
氣候變暖與富營養化交互作用對淺水湖泊水-氣界面N2O通量的影響
2021-06-02 08:27:18胡博文石彭蘭
水生生物學報 2021年3期
胡博文 潘 萌 石彭蘭 徐 軍 張 敏
(1. 華中農業大學水產學院, 池塘健康養殖湖北省工程實驗室, 教育部長江經濟帶大宗水生生物產業綠色發展工程研究中心,武漢 430070; 2. 中國科學院水生生物研究所, 武漢 430072)
溫室氣體(Greenhouse gases)的大量排放與近年來的全球變暖密切相關[1], N2O作為一種重要的溫室氣體在大氣中的濃度以年均(0.25±0.05)%的速度穩定增長[2], 估算對全球變暖的貢獻率已達到6%以上[3], 排在CO2和CH4之后, 位列第三。現有的研究認為, 淡水生態系統中生成的CO2、CH4和N2O主要通過水體和沉積物進一步參與到生物地球化學循環過程[4]。全球變暖不僅會導致平均氣溫的升高, 還會使熱浪(Heat waves, HW)等極端天氣事件的發生頻率顯著增加[5—7]。這些事件將導致淡水生態系統尤其是淺水湖泊的理化性狀及其功能特征受到強烈影響。近年來人類活動對湖泊的干擾加劇, 外源性的氮、磷負荷顯著增加, 致使富營養化趨勢逐漸明顯, 進而對湖泊中氮的循環及N2O排放產生重要影響[8]。
國內外對于湖泊N2O交換過程的研究已有較多報道, 湖泊通常被認為是N2O的潛在排放源。Kortelainen等[9]估算芬蘭及北方湖泊N2O的排放量分別達到0.6和29 Gg N2O-N/y, 表明N2O在這些區域具備顯著的全球變暖潛力(Global warming potential)。國內的許多研究如太湖[10]、白洋淀[11]和鄱陽湖[12]的N2O交換過程在監測的絕大部分時間內也是以排放現象為主。盡管湖泊N2O的排放趨勢明顯, 但在不同區域的響應程度也存在差別。謝燕紅等[13]對內陸水體479個湖泊N2O通量的緯度和營養狀況的分析結果表明, N2O通量呈現隨緯度升高而降低, 隨營養水平升高而升高的變化特征。因此,區域之間的溫差及營養水平的差異均會對湖泊N2O的交換產生顯著的影響。區域性湖泊N2O通量表現出與溫度及營養水平之間的相關性, 但其內在機制仍需通過控制實驗進一步探究。現有的研究主要集中在溫帶地區, 也解釋了該區域影響N2O排放規律的內在聯系, 如Davidson等[4]在長期運行的控制實驗中, 通過設置升溫和營養鹽添加的交互作用觀測溫室氣體交換過程的響應變化, 發現高濃度的氮磷營養鹽添加顯著增加了系統內N2O的排放量, 但升溫的直接影響卻并不顯著; Audet等[14]則在升溫與營養鹽添加的基礎上進行了模擬夏季熱浪的控制實驗, 其結果表明N2O的排放對營養水平的響應不顯著, 而熱浪則顯著促進了夏季N2O排放量的增長。
湖泊中N2O的產生主要依賴于水體與沉積物的微生物過程[15,16], 主要包括硝化、反硝化、硝化-反硝化耦合和硝酸鹽異化還原為銨等作用[17,18]。一般認為硝化作用的最適宜溫度為25—30℃, 反硝化作用的最適宜溫度在30℃以上[19]。水溫的適度升高能夠增強沉積物界面反硝化菌群的活性, Silvennoinen等[20]對不同溶氧與溫度條件下監測沉積物界面反硝化作用的研究發現升溫能夠顯著加快反硝化的速率并且有利于反應的完全進行生成大量N2而降低了產物中N2O與N2的比值。水體中的氮營養元素及其濃度變化對N2O產生所起到的作用已有較多研究, 氮營養元素增加通常能顯著促進N2O的排放[21]。磷營養元素的增加導致水體富營養化, 進而影響N2O的產生過程, 但是否存在直接作用仍需進一步探討。根據現有的研究推測, 在升溫情景下, 水域環境中理化因子和生物因子的積極響應能夠顯著影響N2O微生物過程, 進而引起水-氣界面間N2O交換量的變化。例如水體中的溶氧狀況對氮素轉化途徑的影響起到直接作用[22]。水溫的升高能夠促進沉積物中內源性磷的釋放[23], 并且隨著外源性營養物質的過量輸入, 湖泊暴發水華的風險將顯著上升。藻類的過度生長將消耗掉水體中大量的溶氧, 同時若存在有機碳的外源輸入, 將影響氧氣的消耗速率, 進而為水體創造出更適宜于N2O產生的厭氧微環境[24], 促使富營養化湖泊成為氣候變化的潛在驅動者。
我國的富營養化淺水湖泊大部分集中于長江中下游地區[25], 未來氣候變化趨勢下長江中下游地區典型淺水湖泊生態系統中N2O的交換過程將會發生怎樣的變化, 值得密切關注。中尺度圍隔是一種適合模擬溫度和營養物質的變化如何影響淺水湖泊生態系統在長期和不同營養水平下各種物質動態變化的研究方法[26], 適宜于進一步的探討湖泊生態系統的內在響應機制。本研究以中尺度圍隔(Mesocosm)控制系統模擬21世紀末期長江中下游地區典型淺水湖泊變暖(恒定升溫及波動升溫)與富營養化情景, 旨在評價水體營養水平升高、季節性溫度變化及不同變暖模式對N2O通量的影響, 探討氣候變化與富營養化的交互作用下淺水湖泊水-氣界面N2O交換的響應機制。
1 材料與方法
1.1 實驗地點與系統構建
中尺度圍隔控制系統位于湖北省武漢市華中農業大學水產養殖國家級實驗教學示范中心, 48個相互獨立的塑料水缸(直徑=1.5 m, 高度=1.4 m)均勻分布。控制系統通過3種相互聯系的組件實現加熱功能的穩定運行: 每個圍隔中都包含2個均勻放置的溫度傳感器和1個固定功率的加熱元件(600 W),及一臺全天運行的混合潛水泵。同時, 控制系統通過與遠程電腦的連接實現溫度實時觀測與每日溫度數據云端自動上傳的功能。本實驗于2017年10月期間在梁子湖敞水區采集表層沉積物, 充分混合均勻后加入圍隔系統內, 鋪設沉積物的厚度為10 cm, 并注入深度為1 m的梁子湖水。控制系統內加入湖水的初始TN濃度為0.56 mg/L, 初始TP濃度為0.065 mg/L。在準備工作結束后, 待系統穩定2個月后開始實驗, 本實驗始于2018年1月1日, 止于2018年12月31日。
實驗設計包括3種溫度模式與2種營養鹽模式的處理, 交互后共計6個處理組, 每個處理組6個平行, 利用隨機數抽取器將實驗的處理隨機分配至36個中尺度圍隔中。6個處理組分別表示為: 對照組(C)自然水溫, 在溫度和營養水平方面模擬梁子湖的現有狀態; 固定升溫組(T)依據RCP8.5情景, 模擬本世紀末期的升溫情況[3], 設置恒定升溫4℃; 波動升溫組(V)在固定升溫的基礎上考慮溫度波動的情形, 根據多項研究預測的極端氣候事件持續時間和頻率[3,27—30], 總共設定15次升溫趨勢的波動, 持續時間在2—12d, 以及17次降溫趨勢的波動, 持續時間在3—11d, 最終保持該處理總熱量輸入與恒定升溫處理一致; 磷添加組(P)是在自然水溫條件下每2周向水體中加入磷酸二氫鉀(KH2PO4) 25 μg/L(磷的添加量和頻率根據高偉等[31]及黃文鈺和許朋柱[32]的研究結果并進行適當的調整, 用以模擬梁子湖的富營養化進程); 固定升溫添加磷組(TP)是在自然水溫的基礎上恒定升溫 4℃, 并且每2周向水體中加入KH2PO425 μg/L; 波動升溫添加磷組(VP)是在恒定升溫 4℃同時考慮溫度波動情景, 同時每2周向水體中加入KH2PO425 μg/L。
1.2 樣品采集與測定
實驗期間, 1—2月每2周1次、其余月份每周1次的頻率在控制系統中進行樣品采集及水體理化指標進行現場監測。pH、電導率和溶解氧(Dissolved Oxygen, DO)等參數由HQ40d多功能水質分析儀(HACH, 美國)進行現場測定。利用有機玻璃柱狀采水器(長度=1 m, 直徑=70 mm)取得混合水樣, 過濾水樣通過Whatman GF/C濾膜抽濾后獲得。水體中的TN、TP、硝態氮和氨態氮參照《水和廢水監測分析方法(第四版)》[33]進行測定, 水體葉綠素a(Chlorophylla, Chl.a)的測定方法參照楊玉珍等[34]對“熱乙醇”法的優化。溶解性有機碳(Dissolved Organic Carbon, DOC)由Vario TOC總有機碳分析儀(Elementar, 德國)測定。浮葉植物和沉水植物的生物量分別進行測定,浮葉植物直接估算其蓋度, 沉水植物則估算其浸水體積(Plant volume inhabited, PVI), 具體計算公式為“PVI指數=植物蓋度×植物高度/水深”[35], 以此來反映2種類型水生植物的生物量特征。
N2O交換通量采用漂浮箱-氣相色譜法進行分析測定。漂浮箱箱體為聚乙烯材質的一端開口圓桶(內徑=45 cm, 總高度=50 cm), 箱體外部固定有質量均勻的浮圈, 并通過固定高度使箱體的下部浸沒在水面以下約5 cm, 以此確保采樣期間箱體下部的密閉性, 漂浮箱的工作原件設有氣壓平衡管、采樣管和溫度探頭, 箱體外部均用鋁箔完全覆蓋以反射陽光并盡量減少內部升溫。實驗期間, 每月中下旬選取天氣晴朗的一日于8:30—11:30完成氣體的采集工作, 具體操作步驟為每一批次氣體采集前將漂浮箱橫置3min使其與周圍空氣充分混勻, 隨后將其平穩放置于水面并待水面靜止, 于 2min、14min和26min抽取氣體樣品共3次。第一次取樣時先進行2次排空, 隨后再將30 mL注射器注滿, 余下2次抽取氣體樣品僅需進行一次排空。待完成氣體樣品采集后, 于48h內完成N2O氣體樣品的測定。具體的操作方法為: 使用帶有三通閥的 30 mL醫用注射器將10 mL氣體樣品注入至CA-6手動氣體進樣儀并連用GC 7890氣相色譜工作系統(Agilent, 美國)完成測定工作, N2O濃度由電子捕獲檢測器(ECD)測得, 工作溫度設置為300℃, 利用兩根內徑為2 mm, 長度分別為1和3 m的 80—100目的Porapak Q不銹鋼分離柱完成對N2O的分離和反吹,柱箱溫度55℃, 載氣為高純 N2(純度>99.999%), 流量為25 cm3/min。
通過計算可得到氣體濃度隨時間的變化速率(回歸曲線的斜率,為確保良好的線性關系, 僅選取R2≥0.90的實驗數據)。交換通量公式如下所示:
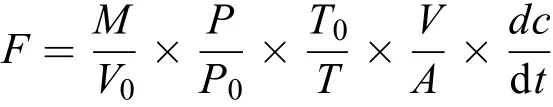
式中,F為N2O交換通量,M為N2O的摩爾質量44 g/mol,P0和T0是標準狀態下的大氣壓力和溫度,V0為目標化合物在標準狀態下的摩爾體積,V/A在規則箱體中轉化為水面上方的有效箱高,P和T為采樣時箱內的實際氣壓和氣溫, dc/dt為漂浮箱體內N2O氣體濃度隨時間變化的斜率(ppm/h)。
通過對實驗期間每一個處理組內的月均N2O交換通量數據進行加法運算, 以此表示N2O年累積交換量。若年累積交換量為負值則處理組在全年尺度上表現為大氣N2O的匯, 而年累積交換量為正值則處理組在全年尺度上表現為大氣N2O的源。年累積交換量公式如下所示:
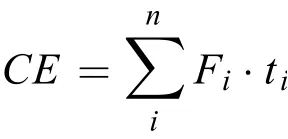
式中,CE(Cumulative exchange)表示N2O的年累積交換量g/(m2·y),n為全年總的采樣次數,Fi表示為第i月的平均N2O交換通量,ti表示為第i月的天數。
1.3 數據分析
實驗數據依據季節性的變化特征將整個實驗周期具體劃分為冬季12月和1—2月, 春季3—5月,夏季6—8月和秋季9—11月。對所有的環境因子進行月均值的處理, 對應于N2O交換通量的月度數據。統計分析均采用R軟件(version 3.6.1)完成。通過對實驗數據的正態性及方差齊次性檢驗發現, 大部分不滿足正態分布與方差齊次性的條件, 進行數據轉化后仍有大部分數據無法滿足檢驗條件, 所以選擇多個獨立樣本的非參數檢驗方法(Kruskal-Wallis 秩和檢驗)檢測處理組間參數的顯著性, 測試不同時期的處理組間對控制系統水-氣界面N2O交換通量的影響, 并通過Spearman相關分析篩選顯著影響于水-氣界面N2O交換的環境因子并探究其影響機制。所有的實驗數據的繪圖工作均通過GraphPad Prism軟件(version 8.0.2)制作完成。最終處理的實驗數據全部以均值±標準誤的結果表示,統計性檢驗以P<0.05標注顯著差異。
2 結果
2.1 控制系統內環境因子的變化特征
水溫實驗期間, 控制系統內不同處理組的日均水溫變化趨勢如圖 1所示。控制系統內T組和TP組的水溫在整個實驗期間恒定高于自然水溫(C組和P組的水溫)4℃左右, 而波動升溫的V組和VP組水溫則在固定升溫4℃的基礎上表現出0—8℃的波動變化。
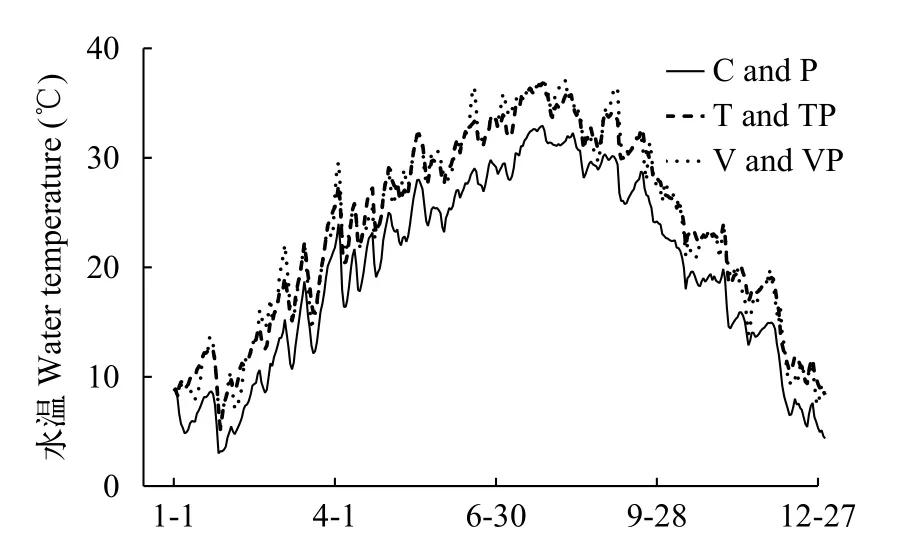
圖 1 實驗期間控制系統內不同處理組的日均水溫變化趨勢Fig. 1 Daily average water temperature in different treatments in mesocosms during the whole experiment period
季節間的水溫變化趨勢明顯, 常溫的處理組(C和P)、固定升溫的處理組(T和TP)和波動升溫的處理組(V和VP)的冬季均溫分別為(7.45±0.28)℃、(11.43±0.28)℃和(11.16±0.33)℃, 春季均溫分別為(20.16±0.47)℃、(24.19±0.48)℃和(24.27±0.48)℃,夏季均溫分別為(29.55±0.22)℃、(33.31±0.21)℃和(33.71±0.25)℃, 秋季均溫分別為(20.86±0.54)℃、(24.81±0.53)℃和(24.63±0.60)℃, 水溫總體表現出冬季低、夏季高和春秋兩季變化快的特征(圖 1)。
電導率、溶解氧和pH控制系統實驗期間電導率(圖 2A)的變化范圍為171.03—426.50 μS/cm,冬季的水體電導率高, 春季則下降至實驗期間的最低水平, 而在夏、秋季又呈現出逐漸回升的趨勢。水體中DO(圖 2B)的波動變化相對強烈, 處理組間的變化保持一致, DO在冬季的濃度均值為(11.50±0.37) mg/L、春季的濃度均值為(9.79±0.16) mg/L、夏季的濃度均值為(4.59±0.31) mg/L和秋季的濃度均值為(3.37±0.28) mg/L。水體pH(圖 2C)在實驗期間的變化范圍在6.88—10.37, 總體上實驗期間不同處理組間pH均表現出冬春季上升、夏秋季下降的趨勢。
碳、氮、磷含量不同處理水體中DOC含量(圖 2D)整體呈現春、夏季上升, 隨后秋、冬季下降的趨勢, 于夏季到達峰值。統計結果表明整個實驗期間內處理組間的DOC含量存在顯著差異(P<0.01), 且C組和P組長期處于較低的水平。水體中的TN(圖 2E)在整個實驗期間持續表現出強烈的波動性, 其變化范圍在0.19—2.16 mg/L, 而季節性的變化趨勢并不明顯(P>0.05)。TP(圖 2F)的變化范圍在0.006—0.352 mg/L, 總體上呈現為隨冬、春和夏季的演替逐漸升高而后在秋季下降的變化特征。水體中(圖 2G)含量有著明顯的季節性變化特征, 冬季的含量最高, 春季相對較低,夏、秋季節緩慢升高, 且整個實驗期間處理組間的含量顯著不同(P<0.01), 其中P組的N含量顯著低于T組(P<0.05)和V組(P<0.05)。水體中(圖 2H)的含量隨季節交替呈現出逐漸升高的變化趨勢, 但在整個實驗周期內處理組間的含量無顯著差異(P>0.05)。
藻類與水生植物的變化在整個實驗期間,控制系統中處理組間的水體Chl.a含量(圖 2I)在夏秋季節均顯著升高, 同時C組的Chl.a含量顯著低于T組(P<0.05)、P組(P<0.05)和VP組(P<0.01)。系統中沉水植物與浮葉植物的季節性變化特征如表 1所示, 沉水植物的優勢種為菹草(Potamogeton crispus),主要出現于冬、春季節, 沉水植物PVI指數(冬春)的變化范圍為0—91.29%, 均值為(44.69±2.25)%, 處理組間在冬季的沉水植物PVI指數顯著不同(P<0.01),表現為C和P組低于T、TP、V和VP組; 而在春季,處理組間沉水植物PVI指數無顯著差異(P>0.05)。此外, 系統中的浮葉植物主要為菱(Trapa bispinosaRoxb.)和芡實(Euryale ferox), 生長期集中于夏秋2個季節, 浮葉植物蓋度(夏秋)為0—98.33%, 均值為(25.35±2.15)%, 處理組間在夏、秋季的浮葉植物蓋度均表現為P組>C組>T組>V組>TP組>VP組。
2.2 控制系統內處理組間N2O交換通量的變化特征
N2O交換通量的季節性變化十分顯著。冬季各處理組均表現為N2O吸收, 控制系統內所有的處理組均表現為N2O的匯; 春季N2O交換通量與全年的變化趨勢一致, C組、T組和V組呈現出N2O從水體向大氣排放, 而P組、TP和VP組則呈現為水體吸收大氣中的N2O, 其中T組的N2O交換通量顯著高于P組(P<0.05)和TP組(P<0.05); 夏季期間N2O在水-氣界面間的交換過程更為復雜, T和TP組表現為水體向大氣吸收N2O的狀態, 而C、V、P和VP組則表現為水體向大氣排放N2O的狀態; 秋季控制系統內(特別是升溫的T組和V組)水-氣界面間N2O的交換則表現出十分強烈的排放現象(圖 3)。
統計檢驗表明(表 2), V組的全年平均N2O交換通量顯著高于添加了磷營養鹽的P組(P<0.01)、TP組(P<0.05)和VP組(P<0.05), 表現出添加磷營養鹽之后控制系統內水-氣界面N2O的排放量顯著減少,這與水-氣界面N2O年累積交換量得到的結果相一致(圖 4), C、T和V組的N2O年累積交換量分別為0.0039、0.0094和0.0200 g/(m2·y), 而P組、TP組和VP組的N2O年累積交換量分別為–0.0049、–0.0065和–0.0018 g/(m2·y), 磷營養鹽的添加促進控制系統內水-氣界面間N2O由排放向吸收的轉換。
2.3 控制系統內處理組間N2O交換通量與不同環境因子的相關性分析
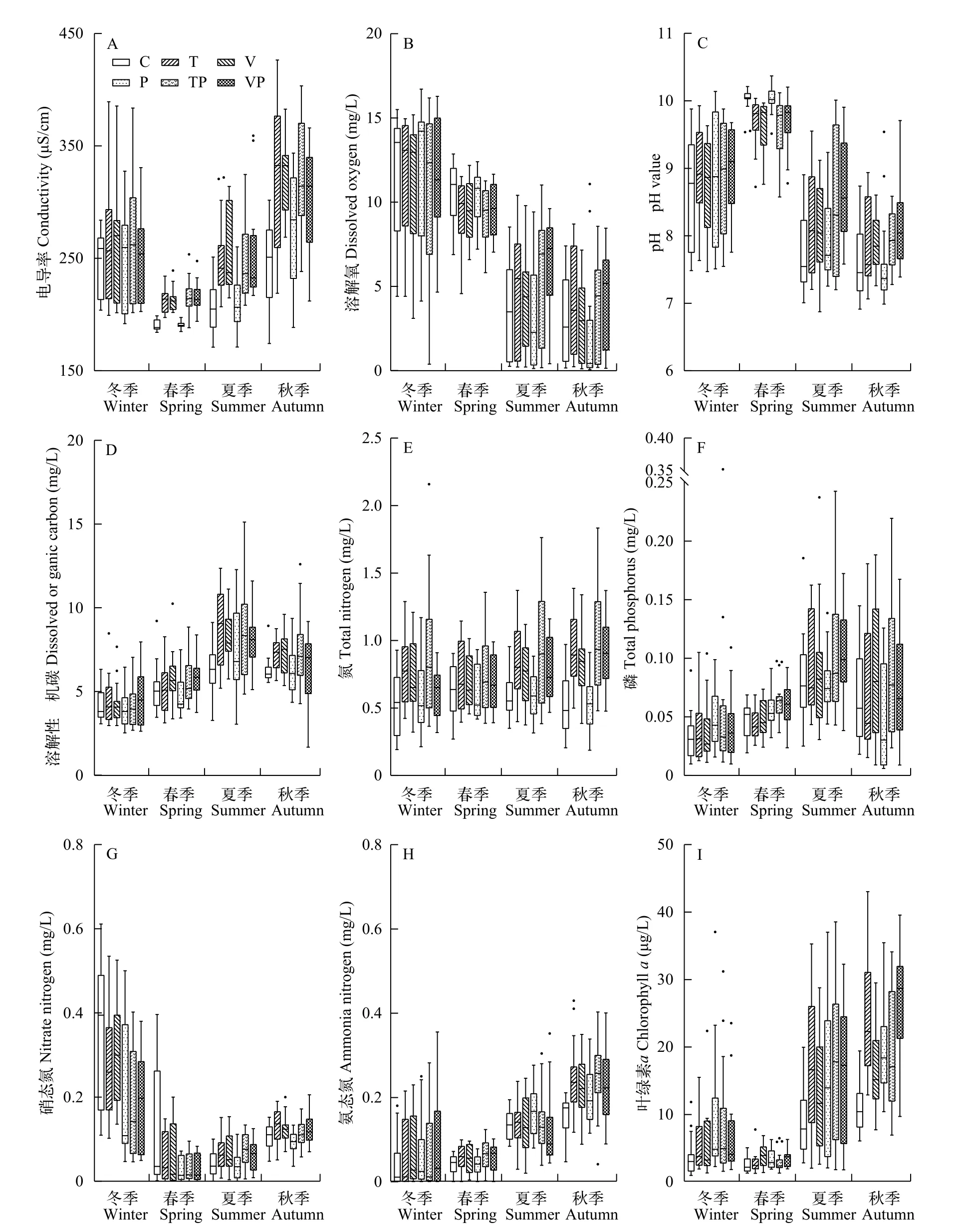
圖 2 控制系統內處理組間環境因子的季節性變化特征Fig. 2 Seasonal variation of environmental factors between experimental treatments in the mesocosms
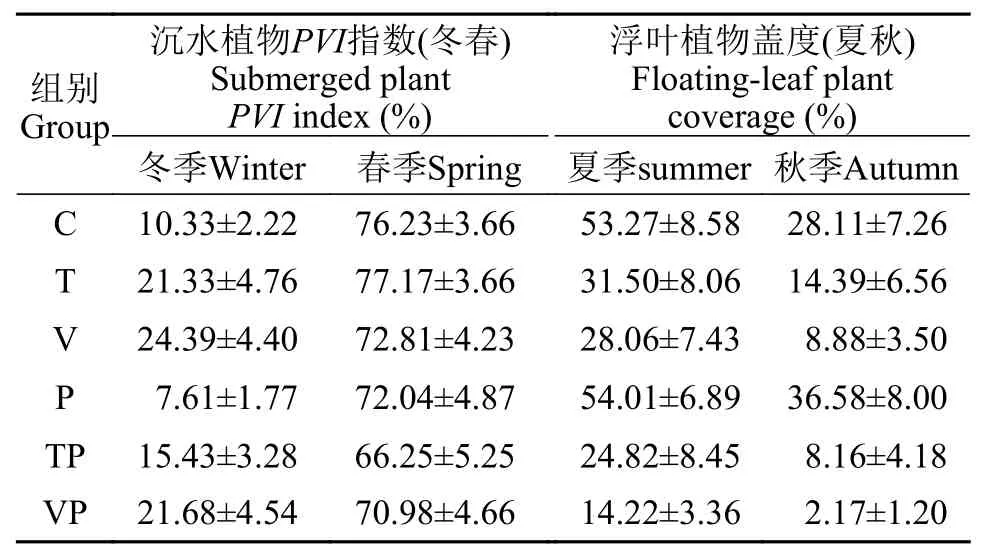
表 1 實驗期間控制系統內不同處理的沉水植物PVI指數和浮葉植物蓋度的季節性變化特征(均值±標準誤)Tab. 1 Submerged plant PVI index and floating-leaf plant coverage of each treatment in mesocosms during the different seasons (Mean±SD)
3 討論
3.1 升溫對水-氣界面N2O交換的影響
現有的研究表明, 淺水泊生態系統中N2O的生成與排放過程均會受到水溫升高的顯著影響。例如當水溫高于5℃時, 沉積物界面的反硝化菌群活性與自然水溫表現出顯著的正相關性[36]。水體中N2O的飽和溶解度也會隨著溫度的升高而降低, 這可能有利于水-氣界面N2O排放量的增加[37]。本研究的結果顯示, 處理組間N2O交換通量的季節性變化十分顯著, 6個處理組在冬季均表現為N2O由水體向大氣的吸收, 而其他季節(尤其是T和V組)則主要表現為N2O由水體向大氣的釋放。統計檢驗表明(表 2), 整個實驗期間V組的年均N2O交換通量顯著高于添加磷營養鹽的各處理組, 而恒定升溫與波動升溫處理中水-氣界面N2O年累積交換量也相對較高。這種效應與代謝理論具有一致性, 代謝理論認為溫度升高與溫室氣體排放速率的增加有顯著相關性[38,39]。Soued等[40]對加拿大魁北克地區不同類型水體的N2O交換通量進行了監測并結合全球區域性湖、庫的研究結果發現, 總體上熱帶地區的年N2O排放量最高, 分別是溫帶地區的10倍和寒帶地區的23倍, 表明溫度是影響N2O排放的一個重要因素。對北歐地區的湖泊[8,41]、我國太湖[42]及三峽水庫[43]等水體N2O季節性排放規律的相關研究也發現, N2O的排放量在平均水溫更高的夏、秋季要高于處于全年平均水溫最低時候的冬季。在近期的一項控制實驗中也得到類似結果, Audet等[14]在長期運行的控制系統中發現模擬夏季熱浪會顯著促進溫室氣體(尤其是CH4和N2O)的排放。進一步說明水溫的升高通常能夠對湖泊水體N2O的排放起到促進作用。
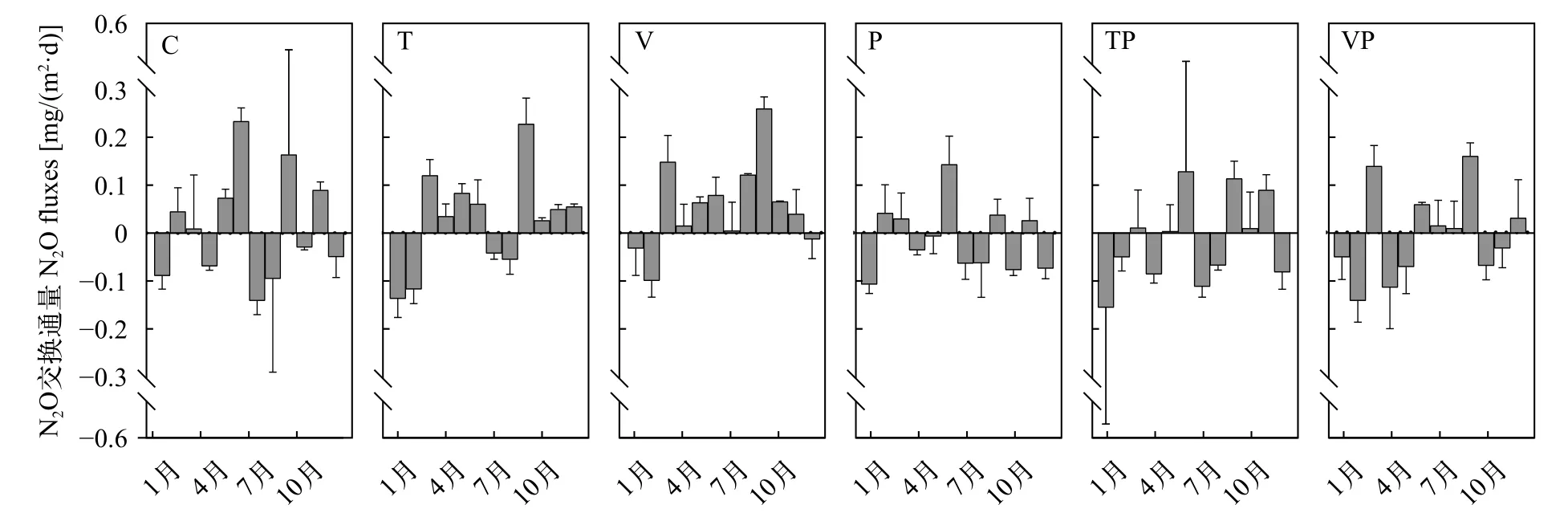
圖 3 控制系統內不同處理N2O交換通量的月度變化特征Fig. 3 Monthly N2O fluxes of each treatment in the mesocosms
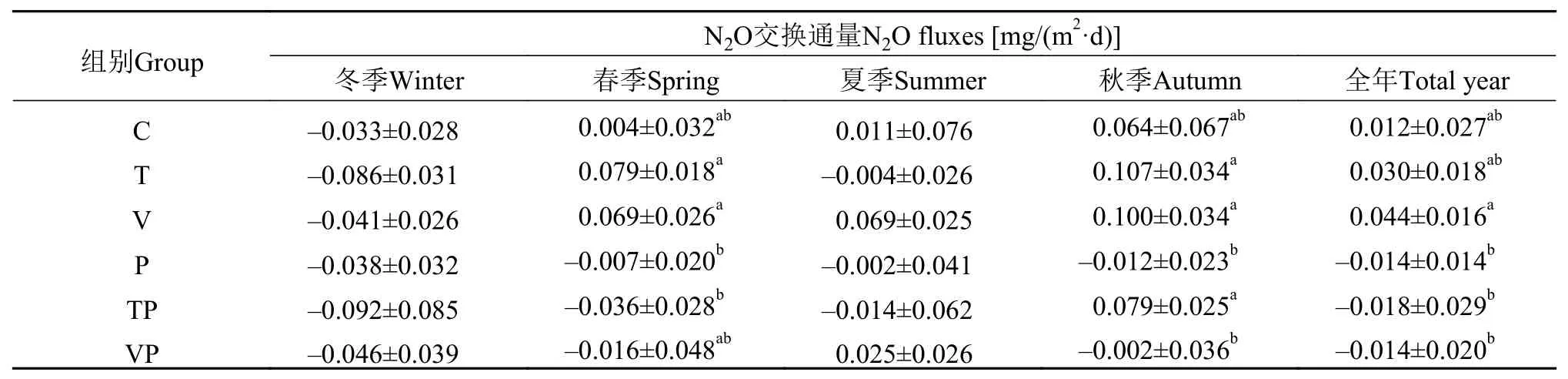
表 2 控制系統內不同季節N2O交換通量的組間變化特征(均值±標準誤)Tab. 2 N2O fluxes of each treatment in mesocosms during the different experiment periods (Mean±SD)
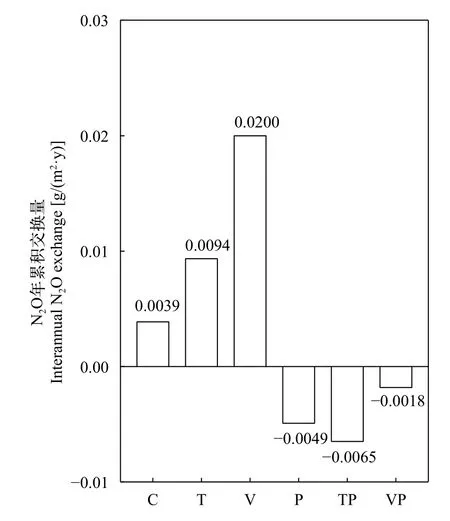
圖 4 控制系統中不同處理水-氣界面間N2O年累積交換量Fig. 4 Total annual N2O flux in water-air interface of different treatments in the mesocosms
3.2 磷添加對水-氣界面N2O交換的影響
有研究表明, 富營養鈣質湖泊的N2O含量與CH4含量分布特征類似, 均呈較高的濃度水平[44],表明湖泊營養狀態與溫室氣體濃度關系密切, 進一步說明富營養化可能導致溫室氣體排放量的增加, 而在本研究中所得到的結果與其不同, 表現為磷營養鹽的添加反而引起了N2O排放量的降低。究其原因, 磷添加對于N2O交換過程的影響仍存在許多不確定性。N2O的產生首要受到水體中氮循環的限制[45], 但有關氮素循環中產生N2O的過程受到的影響因素十分復雜, 其中與本研究相關的主要討論磷添加對于湖泊水體N2O產生速率和排放量的結果仍有待進一步補充。而現有的陸地生態系統中磷元素添加對N2O排放影響的研究結果表明, 磷驅動N2O排放減少的主要機制是由于磷的高有效性使植物增強了對土壤中氮的吸收, 降低了硝化細菌和反硝化細菌反應過程中氮的有效性,從而抑制了N2O的排放[46]。此外, Zheng等[47]的研究發現在熱帶森林中長期增加磷元素可能會使N2O的排放減少。盡管水域環境與陸地之間存在極大的差異性, 但仍可提供一定的參照, 依據現有的研究推測可能是由于磷營養鹽的增加對水生植物與浮游植物生長有促進作用, 初級生產者的大量生長會進一步增強對氮的利用, 導致水體中氮營養元素水平降低, 從而降低了控制系統內水-氣界面間N2O的交換量。
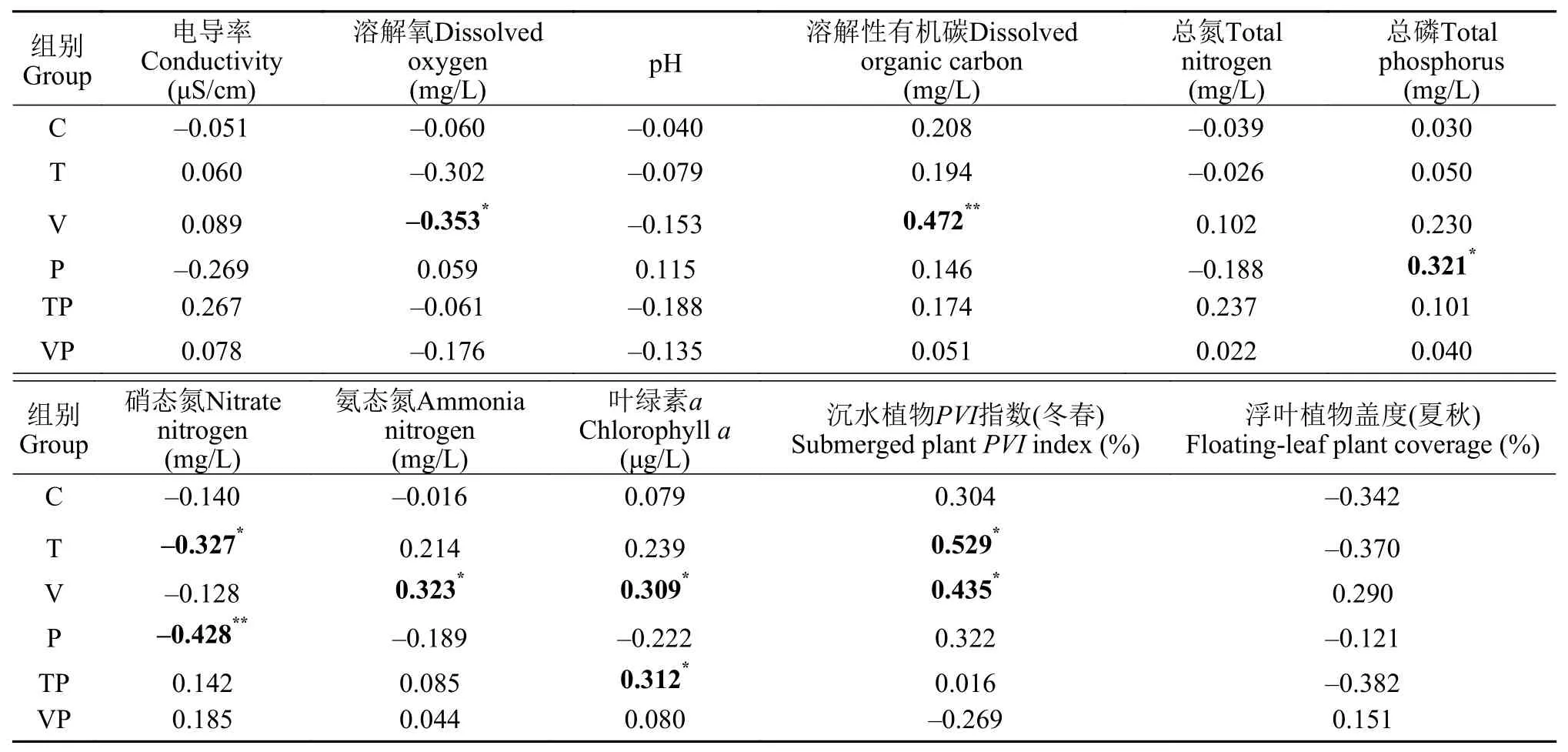
表 3 整個實驗期間不同處理組的N2O交換通量與環境因子的Spearman相關性分析Tab. 3 Spearman correlation analysis of N2O fluxes and environmental factors during the whole experiment period
3.3 升溫與磷添加交互作用對水-氣界面N2O交換的影響
統計檢驗的結果表明, 升溫與磷添加顯著促進了整個實驗期間電導率、pH、DOC和TN含量的升高, 而顯著抑制了浮葉植物的生長。本研究結果表明, N2O生成與消耗的過程與溶解氧、有機物含量、pH、溫度和無機氮可利用性等多重環境因素的調節密切相關[48]。本研究中控制系統水-氣界面N2O交換通量與升溫與磷添加交互作用下的DO、和沉水植物PVI指數(冬春)等環境因子具有顯著的相關性。不同的環境因子在控制系統內受到升溫與磷添加的交互作用而表征出十分顯著的變化。同時, 環境因子對于溫度和磷營養鹽的迅速響應將進一步改變湖泊水體N2O的交換過程。
Kortelainen等[9]在芬蘭87個湖泊中發現N2O含量在pH為6.5左右時最高, 這反映了水體中N2O累積的適宜pH。且有研究表明N2O還原酶更適宜在較高的pH環境中生存[49], 因此在偏堿性的水體中可能更易于消耗掉N2O。在本研究中, 升溫與磷添加交互作用的影響下絕大多數時間水體pH表現為中偏堿性狀態, 水體并未處于N2O生成過程中最適宜的pH范圍之內, 這很可能導致水體中N2O的生成過程受到抑制。
DO是平衡水體中硝化和反硝化反應的重要環境因子, 影響水體中N2O的產生過程[50], 有研究表明湖泊生態系統中的N2O排放通量與水深和DO濃度具有顯著的相關關系, 在水深較淺的區域, DO濃度相對較高, 因而湖濱帶的N2O排放量通常最高[51]。此外, 對氧氣需求相反的硝化細菌與反硝化細菌在整個氮素循環中均起到重要作用, 因而N2O產生速率與DO的關系相對復雜[52], 在本研究中N2O交換通量與DO顯著負相關, 控制系統內冬、春季的DO濃度高于夏、秋季, 較高的DO濃度能夠抑制水體中的反硝化作用[12]。因此, 在本研究期間, 控制系統內冬、春季水-氣界面間N2O主要吸收趨勢為主, 而在夏、秋季則表現出漸強的N2O排放趨勢。
控制系統內N2O交換通量與TN含量的相關性并不顯著, 而與TP、顯著正相關, 與N顯著負相關。這可能是由于水體中TP含量高的富營養化淺水湖泊會導致有關反硝化細菌代謝需求的N和P比例發生變化, 進而影響反硝化過程和N2O的生成[13], 由此可能改變水體的營養狀態, 減弱湖泊應對氣候變化的前哨作用[53]。我們的研究結果顯示, 升溫能夠顯著提升水體中的含量并且促進水-氣界面間N2O的交換, 而添加磷營養鹽則表現出對水體中含量和N2O交換通量顯著的抑制作用。以上表明無機氮是N2O生成過程中重要的環境因子, 但在氣候變化的趨勢下對富營養化湖泊生態系統水-氣界面間N2O交換的響應機制發生了變化。
水體DOC含量能夠反映有機質的動態積累過程[54], 本實驗期間觀察到夏季水體中DOC的含量顯著高于其他季節, 同時Chl.a的含量亦顯著增加, 表明隨季節交替控制系統內有機質逐漸積累并且浮游植物的密度逐步升高。初級生產者本身即是有機質的載體, 同時也會顯著影響控制系統內的生物地球化學循環。Forshay和Dodson[55]發現在水生植物的生長過程中沉水植被豐富的沉積物比無沉水植被的沉積物具有更高的反硝化速率, 其原因是有沉水植物的沉積物在反硝化過程中不會受到碳的限制。在本研究中, 控制系統內沉水植物同樣對沉積物-水界面間N2O的生成產生顯著促進作用。水生植物衰亡后產生的大量凋落物也能為硝化細菌提供充足的碳源和豐富的底物, 亦有利于N2O的生成和排放[56]。控制系統內沉水植物與浮葉植物隨季節的交替, 整個生命周期內有機質不斷積累。因此, N2O交換通量與DOC呈顯著正相關關系表明有機質積累促進了沉積物界面N2O的生成。
4 結論
(1)升溫顯著促進了水-氣界面間N2O的排放,表現出與代謝理論的一致性, 而不同的升溫模式間N2O的交換過程無明顯差異。(2)磷的添加對水-氣界面間N2O的排放過程表現出明顯的抑制作用, 其原因可能是磷的添加促進了初級生產者的生長, 提高了水體環境中氮的利用。N2O的年累積交換量進一步表明, 磷的添加改變了湖泊生態系統水-氣界面間N2O的源匯轉換動態。(3)升溫與磷營養鹽交互作用下環境因子與N2O交換通量有著顯著的相關關系, 控制系統內和沉水植物PVI指數(冬春)表現為顯著正相關, DO、表現為顯著負相關。(4)水-氣界面間N2O的通量與無機氮含量顯著相關, 而升溫與磷添加的交互作用下N2O均表現出相同的變化趨勢,較高的TP含量可能影響反硝化代謝過程中更為適宜的N和P比例, 導致水體的營養狀態發生改變, 進而削弱水-氣界面N2O的排放。(5)初級生產者是有機質的主要載體, 升溫及與磷營養鹽添加的交互作用顯著促進初級生產者的生長。實驗期間隨著季節的交替, 有機質逐漸積累, 水-氣界面間N2O的排放也顯著強烈。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
當代陜西(2020年13期)2020-08-24 08:22:02
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
制造技術與機床(2017年5期)2018-01-19 02:49:17
金秋(2017年4期)2017-06-07 08:22:16
中國材料進展(2016年10期)2016-12-26 06:50:20
濰坊學院學報(2016年2期)2016-12-01 13:00:11
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
