黏質沙雷氏菌靈菌紅素合成基因簇異源表達及其潛在的溫度調控機制
2021-05-31 02:30:10劉方晨賈憲波吳良泉林俊杰陳龍軍林陳強陳濟琛
福建農(nóng)業(yè)學報 2021年3期
劉方晨,賈憲波 ,吳良泉,方 宇,趙 恪,林俊杰,陳龍軍,張 慧,林陳強,陳濟琛
(1. 福建農(nóng)林大學資源與環(huán)境學院,福建 福州 350002;2. 福建省農(nóng)業(yè)科學院土壤肥料研究所,福建 福州 350003;3. 福建農(nóng)林大學生命科學學院,福建 福州 350002)
0 引言
【研究意義】靈菌紅素類(Prodiginines)是微生物源的三吡啶類紅色次生代謝產(chǎn)物,具有抗菌、除藻、抗植物病原體以及其他重要的生物活性[1?4],在醫(yī)藥、環(huán)境治理、紡織染料乃至化妝品等領域具有廣闊的應用前景和市場價值。根據(jù)其烷基側鏈的不同分為靈菌紅素(prodigiosin,PG)、間環(huán)丙菌素(metacycloprodigiosin,MPG)、十一烷基靈菌紅素(undecylprodigiosin,UPG)、壬烷基靈菌紅素(nonylprodigiosin,NPG)、環(huán)丙烷靈菌紅素(cycloprodigiosin,CPG)等。其中PG是該類色素的典型代表之一,主要出現(xiàn)在微生物生長的后期[5],是一種暗紅色的粉末,不溶于水,易溶于甲醇、乙醇、丙酮等有機溶劑[6]。作為靈菌紅素家族中的一員,PG主要是沙雷氏菌屬、放線菌屬、假單胞菌屬、海洋類細菌等的次生代謝產(chǎn)物[7],其中黏質沙雷氏菌(Serratia marcescens)所產(chǎn)生的PG生物活性較好,因此被作為生產(chǎn)PG的模式菌株[8]。研究發(fā)現(xiàn),該菌合成靈菌紅素對溫度有較為嚴格的要求,在溫度高于37 ℃以及低于20 ℃時色素合成被完全抑制[9],28 ℃為其生產(chǎn)靈菌紅素的最佳溫度[10],但其受溫度調控的機制尚不明確。【前人研究進展】黏質沙雷氏菌合成PG是由兩個復雜的分叉途徑共同完成的,而這兩個分叉途徑已被闡明。這兩個分支合成了兩個前體物質,分別是2-甲基-3-n-戊基吡咯(MAP)和4-甲氧基-2,2′-雙吡咯-5-甲醛(MBC),再由MAP和MBC的縮合生成PG[11]。靈菌紅素生物合成途徑由pig基因簇基因共同參與,pig基因簇有pigA-pigN共14個編碼基因,其中pigB、pigD和pigE參與了MAP的合成,而pigA、pigF、pigG、pigH、pigI、pigJ、pigK、pigL、pigM和pigN則參與了MBC的合成。PG合成的前體物質主要有脯氨酸、甲硫氨酸、丙氨酸、絲氨酸、組氨酸、二辛烯醛、丙二酰-CoA和乙酰- CoA[12]。靈菌紅素合成基因簇不僅能夠在黏質沙雷氏菌中合成靈菌紅素,也能在其他菌株中異源合成靈菌紅素,Thomson等[13]首次將黏質沙雷氏菌完整的PG生物合成基因簇克隆下來在歐文氏菌(Erwinia carotovora)中成功表達并合成目的產(chǎn)物。【本研究切入點】黏質沙雷氏菌合成靈菌紅素受溫度調控的機制目前尚未被闡明,其對溫度的具體要求不明確,這對靈菌紅素的規(guī)模化生產(chǎn)造成阻礙。因此,從分子水平研究其受溫度調控的機制意義重大。本研究將靈菌紅素合成基因簇與不同類型啟動子相連接后于大腸桿菌中異源表達,探究更換啟動子是否能夠令其在相對高溫(37 ℃)合成色素。【擬解決的關鍵問題】本研究供試菌株黏質沙雷氏菌FZSF02為本課題組前期篩選保藏的靈菌紅素高產(chǎn)菌株[14],為了探究其pig基因簇能否在大腸桿菌中表達并合成PG,并探究其在37 ℃不產(chǎn)靈菌紅素的原因是其pig基因簇啟動子在37 ℃不表達或表達量降低,還是pigA-N編碼合成的靈菌紅素前體所用的蛋白在37 ℃無活性。本研究通過構建3種包含不同啟動子與pig基因簇的重組質粒,分別轉入大腸桿菌中探究其能否在大腸桿菌中合成靈菌紅素,并通過RTqPCR來檢測37 ℃與28 ℃條件下培養(yǎng)的FZSF02菌株pigA、pigF、pigN基因的轉錄水平變化,進而驗證其在37 ℃條件下不能合成PG究竟是在轉錄水平上的調控,還是因為受到合成PG所需要的酶的影響,抑或是受到兩者的綜合調控。
1 材料與方法
1.1 試驗材料
1.1.1 菌株、質粒以及培養(yǎng)條件 本研究供試菌株和質粒詳情見表1,如無特殊注明:大腸桿菌在37 ℃培養(yǎng),黏質沙雷氏菌在28 ℃培養(yǎng),使用LB培養(yǎng)基(酵母粉0.5% w/v、胰蛋白胨1% w/v、氯化鈉1%w/v),固體培養(yǎng)基含2%瓊脂。抗生素如無特殊注明則為:100 mg·L?1ampicillin(Amp),100 mg·L?1K anamycin(Kan)。
1.1.2 主要試劑及儀器 細菌DNA提取試劑盒(百泰克DP2001)PrimeSTAR?GXL DNA Polymerase(R050A)、PrimeSTAR?Max DNA Polymerase(R045A)、TaKaRa MiniBEST DNA Fragment Purification Kit Ver.4.0 DNA片段純化試劑盒(9761)均購自寶日醫(yī)生物技術(北京)有限公司;2X SanTaq PCR Mix(生工B532061)快速質粒小提試劑盒(DP105)、培養(yǎng)細胞/細菌總RNA提取試劑盒(DP430)均購自天根生化科技(北京)有限公司;TransScript?One-Step gDNA Removal and cDNA Synthesis SuperMix試劑盒(AT311-03)、PerfectStart Green qPCR SuperMix試劑盒(TG-AQ601-02)均購自北京全式金生物技術有限公司;PCR擴增儀(ABI-Veriti? 96-Well);實時熒光定量PCR儀(ABI-QuantStudio? 6 Flex);穩(wěn)壓穩(wěn)流電泳儀(DYY-6 B)以及凝膠成像分析系統(tǒng)(uvitec-Essential V4)。

表 1 供試菌株和質粒Table 1 Bacterial strains and plasmids used in this study
1.2 試驗方法
1.2.1 pig基因簇的克隆 利用細菌基因組DNA提取試劑盒從黏質沙雷氏菌FZSF02中提取其基因組DNA,并以提取的FZSF02基因組DNA為模板,以設計的ExpF和ExpR為引物,利用擴增長片段的高保真DNA聚合酶PrimeSTAR?GXL DNA Polymerase進行PCR擴增得到pig基因簇序列;PCR反應條件為:98 ℃變性10 s,68 ℃退火加延伸 10 min,共33個循環(huán)。
1.2.2 E.Coli DH5α(pTOPO-ppPig)重組菌株的構建及鑒定 利用零背景pTOPO-Blunt Simple平末端克隆試劑盒構建重組質粒pTOPO-ppPig,重組質粒的構建所用引物、菌株、質粒分別如表1和表2所示。利用DNA純化試劑盒將上述克隆出的pig基因簇DNA進行純化,根據(jù)pTOPO平末端克隆試劑盒說明書,構建10 μL的克隆反應體系:pTOPO-Blunt Simple Vctor(30 ng·μL?1)1 μL、10×Enhancer 1 μL、pig基因簇DNA純化產(chǎn)物8 μL。室溫反應20 min后,轉入E. coli DH5α感受態(tài)細胞中,冰浴30 min,42 ℃熱激45 s,冰上放置2 min后,加入0.89 mL LB液體培養(yǎng)基,37 ℃搖床孵育2 h;菌體均勻涂布于含有Amp的LB平板上,28 ℃培養(yǎng)過夜,挑取紅色的單菌落,利用M13F和M13R引物進行PCR鑒定,產(chǎn)物送至上海博尚生物技術有限公司測序,利用NCBI B LAST對發(fā)回的結果進行比對分析。
1.2.3 E.coli DH5α(pTOPO-kpPig)重組菌株的構建及鑒定 以實驗室保存的pTOPO-kan質粒利用KanproF2和KanproR2引物進行PCR擴增將質粒線性化,得到一條不含Kan抗性基因的線性化載體。使用無縫克隆試劑盒將純化的pig基因簇DNA與純化的線性化載體相連接,連接體系為10 μL:線性化載體3.5 μL、pig基因簇DNA3.5 μL、酶3 μL。37 ℃靜置30 min后,再次轉入E.coli DH5α感受態(tài)細胞,余下步驟與1.2.2相同。
1.2.4 E.coli BL21(pEASY-Pig)重組菌株的構建及誘導表達 利用pEASY?- Blunt E2 表達試劑盒構建重組質粒pEASY-Pig,克隆反應體系為:pig基因簇DNA 4 μL、pEASY?- Blunt E2 Expression Vctor 1 μL。室溫反應5 min后,轉入E.coli BL21感受態(tài)細胞中,轉化操作與1.2.2相同,挑取數(shù)個長勢良好的單菌落,接種入1.5 mL離心管,孵育至渾濁后,用T7P、T7T引物(見pEASY質粒說明書)進行PCR擴增驗證,瓊脂糖凝膠電泳后條帶在21 000 bp左右的即為陽性克隆子,已驗證為陽性的菌株取少量菌液劃線于加入Amp和IPTG(0.5 mmol·L?1)的LB固體培養(yǎng)基,置于 28 ℃培養(yǎng)箱中過夜培養(yǎng)并誘導其合成靈菌紅素。
1.2.5 比色法測定靈菌紅素 取少量菌體,溶于5 mL酸性甲醇中,靜置15 min后,10 000 r·min?1超速離心5 min,取上清液,利用紫外分光光度計測定其在535 nm吸光度(靈菌紅素酸性條件下在535 nm處 有特征吸收)[14]。

表 2 供試引物Table 2 Primers used in this study
1.2.6 野生型FZSF02菌株在不同溫度條件下(28 ℃/37 ℃)pigA、pigN和pigF基因轉錄水平的RT-qPCR測定 將野生型FZSF02菌株接種于LB液體培養(yǎng)基中,放置于28 ℃培養(yǎng)作為種子液,在OD600值達到1.0~1.5時進行轉接,接種量為1%,37 ℃和28 ℃樣品各設置3個試驗重復共接種6瓶,接種后分別置于28 ℃與37 ℃條件進行培養(yǎng),培養(yǎng)過夜后利用細菌總RNA提取試劑盒提取其總RNA,并反轉錄成cDNA作為RT-qPCR模板。設計FZSF02的16S rRNA基因引物,以及pigA、pigF和pigN基因引物(如表2所示),按照qPCR試劑盒說明書進行反映,檢測各個基因的轉錄水平,以16S rRNA作為內(nèi)參基因,以37 ℃的基因轉錄水平作為對照(CK),每個樣品3次技術重復,計算28 ℃溫度條件下各基因轉錄水平與其的倍數(shù)關系。
2 結果與分析
2.1 3種重組菌株的成功構建
通過構建3種不同啟動子的pig基因簇重組菌株,研究啟動子在溫度調控PG合成的機制中的作用,利用表2中的引物,用長片段高保真DNA聚合酶分別PCR擴增出完整的pig基因簇、完整的pig基因簇和啟動子,其中得到片段大小為20 984 bp、21 390 bp。瓊脂糖凝膠電泳圖如圖1-A所示,1、2泳道依次為pig基因簇、pig基因簇和pig啟動子。
按照方法中所述利用上述片段與相應載體連接構建出3種重組質粒(圖2)并轉入相應的感受態(tài)細胞中,在帶有Amp的LB固體平板上,篩選出長勢良好的重組菌株單菌落。分別利用T7引物與M13引物對pEASY質粒和兩個pTOPO質粒所連接的片段進行PCR驗證,產(chǎn)物進行瓊脂糖凝膠電泳后如圖1-B所示,pTOPO-ppPig、pTOPO-kpPig、pEASY-Pig重組質粒所連片段(泳道1、2、3),3個泳道的條帶均在大于15 000 bp處有明顯條帶,與預期相符。將PCR驗證正確的質粒送樣測序,測序結果與NCBI中菌株FZSF02靈菌紅素合成基因簇進行比對,發(fā)現(xiàn)其匹配度為100%,可知得到的質粒即為目的質粒,測序 陽性質粒按照上述的質粒名稱進行命名。
2.2 pig基因簇在3種重組菌株中均成功表達
經(jīng)驗證為陽性的單菌落擴大培養(yǎng),將其再次劃線于Amp抗性LB平板上,其中pTOPO-ppPig和pTOPOkpPig重組菌株啟動子為組成型啟動子,不需誘導即可啟動基因表達,將其分別在28 ℃與37 ℃條件下培養(yǎng)48 h,觀察轉入重組質粒后的大腸桿菌能否在對應條件下合成靈菌紅素,結果如圖3所示,經(jīng)48 h的培養(yǎng)后,28 ℃培養(yǎng)條件下瓊脂平板上菌株變成紅色,但與野生型FZSF02的產(chǎn)量仍有差距,而37 ℃培養(yǎng)條件下的重組菌株無色素合成。而pEASY-Pig重組菌株經(jīng)鑒定后劃線于帶有Amp抗性和IPTG的LB固體平板上,IPTG可以誘導T7強啟動子表達,該菌株在28 ℃和37 ℃培養(yǎng)條件下均能夠合成PG。通過比色法測定靈菌紅素結果如表3所示,pTOPOppPig和pTOPO-kpPig重組菌株在37 ℃培養(yǎng)條件下535 nm吸光度幾乎為0,pEASY-Pig重組菌株在37 ℃培養(yǎng)條件下535 nm吸光度為0.026,雖然吸光度較小但可以確定該菌株在37 ℃有合成靈菌紅素,且遠小于該菌株28 ℃靈菌紅素合成量。
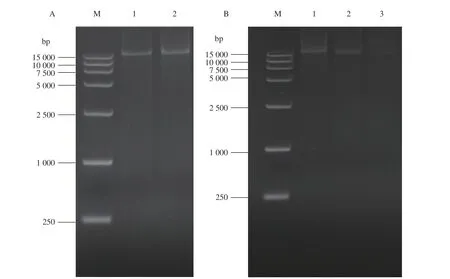
圖 1 pig基因簇、pig基因簇和pig啟動子序列以及3種重組質粒所連片段的驗證PCR擴增結果Fig. 1 PCR amplification results on pig gene cluster, pig gene cluster and pig promoter, and validation on sequences of 3 recombinant plasmid segments
試驗結果表明黏質沙雷氏菌FZSF02的pig基因簇能夠在不同啟動子的作用下在大腸桿菌中轉錄,并合成目的產(chǎn)物靈菌紅素。然而只有攜帶T7啟動子的重組菌株在37 ℃合成少量的靈菌紅素。
2.3 RT-qPCR的檢測
對pigA、pigF、pigN等基因在不同溫度發(fā)酵后轉錄水平的變化進行RT-qPCR檢測,以16S rDNA作為內(nèi)參基因,利用SYBR?Green法實時定量分析在不同培養(yǎng)溫度下(28 ℃與37 ℃)的基因轉錄水平,并用2???CT的方法計算37 ℃培養(yǎng)條件與28 ℃培養(yǎng)條件下的基因轉錄水平的倍數(shù)關系(圖4)。結果顯示,與28 ℃培養(yǎng)條件下相比,37 ℃培養(yǎng)條件下各基因的轉錄水平呈下調趨勢,pigA、pigF和pigN基因分別下調了2倍、3倍、2倍以上。推測在37 ℃培養(yǎng)條件下野生型黏質沙雷氏菌FZSF02不合成PG是因為其pig基因簇中的編碼基因轉錄水平下調,導致其編碼合成的蛋白(酶)的數(shù)量下降,進而影響PG的合成。
3 討論與結論
黏質沙雷氏菌合成PG有嚴苛的溫度要求,一般在24~30 ℃,最適溫度為28 ℃,在37 ℃培養(yǎng)條件下PG的合成受到抑制[15],高溫抑制靈菌紅素合成的機制尚不明了[16]。有可能是調控PG合成的pig基因簇及其操縱子在37 ℃轉錄表達水平下調或不表達,也可能是參與PG前體MAP和MBC合成的某種或幾種酶失去活性。有研究表明,溫度對OmpR家族的CpxR雙組分調控系統(tǒng)以及pig基因簇啟動子有調控作用并影響PG的合成[17]。ERIC等[18]發(fā)現(xiàn)PG生物合成操縱子在37 ℃的轉錄低于30 ℃,并能夠調控色素的合成。Andrea等證明[19]黏質沙雷氏菌調控色素合成的pig基因簇能夠在惡臭假單胞菌KT2440中異源表達,并高效的合成靈菌紅素。本研究應用異源表達和pig基因轉錄水平兩種方案探討?zhàn)べ|沙雷氏菌合成靈菌紅素的溫度依賴性的內(nèi)在機制。
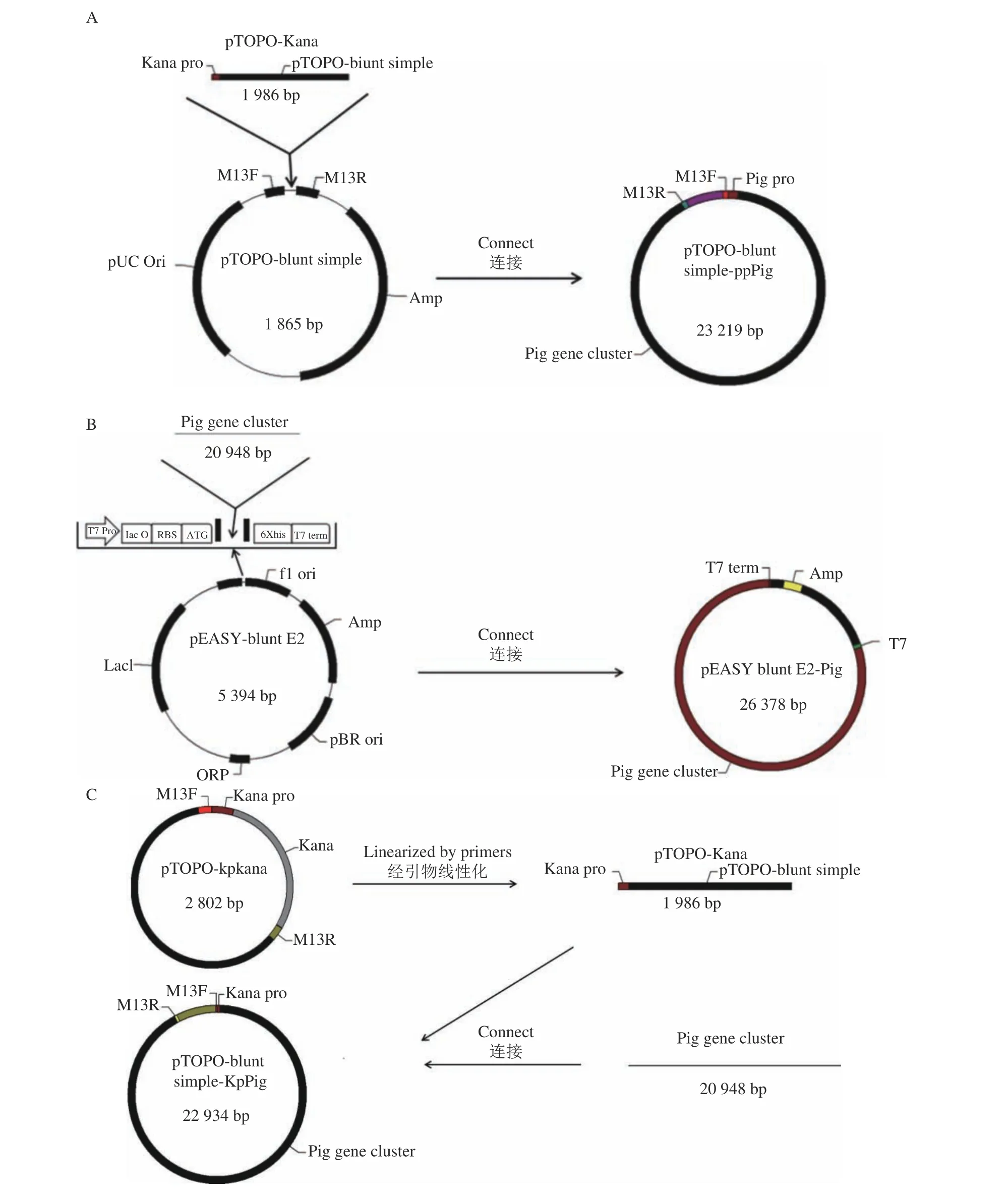
圖 2 3種重組質粒的構建過程Fig. 2 Construction of 3 recombinant plasmids
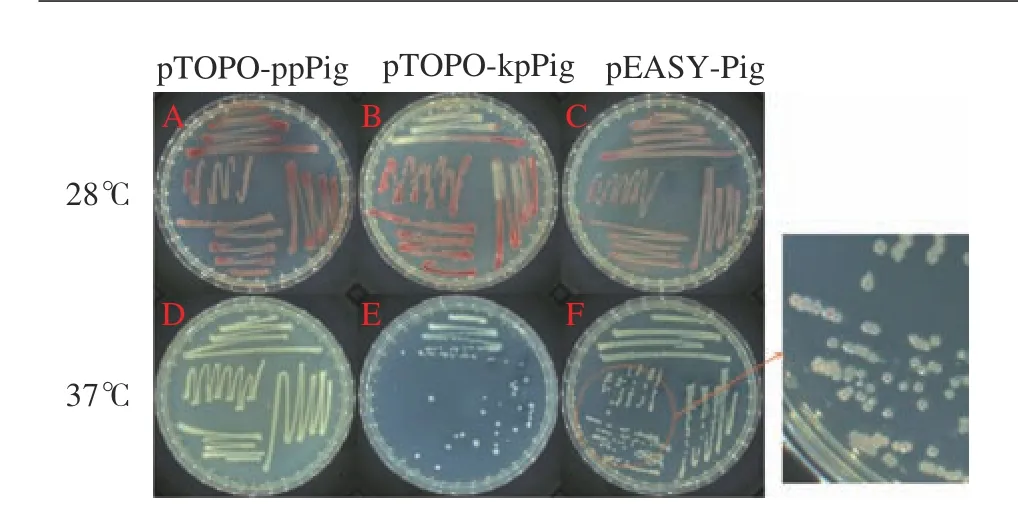
圖 3 3種重組菌株在28 ℃和37 ℃培養(yǎng)條件下靈菌紅素的合成情況Fig. 3 Prodigiosin synthesis of 3 recombinant strains cultured at 28 ℃ and 37 ℃
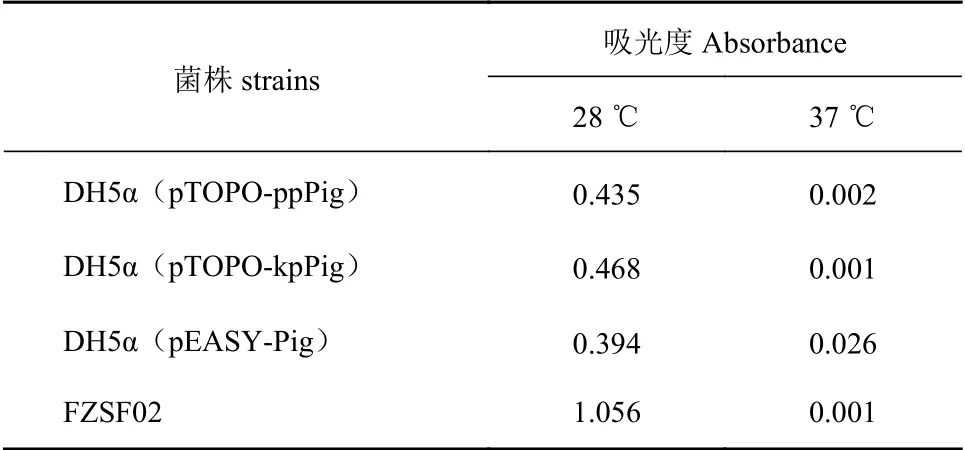
表 3 菌株在535 nm下的吸光度Table 3 Absorbance at 535 nm of recombinant strains
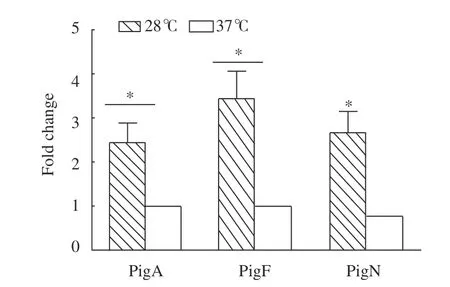
圖 4 粘質沙雷氏菌在不同溫度條件下所選基因的轉錄水平倍數(shù)變化Fig. 4 Expressions of selected genes of S. marcecens at different temperatures
通過構建不受溫度影響的啟動子與pig基因簇連接的重組質粒并在大腸桿菌中異源表達,進而驗證啟動子在溫度調控中的影響,并進一步探究溫度調控PG生物合成可能的機制。首先將FZSF02中的pig基因簇本身的啟動子和pig基因簇一同轉入大腸桿菌,觀察pig基因簇在本身啟動子的作用下能否在大腸桿菌中的表達,結果顯示pig基因簇可以在其本身啟動子的啟動下在大腸桿菌中轉錄表達。隨后利用不受溫度影響的T7啟動子和通用型啟動子(可在37 ℃表達)在大腸桿菌中啟動pig基因簇合成PG,結果顯示3種重組菌株都可以在28 ℃合成PG。其中只有攜帶T7啟動子的重組菌株在37 ℃合成少量的PG,推測是由于T7強啟動子的表達量要遠遠大于pig基因簇本身啟動子和通用型啟動子,這意味著該菌株可以編碼合成更多靈菌紅素前體合成所需要的蛋白(酶),這些蛋白(酶)在37 ℃并不是完全失活,而是活性大大降低了,所以擁有超量蛋白(酶)的pEASY-Pig重組菌株才能在37 ℃合成少量靈菌紅素。而另外兩種菌株則因為合成MAP與MBC的蛋白(酶)在37 ℃條件下失去活性或活性大幅降低而無法合成PG。所以,該試驗證明黏質沙雷氏菌菌株只在低溫下合成靈菌紅素與pig基因簇所編碼的蛋白在37 ℃高溫下活性降低或失活有關。
pigF與pigN基因參與了MBC合成的關鍵步驟,編碼直接或間接參與MBC合成的蛋白(酶):氧甲基轉移酶(PigF)和氧化還原酶(PigN),PigA也參與合成MBC前體物質;它們的轉錄水平反映了MBC的合成水平[9]。本研究利用RT-qPCR分析了FZSF02菌株pig基因簇中的pigA、pigF和pigN基因在37 ℃與28 ℃的轉錄水平差異,發(fā)現(xiàn)在37 ℃培養(yǎng)條件下上述3種基因的轉錄水平相對于28 ℃出現(xiàn)不同程度的下調,這與徐紅等[20]的研究是一致的。上述3個基因轉錄水平分別下調了約59%、69.8%以及61.8%,導致其編碼合成參與靈菌紅素前體合成的蛋白(酶)減少,進而MBC的合成量減少,最終導致靈菌紅素合成量減少。且FZSF02菌株在28 ℃培養(yǎng)條件下色素產(chǎn)量較高,37 ℃培養(yǎng)條件下幾乎沒有色素合成(表3)。該試驗說明在黏質沙雷氏菌FZSF02菌株中pig基因簇上的相關基因除其翻譯的蛋白活性受溫度影響外,基因簇本身的轉錄水平也受溫度控制,即低溫有利于啟動子啟動該基因簇的轉錄。
綜上,初步推測黏質沙雷氏菌FZSF02不能在37 ℃合成PG,一方面是由于pig基因簇在37 ℃轉錄水平下調,導致其編碼的合成MBC與MAP所需要的酶量減少;另一方面是因為37 ℃嚴重影響相關酶的活性。所以,溫度影響?zhàn)べ|沙雷氏菌合成PG的內(nèi)在機制是綜合性的。
本研究通過重組菌株成功異源表達pig基因簇并合成靈菌紅素,給出了黏質沙雷氏菌FZSF02在37 ℃合成PG受到抑制的原因及相關證據(jù),增進了對黏質沙雷菌合成PG的認識,為規(guī)模化發(fā)酵生產(chǎn)PG提供了一定理論依據(jù)。
