y, Li J Z. Response of methanogens on temperature stress in an UASB reactor [J]. China Environmental Science, 2016,36(4):1082-1086.
[11] Martinez C M, Alvarez L H, Celis L B et al. Humus-reducing microorganisms and their valuable contribution in environmental processes [J]. Applied Microbiology and Biotechnology, 2013,97(24): 10293-10308.
[12] Abubackar H N, Keskin T, Yazgin O, et al. Biohydrogen production from autoclaved fruit and vegetable wastes by dry fermentation under thermophilic condition [J]. International Journal of hydrogen Energy, 2019,44:18776-18784.
[13] Qin Y, Li L, Wu J, et al. Co-production of biohydrogen and biomethane from food waste and paper waste via recirculated two- phase anaerobic digestion process: Bioenergy yields and metabolic distribution [J]. Bioresource Technology, 2019,276:325-334.
[14] Antwi P, Li J, Boadi P O, et al. Functional bacterial and archaeal diversity revealed by 16S rRNA gene pyrosequencing during potato starch processing wastewater treatment in an UASB [J]. Bioresource Technology, 2017,235:348–357.
[15] Wang J, Ruan C, Song L, et al.sp.nov., a planctomycete isolated from a deep-sea water sample of the Northwest Indian Ocean [J]. International Journal of Systematic and Evolutionary Microbiology, DOI:10.1099/ijsem.0.004301.
[16] Kulichevskaya I S, Ivanova A A, Detkova E N, et al.gen. nov., sp. nov., a stalked planctomycete from a littoral wetland of a boreal lake [J]. International Journal of Systematic and Evolutionary Microbiology 2015,65:1659– 1665.
[17] Yamada T, Imachi H, Ohashi A, et al.gen. nov., sp. nov. and. nov., sp. nov., strictly anaerobic, filamentous bacteria of the phylum Chloroflexi isolated from methanogenic propionate-degrading consortia [J]. International Journal of Systematic and Evolutionary Microbiology, 2007,57(10): 2299–2306.
[18] Nunoura T, Hirai M, Miyazaki M, et al. Isolation and characterization of a thermophilic, obligately anaerobic and heterotrophic marine Chloroflexi bacterium from a Chloroflexi-dominated microbial community associated with a Japanese Shallow Hydrothermal system, and proposal forgen. nov., sp. nov [J]. Microbes and Environments, 2013,28(2):228-235.
[19] Spring S, J?ckel U, Wagner M, et al.gen. nov., sp. nov., a novel facultatively anaerobic, N2O-producing bacterium isolated from activated sludge, and transfer oftogen.nov., comb. nov [J]. International Journal of Systematicand Evolutionary Microbiology, 2004,54:99-106.
[20] Strepis N, Sánchez-Andrea I, van Gelder AH, et al. Description ofilyis sp.nov.by combined physiological and in silicogenome hybridization analyses [J]. International Journal of Systematic and Evolutionary Microbiology, 2016,66(10):3957-3963.
[21] Jabari L, Gannoun H, Cayol J L, et al.gen.nov.,sp.nov., A member of the familyisolated from an upflow anaerobic filter treating abattoir wastewaters [J]. International Journal of Systematic and Evolutionary Microbiology, 2012,62:2522–2527.
[22] Jones J R, Massanet-Nicolau J, Mulder M J J. Increased biohydrogen yields, volatile fatty acid production and substrate utilisation rates via the electrodialysis of a continually fed sucrose fermenter [J]. Bioresource Technology, 2017,229:46-52.
Improving fermentative hydrogen production of anaerobic sludge by redox mediators.
ZHANG Li-guo1,2, AI Bing-ling3, LI Jian-zheng4, BAN Qiao-ying1,2*
(1.College of Environment and Resource Sciences, Shanxi University, Taiyuan 030006, China;2.Shanxi Laboratory for Yellow River, Taiyuan 030006, China;3.Haikou Experimental Station, Chinese Academy Tropical Agricultural Sciences, Haikou 571101, China;4.School of Environment, Harbin Institute of Technology, Harbin 150090, China)., 2021,41(5):2196~2202
To improve the hydrogen production activity of anaerobic sludge, the effects of redox mediators (ROMs) on the hydrogen production efficiency of anaerobic sludge using glucose as substratewere investigated by batch culture. The microbial community structure was revealed by Illumina MiSeq sequencing. The results showed that the cumulative hydrogen production and maximum hydrogen production rate (max) of the control were respectively 11.0mL and 0.28mL/h under conditions of the fermentative volume of100mL and the initial glucose concentration of 500mg/L. When humic acid and anthraquinone-2-sulfonic acid (AQS) as ROMs were added to the system, the hydrogen production capacity of anaerobic sludge was significantly improved. The cumulative hydrogen production was higher than the control by 56.4% and 13.6%, respectively. Whilemaxwas increased by 53.6% and 10.7%, respectively. On the contrary, the hydrogen production capacity of anaerobic sludge was inhibited when graphene oxide (GO), hennaquinone and anthraquinone-2,6-disulfonic acid (AQDS) were used as ROMs. Illumina MiSeq sequencing revealed that the dominant microbial groups from each sample were different.,,,andwere the dominant genera in control. The major genera were shifted to,andin humic acid, AQS and hennaquinone fermentation systems. While,,andbecame the predominant genera in AQDS and GO fermentation systems. Redundancy analysis indicated that hydrogen production was positively correlated with,,,and, whereas negatively correlated with,,and.
anaerobic sludge;redox mediators;hydrogen production by fermentation;microbial community
X703.5
A
1000-6923(2021)05-2196-07
張立國(1980-),男,河南安陽人,副教授,博士,主要從事廢水厭氧生物處理.發表論文30余篇.
2020-09-15
國家自然科學基金資助項目(51708341,51708548);哈爾濱工業大學城市水資源與水環境國家重點實驗室開放基金項目(QA202137)
* 責任作者, 副教授, banqiaoying@163.com
猜你喜歡
工業設計(2022年8期)2022-09-09 07:43:20
軍民兩用技術與產品(2021年10期)2021-03-16 06:05:30
北京測繪(2020年12期)2020-12-29 01:33:58
裝備制造技術(2019年12期)2019-12-25 03:06:46
中國洗滌用品工業(2019年4期)2019-05-11 09:27:34
豬業科學(2018年4期)2018-05-19 02:04:38
家庭影院技術(2017年9期)2017-09-26 03:41:45
浙江農業科學(2016年11期)2016-05-04 04:16:49
化工進展(2015年6期)2015-11-13 00:27:33
無機化學學報(2014年9期)2014-02-28 17:32:57
