基于代謝組學分析平菇栽培用培養料發酵過程中代謝物的變化
2021-06-02 11:04:29劉皓皓劉芹崔筱靳榮線高玉千李亞楠孔維麗裴瑞杰邱立友
中國瓜菜 2021年4期
劉皓皓 劉芹 崔筱 靳榮線 高玉千 李亞楠 孔維麗 裴瑞杰 邱立友


摘 要:為了解發酵培養料制備過程中微生物代謝產物的組成和變化規律,采用非靶向代謝組學技術,檢測分析發酵2、4、6、8和10 d時培養料的代謝組數據。結果表明:在正離子(POS)模式下,分別在T2 vs T1、T3 vs T2、T4 vs T3和T5 vs T4中篩選得到331、145、161和115種差異代謝物;在負離子(NEG)模式下,分別篩選得到251、159、106和76種差異代謝物。這些差異代謝物主要包括糖類及其衍生物、氨基酸、肽類及其類似物、脂肪酸類、維生素類、苯丙素類和聚酮類物質及其他次級代謝產物。此外,在發酵培養料中還含有植物生長調節劑(如吲哚-3-乙酸、茉莉酸和赤霉素)和抑菌物質(如綠原酸、抗生素、白藜蘆醇)。該研究可為制備質量穩定的發酵培養料和掌握發酵培養料栽培平菇技術提供參考。
關鍵詞:平菇;發酵料;代謝組學;差異代謝物
中圖分類號:S646.1+4 文獻標志碼:A 文章編號:1673-2871(2021)04-021-10
Abstract: Non-targeted metabolomics technique was used to reveal the dynamic changes in the composition and variation of microbial metabolites in corncob composting for preparing cultivation medium of Pleurotus ostreatus. Results showed that 331, 145, 161 and 115 differential metabolites at POS mode were screened, including carbohydrates and derivants, amino acids, peptides, and analogues, fatty acids, vitamins, phenylpropanoids and polyketides, and other secondary metabolites, between T2 vs T1, T3 vs T2, T4 vs T3 and T5 vs T4, respectively. And 251, 159, 106 and 76 differential metabolites at NEG mode were screened, respectively. Furthermore, the fermentation materials contained plant growth regulators (such as indole-3-acetic acid, jasmonic acid and gibberellin) and antibacterial substances (such as chlorogenic acid, antibiotics, resveratrol). This study provided a reference for the preparation of quality-stable fermentation materials and the further understanding of the cultivation of Pleurotus ostreatus with fermentation materials.
Key words: Pleurotus ostreatus; Fermentation material; Metabolomics; Differential metabolites
平菇Pleurotus ostreatus (Jacq.) P. Kumm.,是世界上栽培最廣泛的食用菌之一[1-3],也是重要的食藥用真菌[4-5]。平菇的栽培技術主要有3種:生料栽培、熟料栽培和發酵料栽培[6]。與生料和熟料栽培相比,發酵料栽培平菇由于具有低污染、低成本、工藝簡單和經濟效益高的優點而在世界范圍內得到了廣泛應用[6]。平菇栽培用發酵料是以秸稈、玉米芯等農業副產物為主要原料,加入少量的麩皮、石灰等,在微生物的參與下,經短期、好氧發酵而制備的[3]。玉米芯約占玉米產量的21%,是一種產量巨大的農副產品,來源廣泛、價廉易得[7]。目前,玉米芯除用于制備糠醛、木糖醇等外,很大一部分被作為農業廢棄物直接燃燒,造成資源浪費和環境污染。玉米芯組織均勻、硬度適宜、吸水性強,將其堆制發酵后用于平菇栽培,不僅有助于平菇產量和品質的提高,而且可以實現農業副產物的高效生物轉化[8]。
筆者在前期的研究中發現:平菇玉米芯發酵培養料制備過程中,變形菌門Proteobacteria在早期階段(T1)占優勢(相對豐度35.67%~44.10%),在高溫階段(T2,T3),厚壁菌門Firmicutes成為新的優勢門(相對豐度41.50%~48.52%),而在堆肥后期(T4、T5),變形菌門重新成為優勢門(相對豐度33.55%~36.89%)[9]。這些微生物對發酵過程中植物秸稈中木質纖維素的降解起著至關重要的作用。Pearson相關分析表明,發酵培養料制備過程中的理化性質變化與其中微生物菌落的組成變化顯著相關[9]。微生物能將培養料中復雜的有機質(纖維素、半纖維素、木質素等)進行降解,使其變成簡單、易于利用的低分子物質;有些特定微生物如Actinobacteria、Thermus和Bacillus還可以產生抗菌、殺蟲的物質,這可能是發酵培養料栽培平菇可以采用開放式接種的原因之一[10]。
目前對玉米芯發酵培養料的研究主要集中在栽培平菇[11-13]和制備過程中發酵培養料中微生物及其功能的演替方面[3],而對玉米芯發酵培養料制備過程中微生物代謝產物的研究還未見報道。代謝組學(metabolomics)是繼基因組學、轉錄組學、蛋白質組學后出現的新興組學技術,是系統生物學的重要組成部分[14]。代謝組學是從整體角度出發,運用現代檢測技術對盡可能多的代謝產物進行分析檢測[15]。目前,代謝組學在真菌領域的應用也日益廣泛,為人們提供了一個了解真菌代謝的獨特途徑[14-16]。
因此,筆者通過采用非靶向代謝組學技術分析檢測玉米芯發酵培養料制備過程中發酵微生物代謝產物的種類和含量,進而探討制備過程中主要代謝物的組成及變化規律,為穩定發酵料質量提供理論基礎。
1 材料與方法
1.1 發酵料
發酵料的質量分數配比為:玉米芯84%,麩皮10%,石灰5%,尿素1%;含水量68%。
1.2 試劑與儀器
玉米芯(中值粒徑0.5 cm)、麩皮、石灰、尿素均購自本地農貿市場;LC-MC級甲醇、水、甲酸、醋酸銨(CNW Technologies GmbH, Germany)。
BSA124S-CW天平(Sartorius);1290 UHPLC 色譜儀(Agilent Technologies Inc., USA);Triple TOF 6600質譜儀(AB Sciex Pte. Ltd., USA);D3024R高速冷凍離心機(Scilogex, USA);JXFSTPRP-24全自動樣品快速研磨儀(上海凈信實業發展有限公司);GWB-1普析純水儀(北京普析通用儀器有限責任公司);KS-7200DV超聲儀(昆山潔力美超聲儀器有限公司)。
1.3 發酵料制備和取樣
該試驗于2020年3—4月在河南省農業科學院現代農業科技試驗示范基地進行。根據以往的報道[12],制備平菇發酵料。發酵料制備試驗包含6個重復,每個重復包含250 kg原料(干質量)。玉米芯原料按照質量比為1∶2.4~2.5的料水配制,機械攪拌30 min,堆置1 d后進行建堆,堆高60 cm,寬1.5 m左右,長度不限。此時培養料的含水量約為70%,pH值9~10,顏色為金黃色。用直徑5 cm的木棒在料堆上部、橫豎間隔30~40 cm打通風孔,木棒要求插到料堆底部。發酵周期為10 d,第3天溫度升至45~55 ℃開始翻堆,培養料溫度升至70 ℃以上每隔1 d翻堆1次,共4次,直至發酵完成。分別在發酵2、4、6、8、10 d時采集樣品,標注為T1、T2、T3、T4、T5。為了獲得具有代表性的樣本,分別從發酵料堆的9個不同位置采集子樣本,然后將子樣本進行混合得到一個樣本[3]。
1.4 代謝組學分析
取發酵培養料樣品進行代謝組學分析,按北京諾禾致源科技股份有限公司非靶向代謝組學分析方法提取代謝物、超高效液相色譜-四級桿串聯飛行時間質譜(ultra performance liquid chromatography-quadrupole time-of-flight mass spectrometry,UPLC-QTOF-MS)分析代謝物,采用ProteoWizard和XCMS軟件對代謝組下機數據(.raw)實施峰的識別、提取、積分及對齊等處理,利用mzCloud(https://www.mzcloud.org/)、mzVault和Masslist數據庫進行峰比對,同時采用blank樣本去除背景離子,進一步進行峰面積的批次歸一化和自適換算標準化處理,從而得到代謝物的鑒定和定量結果。采用SIMCA軟件(version 14.1, sartorius stedim data analytics AB, Umea, Sweden)對數據進行主成分分析(principal component analysis, PCA)和正交偏最小二乘判別分析(orthogonal partial least squares discriminant analysis, OPLS-DA)[17]。
1.5 數據處理分析
試驗數據均采用Excel 2016進行記錄整理,采用SPSS(version 20.0)進行Pearson相關性分析以及單因素ANOVA檢驗比較平均值,統計分析結果p<0.05為差異性顯著,試驗結果用平均值和SEM表示。
2 結果與分析
2.1 發酵培養料代謝物的主成分分析
基于非靶向代謝組學技術對發酵2、4、6、8、10 d的培養料樣品進行檢測,分別在正離子(POS)和負離子(NEG)模式下鑒定得到1208和472種代謝物,對所有代謝物進行PCA分析(圖1)。從圖1可以看出,培養料樣品中的代謝物分成5組,組內樣本均集聚良好,這說明5組培養料樣品的微生物代謝物具有顯著的差異。
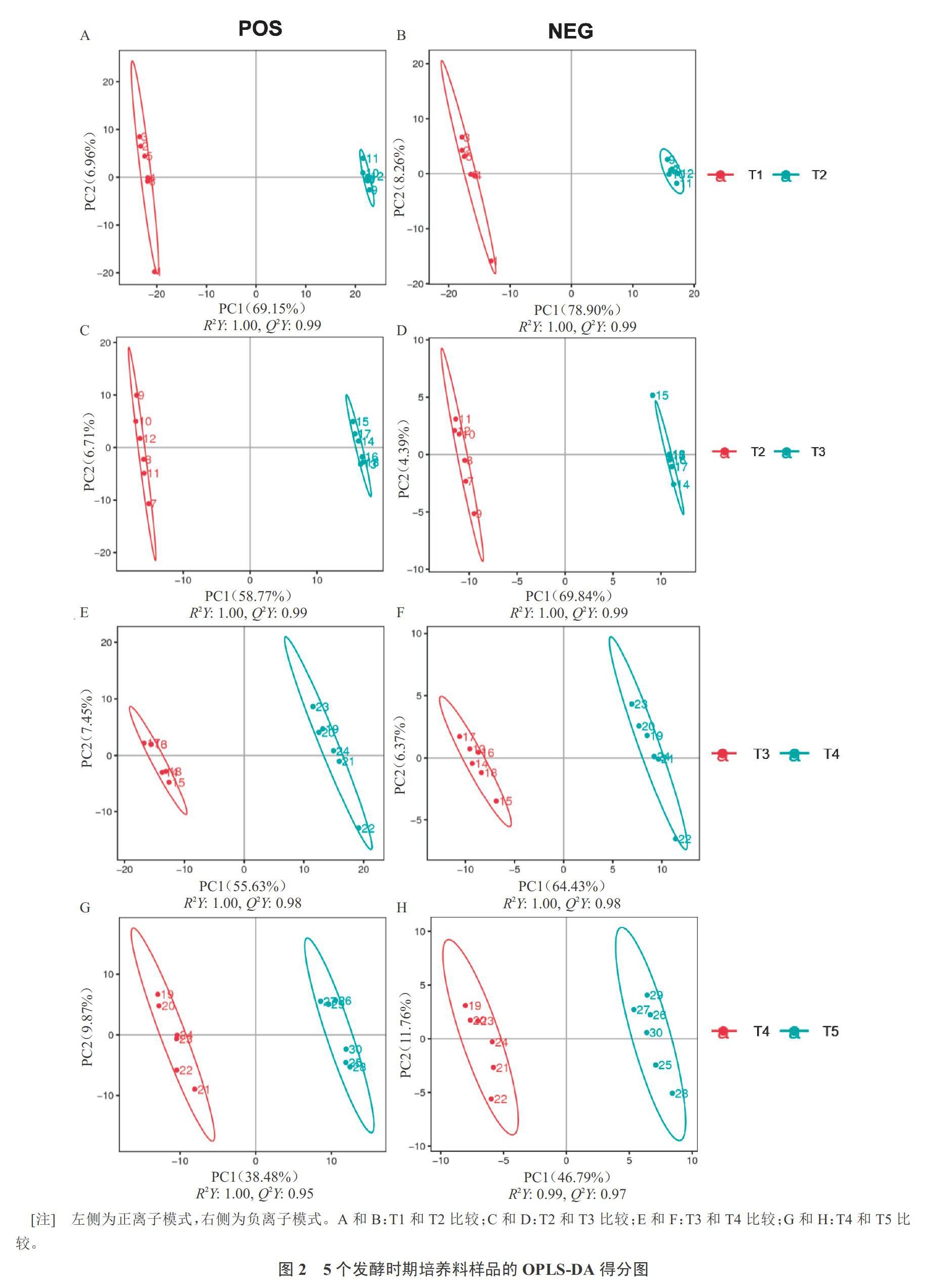
為更充分地提取不同發酵時期培養料樣品中代謝物的差異信息,分析篩選不同組間的差異代謝物,因此進一步采用有監督的OPLS‐DA分析數據。由圖2可知,T2 vs T1(圖2-A~B)、T3 vs T2(圖2-C~D)、T4 vs T3(圖2-E~F)、T5 vs T4(圖2-G~H)中的樣品點均分布在不同的區域且明顯分為兩個簇,實現了兩組樣品間的完全分離。結果表明,不同發酵時期培養料樣品中微生物代謝產物在種類和(或)含量上存在明顯差異。POS模式下T2 vs T1、T3 vs T2、T4 vs T3、T5 vs T4的OPLS‐DA模型質量參數分別為:R2Y(cum)=1.00,Q2Y(cum)=0.99;R2Y(cum)=1.00,Q2Y(cum)=0.99;R2Y(cum)=1.00,R2Y(cum)=0.98;R2Y(cum)=1.00,Q2Y(cum)=0.95。NEG模式下分別為:R2Y(cum)=1.00,Q2Y(cum)=0.99;R2Y(cum)=1.00,Q2Y(cum)=0.99;R2Y(cum)=1.00,Q2Y(cum)=0.98;R2Y(cum)=0.99,Q2Y(cum)=0.97。R2Y和Q2Y均大于0.5,這表明依據兩種采集模式所得數據建立的OPLS‐DA模型都具有較好的擬合性和較強的預測能力。
同時采用200次響應的置換檢驗來驗證模型的過擬合現象,避免采用OPLS-DA方法分析時出現過擬合現象。由圖3可知,Q2的回歸直線與y軸的交點均在負半軸。POS模式下,T2 vs T1的R2Y(cum)(0,0.81),Q2(cum)(0,-0.68);T3 vs T2的R2Y(cum)(0,0.91),Q2(cum)(0,-0.67);T4 vs T3的R2Y(cum)(0,0.90),Q2(cum)(0,-0.67));T5 vs T4的R2Y(cum)(0,0.91),Q2(cum)(0,-0.73)。NEG模式下,T2 vs T1的R2Y(cum)(0,0.51),Q2(cum)(0.0,-0.85);T3 vs T2的R2Y(cum)(0,0.83),Q2(cum)(0,-0.74);T4 vs T3的R2Y(cum)(0,0.83),Q2(cum)(0,-0.79);T5 vs T4的R2Y(cum)(0,0.81),Q2(cum)(0,-0.87)。這說明建立的OPLS‐DA模型是穩健可靠的,不存在過擬合現象。因此,可以用來探索發酵培養料制備過程中,不同發酵階段培養料樣品中的微生物代謝產物的差異。
2.2 發酵料制備過程中的差異代謝物的篩選
以p<0.05和VIP>1為條件篩選出具有差異性表達的化合物,T2 vs T1、T3 vs T2、T4 vs T3、T5 vs T4,在POS模式下,分別得到331、145、161和115種差異代謝物;在NEG模式下,分別得到251、159、106和76種差異代謝物(圖4)。
對檢測到的代謝物進行log2處理后,在POS和NEG模式下變化最顯著的20種(上調和下調)差異代謝物見表1。T2 vs T1時,發酵培養料樣品中1-(2-呋喃基)戊烷-1,4-二酮、2-(4-氯苯基)-N,N-二甲基乙硫酰胺、D-氨基葡萄糖、N-(4-氯苯基)-N'-(2-苯氧基苯基)等物質相對含量顯著增加;D-(+)-麥芽糖、5-羥賴氨酸、D-棉子糖、α,α-海藻糖等物質相對含量顯著降低。T3 vs T2時,截短側耳素、圣草酚、苯乙醛等物質相對含量顯著增加;而黑色素、縮二脲、丁酸賴氨酸等物質相對含量顯著降低。T4 vs? T3時,阿魏酸、黃豆苷元、橙皮素等物質相對含量顯著增加;對香豆酰胍丁胺、兒茶酚、原兒茶酸等物質相對含量顯著降低。T5 vs T4時,L-正亮氨酸、7-羥基香豆素、環氧霉素等物質相對含量顯著增加;兒茶酚、茶堿、2-氨基-1,3,4-十八碳三醇等物質相對含量顯著降低。
2.3 不同發酵時期差異代謝物含量比較
對不同差異代謝物進行歸類,T2和T1、T3和T2、T4和T3、T5和T4的之間的差異代謝物的比較見圖5。在不同發酵時間的樣品中檢測出的差異代謝物主要有:糖類及其衍生物(棉子糖、蔗糖、核糖、乳糖、氨基葡萄糖、阿拉伯糖醇等),脂肪酸類(壬酸、戊酸、己酸、辛酸、順式5,8,11,14,17-二十碳五烯酸、16-羥基十六烷酸等),氨基酸、肽類及其類似物(N-乙酰基天冬氨酸、甘氨酰-L-亮氨酸、L-谷氨酸、天冬酰胺、天冬氨酸、D-丙氨酰-D-丙氨酸等),維生素類(生物素、核黃素、煙酸、泛酸、吡哆醇等),抑菌物質類(綠原酸、截短側耳素、土霉素、環氧霉素、橙皮素、白藜蘆醇等),苯丙素類和聚酮類物質(3-(4-羥基苯基)丙酸、柚苷二氫查耳酮、香豆素、3,4-二羥基苯基丙酸等),核苷、核苷酸及類似物(黃嘌呤、尿苷、cGMP、鳥苷一磷酸、脫氧胞苷等)和其他代謝物(6β-羥基皮質醇、對香豆酰胍丁胺、1-(4-溴苯基)2-苯基乙-1-酮等)等。從代謝組數據中提取上述物質的相對峰面積,峰面積與含量成正比,可以用來表征代謝物的含量。其中糖類及其衍生物和氨基酸、肽類及其類似物的變化趨勢相似,T2 vs? T1和T3 vs T2,糖類及其衍生物的相對含量分別下降了5.00%和4.83%,而氨基酸、肽類及其類似物相對含量分別下降了3.36%和5.92%;T4 vs T3和T5 vs T4,糖類及其衍生物的相對含量分別上升了2.43%和5.05%,而氨基酸、肽類及其類似物相對含量分別上升了1.54%和6.03%。與T1相比,T2時期發酵培養料中的苯丙素類和聚酮類物質相對含量下降了4.39%,而T3、T4和T5時的苯丙素類和聚酮類物質相對含量分別比發酵前一時期(T2、T3和T4)高9.47%、0.91%和3.15%。抑菌物質和維生素類的相對含量均較發酵前一時期有所增加,其中T2、T3、T4和T5發酵料中抑菌物質相對含量較T1、T2、T3和T4分別上升了7.28%、13.84%、3.83%和3.81%;而維生素類相對含量分別上升了0.14%、13.66%、6.96%和4.77%。
2.4 不同發酵時期植物生長調節劑豐度變化
從圖6可以看出,不同發酵時期培養料中吲哚-3-乙酸和赤霉素的相對峰面積變化趨勢相似,在T5時期達到最高值,茉莉酸的相對峰面積隨發酵時間的延長呈現出先升高后下降的趨勢,在T2時達到最高值。脫落酸相對峰面積變化趨勢與茉莉酸類似,最大值出現在T4。
3 討論與結論
筆者在本研究中采用非靶向代謝組學技術對不同發酵時期的平菇培養料中的代謝物進行分析,差異代謝物總數最多的是T2 vs T1,其次是T3 vs T2,最少的是T5 vs T4。這與筆者前期對5個發酵時期培養料中微生物的代謝功能差異的分析結果基本一致[3]。發酵培養料中的漆酶、木聚糖酶和蛋白酶活性峰值出現在發酵降溫階段[3],表明木質纖維素和蛋白質的分解主要是在發酵后期發生,因此T2 vs T1和T3 vs T2時糖類及其衍生物和氨基酸、肽類及其類似物相對含量顯著下降。這可能是因為發酵前期(T2和T3)微生物代謝旺盛,需要大量的糖類和氨基酸以滿足其生長和繁殖所需,導致T2和T3時發酵料中的糖類和氨基酸含量均較發酵前一時期(T1和T2)有所下降;到發酵后期(T4和T5),木質纖維素降解酶和蛋白酶活性大幅升高,大量木質纖維素和蛋白質被降解生成低分子糖、肽類和氨基酸,因而其含量均比發酵前一時期(T3和T4)有所增加。發酵料中的Actinobacteria、Thermus和Bacillus屬的微生物具有產生抑菌物質的能力[18]。發酵料中的抑菌物質含量隨發酵進程逐漸增加,為后期平菇開放式接種創造了良好的條件。這與筆者前期研究結果一致,隨著發酵時間的延長,發酵料浸提液對青霉和木霉的抑制作用也隨之增強[19]。維生素可以作為輔酶因子參與多種重要的生化反應,是生物不可或缺的生長因子。不同微生物之間可能憑借維生素的種間傳遞而建立相互作用關系,對微生物群落結構形成和功能發揮起著重要作用[20]。與抑菌物質含量變化趨勢相似,發酵培養料中的維生素含量也較發酵前一時期有所增加。在自然界中,木質纖維素可以被微生物分解成低分子的芳香族化合物,如水楊酸、阿魏酸等[21-22]。與T1相比,T2時期發酵培養料中的苯丙素類和聚酮類物質相對含量明顯下降,這可能是因為微生物優先利用易降解的低分子物質,而T3、T4和T5時的苯丙素類和聚酮類物質相對含量分別比發酵前一時期有所增加,隨著發酵的繼續,玉米芯中易于降解的物質被利用完,微生物開始降解復雜高分子物質如木質素和酚類物質,進而生成了較多的低分子苯丙素類和聚酮類物質。研究表明,植物生長調節劑如吲哚乙酸、赤霉素、脫落酸可以促進平菇菌絲的生長[23-24]。筆者前期研究發現,T5時期發酵培養料浸提液對平菇菌絲生長促進作用最為明顯,尤其是體積分數為80%時,平菇菌絲生長速度和生物量分別比對照增加0.53 cm·d-1和0.179 g[25]。這可能與發酵培養料中的植物生長調節劑如脫落酸、吲哚-3-乙酸和赤霉素的相對含量均在發酵后期達到最高值有關。
綜上所述,不同發酵時期的玉米芯培養料中的微生物代謝產物具有明顯的差異,并且其中含有豐富的抑菌物質和植物生長調節劑。該研究為平菇栽培獲得穩定的發酵料質量提供了科學依據和理論基礎。
參考文獻
[1] NAM W L,SU M H,PHANG X Y,et al.Production of bio-fertilizer from microwave vacuum pyrolysis of palm kernel shell for cultivation of Oyster mushroom(Pleurotus ostreatus)[J].Science of The Total Environment,2018,624:9-16.
[2] JAYASINGHEARACHCHI H S,SENEVIRATNE G.Can mushrooms fix atmospheric nitrogen[J].Journal of Biosciences,2004 29(3):293-296.
[3] KONG W L,SUN B,ZHANG J Y,et al.Metagenomic analysis revealed the succession of microbiota and metabolic function in corncob composting for preparation of cultivation medium for Pleurotus ostreatus[J].Bioresource Technology,2020,306:123156.
[4] 戴玉成,楊祝良.中國藥用真菌名錄及部分名稱的修訂[J].菌物學報,2008,27(6):801-824.
[5] S?NCHEZ C.Cultivation of Pleurotus ostreatus and other edible mushrooms[J].Applied Microbiology and Biotechnology,2010,85(5):1321-1337.
[6] HERN?NDEZ D,S?NCHEZ J E,YAMASAKI K.A simple procedure for preparing substrate for Pleurotus ostreatus cultivation[J].Bioresource Technology,2003,90(2):145-150.
[7] 余敏佳,宋婷婷,范麗軍,等.培養料中玉米秸和玉米芯的不同顆粒度對栽培雙孢蘑菇效應[J].食藥用菌,2018,26(5):289-293.
[8] 李秋月,馮占,金宇昌,等.不同產地玉米芯的持水率及其相關性分析[J].食藥用菌,2020,28(6):412-417.
[9] KONG Z J,WANG X Q,WANG M M,et al.Bacterial ecosystem functioning in organic matter biodegradation of different composting at the thermophilic phase[J].Bioresource Technology,2020,317:123990.
[10] CARRASCO J,PRESTON G M.Growing edible mushrooms:a conversation between bacteria and fungi[J].Environmental Microbiology,2020,22(3):858-872.
[11] 劉志強.平菇發酵料袋栽技術[J].現代農業科技,2019(24):58-59.
[12] 孔維麗,崔筱,袁瑞奇,等.河南省適宜發酵料栽培的平菇優良品種篩選[J].中國食用菌,2019,38(11):23-28.
[13] 聞亞美,邵欣欣,丁亞通,等.7種平菇發酵料生產拌料用殺菌劑藥效對比及安全性評價[J].中國瓜菜,2020,33(8):49-52.
[14] 雷露,吳天祥,王川南.基于UPLC-QTOF-MS代謝組學研究灰樹花發酵的代謝差異[J].菌物學報,2020,39(10):1920-1932.
[15] BOLTEN C J,KIEFER P,LETISSE F,et al.Sampling for metabolome analysis of microorganisms [J].Analytical of Chemistry,2007,79(10):3843-3849.
[16] 尉杰,趙鋮,陸瑞利,等.菌物研究中的代謝組學應用及存在的問題[J].菌物研究,2020,DOI:10.13341/j.jfr.2020.0003.
[17] 張萌,羅芳,王敏,等.奶牛分娩后早期血漿代謝物變化研究[J].生物技術通報,2020,36(6):191-199.
[18] ZHANG X,ZHONG Y,YANG S,et al.Diversity and dynamics of the microbial community on decomposing wheat straw during mushroom compost production[J].Bioresource Technology,2014,170:183-195.
[19] 崔筱,劉芹,丁亞通,等.平菇培養料發酵后對霉菌抑制作用機理研究[J].河南農業科學,2021,50(3):109-116.
[20] 葛磊鑫.基于B族維生素合成與利用特征的湖泊微生物種間關系分析[D].武漢:華中師范大學,2019.
[21] 歐陽光察,薛應龍.植物苯丙烷類代謝的生理意義及其調控[J].植物生理學通訊,1988,9(3):9-16.
[22] 王強.降解木質纖維素的側耳屬菌株的篩選及酶學特性研究[D].河北邯鄲:河北工程大學,2019.
[23] 吳亞召,李峻志,李安利,等.脫落酸在平菇生產中的應用研究[J].中國食用菌,2011,30(4):36-37.
[24] 李輝,張志強,田昊.植物生長調節劑對平菇菌絲生長的影響[J].北方園藝,2013(23):158-160.
[25] 胡素娟,宋凱博,劉芹,等.玉米芯發酵料水浸提液促平菇菌絲生長的機理研究[J].河南農業大學學報,2021,https://doi.org/10.16445/j.cnki.1000-2340.20210323.001.
