半導體合成生物學的研究進展
2021-06-03 07:39:28王欣趙鵬李清揚田平芳
化工學報 2021年5期
關鍵詞:生物
王欣,趙鵬,李清揚,田平芳
(1北京化工大學生命科學與技術學院,北京100029;2華南理工大學食品科學與工程學院,廣東廣州510641)
引 言
合成生物學(synthetic biology)是從最基本要素開始設計和構建新生物體系,或修改現有生物體系的一門交叉學科[1-2]。近年來,人工基因電路已取得顯著進展,在此基礎上耦合半導體材料形成了生物-非生物混合體系(living-nonliving hybrid system)。在該雜合體系中,人工細胞可與半導體材料建立聯系,由此形成一個新的研究方向——半導體合成生物學(semiconductor synthetic biology/SemiSynBio,SSB)。半導體合成生物學探索工程細胞與半導體材料之間的協同作用。無論是半導體材料還是細胞,二者都涉及電子的受控流動,區別在于半導體材料涉及物理現象中電子在線路中的長程運動,而細胞則涉及化學反應中電子在分子間的短程運動。當半導體縮小到物理極限時,就能幾乎匹配活細胞以化學方式處理的電子傳遞。因此在半導體合成生物學領域,物理信號將超越以往簡單的傳遞方式,轉變為細胞-半導體間的雙向通信,即一方面細胞能接收來自半導體的電、熱、機械等物理信號,從而調控其代謝行為;另一方面半導體材料也能感知來自細胞的電子、代謝物及生物大分子等信號,從而實現物質或信號的輸出[3]。由此可見,半導體合成生物體系包括:(1)以活細胞為感應基礎的“生物前端”層;(2)以非生物材料為信息計算模塊的“半導體后端”層。因此,該研究方向屬于交叉學科,具有理論和應用雙重意義。本文綜述了近年來半導體合成生物學具有代表性且發展迅速的領域。
1 半導體-細胞雜交光合體系介導的生物催化
為滿足日益增長的能源需求并克服化石燃料的局限性,人們正在尋求可持續的新能源生產方法。光合作用對碳元素的利用率接近100%,但整個過程的能量轉換效率非常低(一般在5%以下)[4]。固態半導體光吸收器比生物體更能有效地捕獲光,能量轉換效率接近20%(Shockley-Queisser極限為33.7%)[5-6]。因此,將高選擇性生物催化體系和高效光收集器進行集成,建立混合系統,其碳利用效率將超過天然光合作用[7-9]。該半導體-細胞混合體系包括三部分:細胞、半導體材料及界面接口。
細胞通過復雜的表面結構與外環境進行物質與能量交換。非光合微生物例如大腸桿菌(Escherichiacoli)[10]、酵 母 菌(Saccharomyces cerevisiae)[11]、卵形鼠孢菌(Sporomusa ovata)[12]、熱醋穆爾氏菌(Moorella thermoacetica)[13]和羅爾斯通氏菌(Ralstonia eutropha)[14-15]等已被開發為半導體-細胞雜交體系進行人工光合作用。在這種混合雜交系統中,厭氧菌能在有氧條件下接收外界非生物組件傳遞的信號或還原當量,之后利用細胞內特有途徑進行氧化-還原反應,從而合成化學品或能源燃料。非生物部分應具有親微生物表面,可促進材料和微生物的穩定整合。與聚合物和金屬相比,半導體材料因其在生物界面處的多種信號傳導機制而更適用于電子和光子生物界面。同時,高性能無機半導體可被精確制造成各種納米級結構,以匹配亞細胞和分子成分的大小。例如,硅納米線(SiNWs)的直徑(d=1~100 nm)比哺乳動物細胞(dcell≈10μm)小幾個數量級,且具有較大的長徑比(約103),這有助于在分子水平上研究復雜的信號調控模式[16]。除硅(Si)[17]之外,常見的非生物體部分還包括磷化銦(InP)[11]、硫化鎘(CdS)[13,18]、磷酸鈷(CoP)[19]、金納米團簇(AuNCs)[20]、石墨相碳氮化物(g-C3N4)[14-15]等半導體材料。生物體和非生物體之間的耦合需要穩定且高效的界面接口,如此才能構建與自然系統相同的信號傳導機制,從而準確地調控細胞-半導體混合系統。界面接口的形成則需要細胞和半導體材料間極為緊密和高表面積的接觸,以便維持細胞活力和半導體性能[21]。在細胞-半導體雜交體中實現這種界面需要做到三個方面:(1)選擇并設計適當的生物和非生物成分以確保二者的相容性;(2)通過親和結合或自組裝進行化學耦合;(3)建立人工材料與細胞之間的能量轉導及耦合。
細胞與半導體的連接一般為直接物理接觸(圖1)。Kim等[16]將哺乳動物細胞培養在含垂直排列的SiNW陣列的基底上,數天后,SiNW陣列與正在生長的細胞緊密結合,該研究表明無外力作用下半導體材料與細胞的自然結合。Sakimoto等[13]則采用生物沉淀的方法使M.thermoacetica表面附著CdS納米顆粒,建立了良好的生物界面。再如,Wei等[10]在大腸桿菌中融合表達外膜蛋白OmpA和金屬結合蛋白PbrR,可特異性吸附Pb和Cd離子,從而在菌表面形成PbS和CdS界面層,促使菌體在有氧條件下持續產氫。Tremblay等[14-15]則將g-C3N4顆粒與R.eutrophaH16共培養構建雜交光合系統,轉化光能后得到部分還原當量,然后通過乙酰乙酰輔酶A還原酶PhbB直接供給聚羥基丁酸酯的合成。在人工光合雜交系統中,半導體光吸收器具有較高的載流子遷移率,比生物體更高效地吸收光,但卻不能有效地將光激發電子的能量轉移至碳鍵合成中。因此,利用非光合細菌的優異胞內固碳能力將半導體吸收的光能轉換成化學能,從而最大限度地利用太陽能。此前研究中,自養細菌已廣泛用于生產簡單有機分子,而將異養生物與半導體光收集器融合,則在復雜代謝物的合成方面更具優勢。
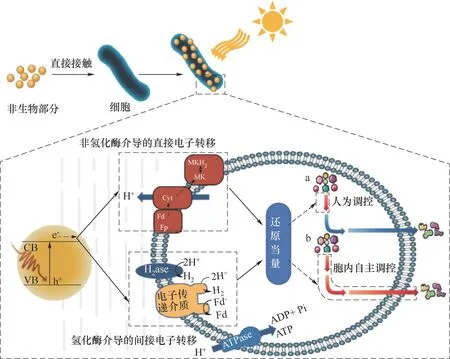
圖1 半導體-細胞雜合體系介導的生物催化示意圖Fig.1 Schematic diagram of biocatalysis mediated by semiconductor-cell hybrid system
人工光合雜交系統的電子轉移機制可根據細胞膜上是否存在氫化酶而分為兩種:(1)非氫化酶介導的直接電子轉移,該過程發生在光合作用的前3 h,其特點符合慢電荷轉移動力學,當細胞與半導體直接接觸后,通過自身的還原蛋白[例如,細胞色素(Cyt)、鐵氧還蛋白(Fd)、黃素蛋白(Fp)]或導電鞭毛將胞外電子傳遞到自身細胞的化學反應中,并不依賴H2或NAD(P)H作為還原當量的來源[22];(2)氫化酶介導的間接電子轉移,該過程遵循驅動電子到膜結合氫化酶的電荷轉移動力學,在24 h內積累足量的H2、甲酸等代謝物,并通過HydABC絡合物氧化進入Wood-Ljungdahl途徑,為細胞提供還原當量(圖1)[23-25]。總之,以細胞色素和氫化酶為代表的膜結合蛋白在電子-空穴對分離過程中發揮重要的電子傳遞作用[22,26]。Jensen等[27]在大腸桿菌中異源表達希瓦氏菌(Shewanella oneidensis)的胞外電子傳遞色素蛋白(CymA、MtrA、MtrB、MtrC),重構了胞外電子傳遞路徑(CymA-MtrCAB),使外膜上的固體金屬氧化物被還原。該研究表明,合成生物學可改變細胞特性,使其成為與無機納米材料相容的界面細胞。因此,研究重點將著眼于提高材料的電子-空穴對分離效率以及電子從半導體到細胞的轉移能力[28-29]。深入了解電子在細胞內的分子響應機制有助于改進半導體材料對信號的集成和傳導性能。
當電子或還原當量通過細胞膜進入胞內,細胞內分子響應機制可分為人為調控和胞內自主調控(圖1)。人為調控是根據需求對胞內代謝途徑進行改造,使細胞從半導體材料中獲得的能量集中用于某特定代謝途徑或途徑中某一步驟,對細胞獲得的還原力進行有目的地分配。該方面Guo等[11]構建的基因工程酵母-InP雜化平臺是一成功案例。NADPH是生物合成中的關鍵氧化還原輔因子。酵母中的NADPH主要來自磷酸戊糖途徑(PPP)。當缺失葡萄糖-6-磷酸脫氫酶基因zwf1時,PPP的氧化部分被破壞,極大降低胞漿NADPH的再生能力,直接影響莽草酸的合成,最終導致其前體3-脫氫莽草酸(DHS)的積累[30]。研究者在此基礎上利用附著在酵母細胞表面的光收集半導體顆粒InP提供缺失的還原當量,促使DHS高效合成莽草酸[11]。此外,在釀酒酵母表面覆蓋氮化鎵(GaN)納米薄膜,使其能夠吸收紫外線提供的能量,在細胞表面積累電荷,激活細胞壁中的幾丁質合成途徑[31]。另一種胞內自主調控機制則是遵循細胞正常代謝途徑所需,利用半導體材料的電子或還原當量生產簡單有機小分子。如Sakimoto等[13]建立了M.thermoacetica-CdS雜交體系,CdS經光照激發的光生電子產生還原當量[H]直接進入Wood-Ljungdahl途徑,驅動CO2經乙酰-CoA合成乙酸。Wood-Ljungdahl途徑是已知厭氧生物固碳途徑中能耗最低、路徑最短的途徑,符合細胞自主調節的能量分配原則。
半導體-細胞雜交體系介導的生物催化發展極為迅速,已開發出若干細胞和半導體材料的組合。除光合雜交體系外,Kladko等[32]開發了一種海膽狀磁性納米顆粒。附著在酵母細胞表面的磁性顆粒可經低頻磁場(100 Hz)驅動而改變空間排布,在不影響細胞生存的范圍內改變細胞膜通透性,使釀酒酵母利用葡萄糖生產乙醇的轉化率提高了150%。隨著對雜交系統的電子轉移機制及細胞能量代謝方式的深入研究,半導體合成生物學將為清潔能源開發、大宗化學品生產及高值藥物合成等提供重要技術支撐。
2 依托半導體-智能細胞的新型生物傳感
生物傳感器(biosensor)是一種可識別生物分子并將代謝物濃度轉化為光、電等信號的檢測儀器。它以生物敏感材料為識別元件,以可響應光、電、壓力、場效應等信號的材料為理化換能器,實現生物信號的轉換與輸出。自1967年第一個生物傳感器——葡萄糖傳感器[33]問世以來,科學家們以酶、抗體、細胞器、動植物組織等為特異性響應元件,開發出多種生物傳感器。傳統生物傳感器靈敏度高、特異性好,但功能有限。活細胞-半導體材料雜交體系可通過胞內生化反應實現對外環境的實時監測,且可對外界信號做出反應,從而執行普通生物活性材料無法完成的功能。這種新型智能生物傳感器在個性化診斷、疾病治療、微觀生物致動機器人開發等方面發揮重要作用。
生物標志物是指器官、組織或細胞結構功能發生改變的一類生化指標,為疾病診斷提供確切依據。生物標志物的檢測通常是將樣本(包括血液、體液、組織液等)取出后在體外進行,樣本穩定性及檢測時效性對結果有一定影響。為此,Mimee等[34]制備了一種可攝入的微型生物電子器件(IMBED),用于診斷胃腸道疾病。在該設備中,工程益生菌與半導體微電子系統形成的雜交系統共同集成于一種可吸收的微型膠囊中[圖2(a)]。當患者攝入該膠囊后,血紅素可透過半透膜與細胞接觸,并借助細胞外膜轉運蛋白ChuA進入胞內與轉錄阻遏物HrtR互作,表達細菌熒光素酶操縱子luxCDAB。工程菌將血液信號轉換為生物光信號,隨后由光電探測器轉換為電信號,并從設備無線傳輸到外部無線電或蜂窩電話,用于讀取和分析。裝載不同工程菌的設備可對血紅素、酰基高絲氨酸內酯(AHL)以及硫代硫酸鹽等腸道炎癥的相關生物標記物產生響應,為胃腸道疾病檢測提供一種快速、微創且經濟有效的檢測手段。除了用于個性化診斷,智能化生物傳感器的信號接收器和轉換器的定位并非一成不變。當半導體材料成為信號接收載體,活細胞就能以此調節自身功能從而實現疾病精準治療。葉海峰團隊[35]將電子設備生成和讀取數字信號的能力與光遺傳工程細胞相結合,通過智能手機無線調控工程菌在小鼠體內生產胰島素或胰高血糖素樣肽1(shGLP-1),推動糖尿病細胞療法的臨床應用。他們以一個32位的嵌入式微處理器芯片作為系統的智能控制器,在接收電子指令后觸發生物相容性的遠紅光源(FRL),并激活細菌的環二鳥苷酸(c-di-GMP)合酶BphS和c-di-GMP特異性磷酸二酯酶YhjH,從而實現基因表達調控[圖2(b)]。通過該半自動生物傳感平臺,采用手機App即可實現遠程控制糖尿病患者血糖,顛覆了傳統口服和藥物注射治療糖尿病的方法。
DNA是一種天然生物高分子,可通過堿基互補配對完成精準高效自組裝,也可響應不同刺激改變自身結構。因此,DNA作為結構基元可合成多種功能材料。DNA水凝膠不僅包含水凝膠的骨架結構,而且具有DNA的生物功能。基于DNA構象變化的可逆性,可實現水凝膠溶液狀態和凝固狀態的可逆調控,體現了材料結構與功能的完美融合[36]。仰大勇課題組[37]提出了構建超軟動態DNA水凝膠的新策略,制備了一種能對不同極性溶劑快速而靈敏響應的DNA/多巴胺接枝葡聚糖軟體水凝膠,并成功利用這種DNA水凝膠導線電路控制釀酒酵母發酵過程[圖3(a)]。此外,該課題組還基于滾環擴增(RCA)方式合成了具有形狀適應性的DNA水凝膠機器人。在遠距離磁力驅動下,該機器人可通過快速變形和恢復形狀來實現狹窄和結構復雜環境中的導航。具有良好生物相容性的DNA機器人成為智能運輸活細胞的工具,可在體內診斷治療、植入式醫療和微創手術等方面執行復雜任務[圖3(a)][38]。Hamada等[39]受新陳代謝的啟發,提出DASH機制——基于DNA的層次材料組裝與合成。該理論認為,DNA分子可被合成并組裝為一種層級結構,它在液體環境中可攝取外界能量,并按指令自動進行生長與降解,從而得到一種由人工代謝提供動力的動態材料[圖3(a)]。這種材料有望用于開發新一代生物芯片或傳感機器人。
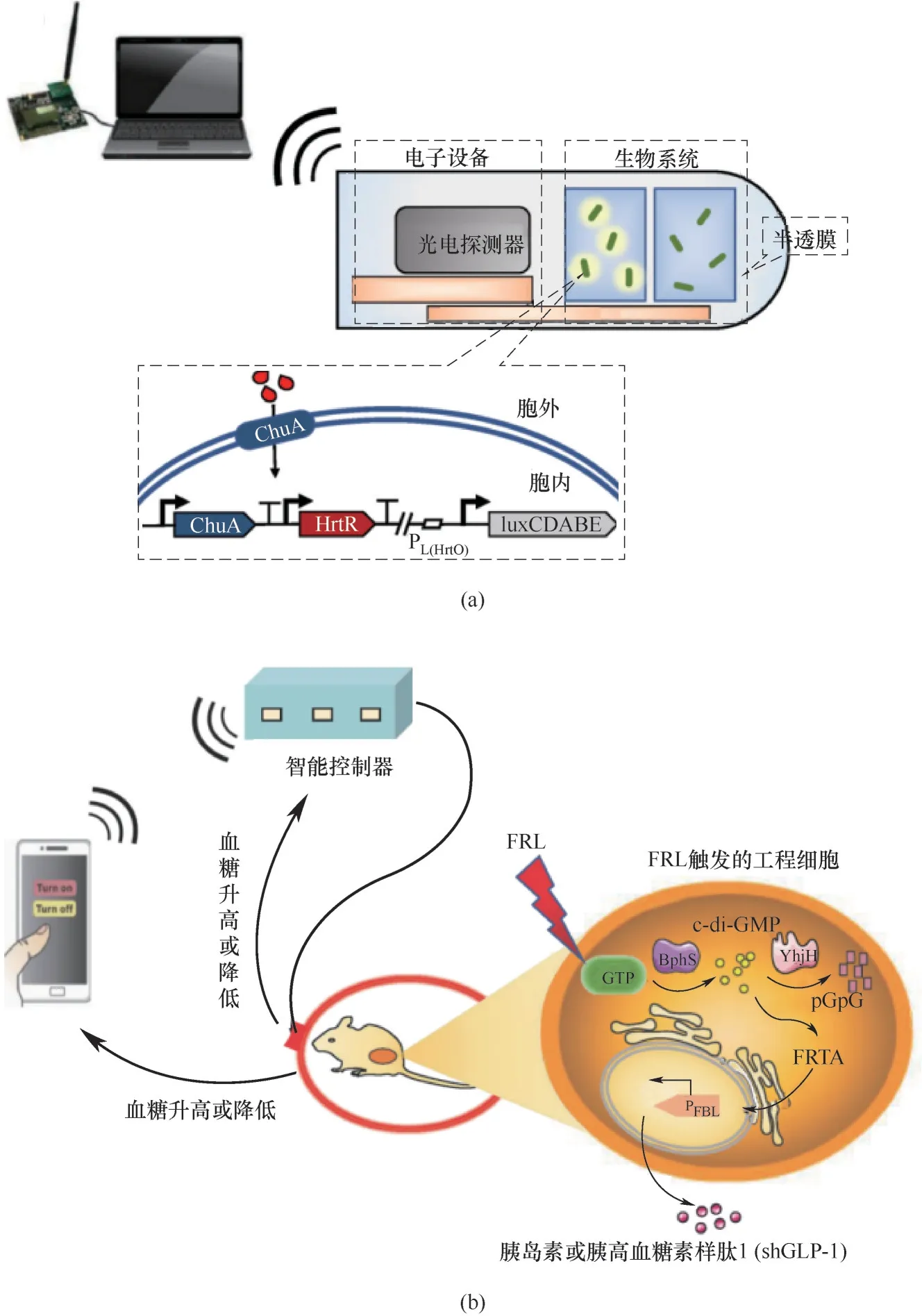
圖2 可吸收的微電子設備(IMBED)示意圖(a);智能手機調控工程細胞的表達,實現半自動血糖穩態(b)Fig.2 Schematic diagram of absorbable microelectronic equipment(IMBED)(a);Smart phones regulate engineered cells to achieve semi-automatic blood glucose homeostasis(b)
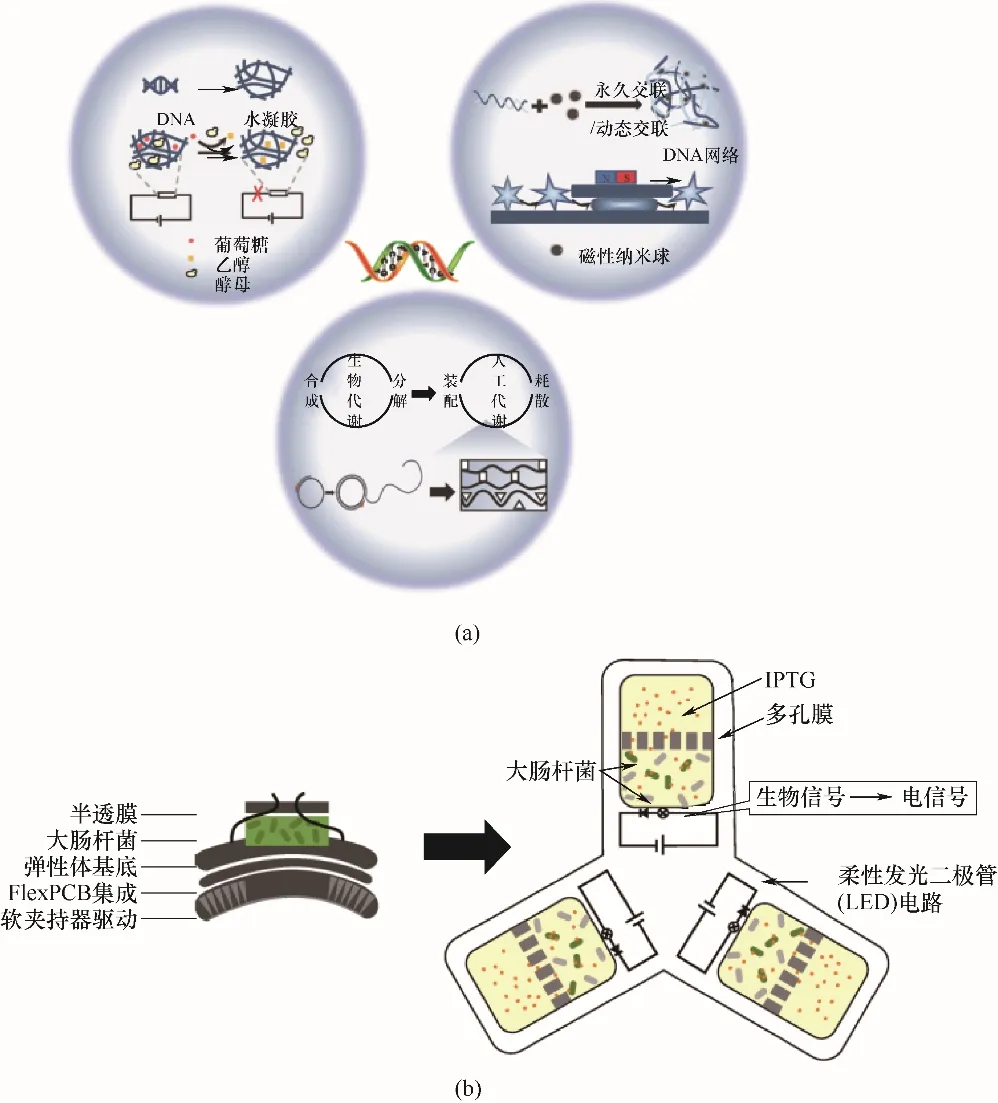
圖3 DNA水凝膠的應用(a);工程細胞-柔性材料軟機器人示意圖(b)Fig.3 Applications of DNA hydrogel(a);Schematic diagram of engineered cell-flexible materials-based soft robot(b)
當半導體-智能細胞與生物基材料耦合時,信號轉換與傳遞得到進一步擴展,一種新型生物混合機器也隨之誕生[40-42]。Justus等[43]用聚醚砜膜(PES膜)、聚碳酸酯軌跡蝕刻膜(PCTE膜)和多孔PDMSNaHCO3膜將重組大腸桿菌與嵌入式電子元件集成到密封的彈性體材料中[圖3(b)]。PES膜和PCTE膜孔徑分布均勻,可在截留大腸桿菌的同時允許化學刺激從外部環境傳輸到設備;多孔PDMS-NaHCO3膜具有彈性和光學透明性,可使工程菌在流體通道的液體介質中運動并輸送光遺傳信號。當生物傳感模塊中的菌株接受刺激并合成熒光蛋白后,嵌入式電子元件被激活,將生物信號轉換成電子信號并通過調整下一級層中水凝膠的狀態,來控制微型軟夾持器的抓取行為。這項工作真正賦予機器人的自主功能,為軟材料和集成界面機器人的研發開辟了新方向。
半導體器件的特點是響應速度快、可放大信號并具有一定容錯性,這些優點很適于調控生物系統的復雜動力學[44-45];而活細胞具有復雜的基因表達及代謝能力,能夠執行無機材料無法完成的工作。因此,二者結合將同時打破材料及生物系統等領域的技術瓶頸,具有廣闊應用前景。
3 基于DNA的大規模信息存儲
目前,數據存儲介質大多是性能良好的半導體材料。數據存儲密度與材料的物理尺寸以及電路集成能力密切相關。然而,半導體材料構成的電路已接近其性能極限;此外,亞納米水平的集成也因元件間距過小而面臨量子干擾、庫侖阻塞等物理效應的限制。因此,開發新的數據存儲方法才能解決當今信息太多而存儲能力弱的矛盾。分子數據存儲是一種密集且持久的信息存儲方式,但半導體材料需在極低溫度下(約-210℃)才能保持單分子級的數據存儲性能,昂貴的冷卻系統限制了該方法大規模應用。DNA作為生物遺傳信息的載體,其獨特的生物學特性使其在存儲時間、存儲密度和數據讀取等方面具有優勢。
首先,DNA具有雙螺旋結構,脫氧核苷酸外側通過共價的3,5-磷酸二酯鍵結合,內部則通過堿基互補配對形成大量氫鍵。DNA的穩定結構確保其較長的半衰期,因而可長期甚至永久存儲數據[46-48]。第二,DNA具有極高的數據存儲密度。DNA分子中的堿基分為腺嘌呤(A)、鳥嘌呤(G)、胞嘧啶(C)和胸腺嘧啶(T)四種(以A-T,C-G方式配對,形成堿基對,bp),它們類似于計算機二進制代碼中的“0”和“1”[49-50]。與二進制代碼相比,DNA序列由于堿基種類的倍增而可容納更多信息。例如,對于一段長度為X的字符串,二進制系統只能包含2X倍信息,而含四種堿基的DNA則可容納4X倍信息,即單個脫氧核苷酸可表示兩個比特的信息[51]。此外,單個脫氧核苷酸分子的質量極小(約為4.982×10-22g),理論上每克單鏈DNA能夠存儲高達455艾字節的數據(1艾字節=260字節≈1.15×1018字節)[52]。當這些龐大數據信息存儲在大腸桿菌基因組DNA(5.44×106bp)時,卻只占用不到1 cm3的空間。電子顯微觀察發現,約200 bp的DNA分子被壓縮在直徑為10 nm的核小體結構中。一個核小體的尺寸及其包含的遺傳信息量已經優于一個晶體管電路所具有的信息存儲能力。DNA在細胞內還可進一步纏繞形成染色體,最終完成近8400倍的壓縮。這意味著如果能成功利用DNA來存儲數據,那么當前的海量信息可包裝在一個0.00352 m3盒子里,約1 kg DNA即可滿足2040年世界的存儲需求(3×1024位)[51]。第三,DNA可通過聚合酶鏈式反應(PCR)方便快速且高保真地復制,這使得大規模數據備份變得簡單[53]。同時,日益成熟的全基因合成及測序技術為大片段DNA的數據寫入和讀取提供了保障[52,54]。
DNA數據存儲的基本過程包括:(1)編碼——將數字信息編碼為DNA序列;(2)合成——將序列寫入實際的DNA分子;(3)存儲——將合成的DNA片段保存在載體或細胞中;(4)訪問——檢索和選擇性讀取序列信息;(5)解碼——將測定的序列信息轉換回數字信息(圖4)[55]。
信息寫入需通過計算機算法將二進制代碼映射為DNA序列。由于保真度的要求及合成技術的限制,DNA序列不能無限延長,因此信息常被存儲于多個DNA片段。即每段DNA序列都被加入一些冗余信息,用于檢索和排序以確保信息的正確讀取。盡管額外的冗余有助于提高信息讀取的準確性,但同時也占用了空間,采用適當的編碼方案和糾錯策略可以平衡讀取準確性和數據冗余度之間的沖突[56]。在設計編碼方案時要充分考慮堿基比率的均一性,從而避免特殊序列(如GC%過高或包含大量重復片段)對擴增效率及測序準確率的干擾[57-58]。此外,目前的DNA合成和測序技術仍難以保證100%的準確率,所以糾錯策略必不可少[59]。最直接的糾錯方法就是添加冗余信息,用于提高在數據丟失或錯誤情況下仍能檢索到原始信息的概率。冗余越多,存儲結果對錯誤的容忍度越高。例如,Blawat等[60]使用前向糾錯方案實現對DNA中22 MB數字數據的存儲和無錯誤檢索。Erlich等[58]報道了一種魯棒性很強的“DNA噴泉碼”的存儲策略,該策略在DNA寡核苷酸中存儲了2.14×106字節的內容,并能夠從相當于Illumina測序儀的一個測序覆蓋率中完美地檢索到信息。迄今為止,若干課題組建立了多種糾錯方法以適應不同存儲情況[52,61-62]。除了編碼過程有嚴格要求,隨機訪問能力也是DNA數據存儲面臨的一項重大挑戰。從計算機科學的角度來看,預計存儲的數據將具有隨機訪問權限。缺乏隨機訪問會阻礙數據容量的擴大,當檢索少量數據時,對整個數據集進行排序和解碼是不切實際的[56]。目前比較流行的訪問DNA數據的方法是磁珠提取和PCR擴增。Baum[63]基于分子探針原理,展示了如何利用磁珠調取已做好標識符的數據項。基于PCR的隨機訪問由于使用數據片段對應的唯一引物,因而具有較高特異性[64]。Organick等[65]研究發現,當樣本池的規模達到TB級(1012字節),PCR方法仍可準確調取所需數據,但該規模不足以滿足未來分子數據存儲的要求。由于被調取的DNA片段需經測序解碼來得到原始信息,所以測序能力直接影響該存儲方式的發展。目前使用最廣泛的是Illumina公司開發的DNA測序平臺。該平臺基于合成測序和圖像分析的概念,以熒光斑點顏色指示序列中的各個堿基,精確的光學捕獲裝置和圖像處理技術有助于提高測序通量[66-67]。另一是牛津納米孔技術公司(ONT)商業化的納米孔測序技術,可在捕獲DNA后使其通過一個電壓箝位的納米級孔隙,不同堿基將導致孔隙電流出現相對應的微小波動,從而實時讀取序列數據[68]。
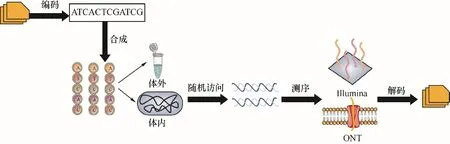
圖4 DNA數據存儲流程圖Fig.4 Flowsheet of digital information storage in DNA
DNA數據存儲方興未艾,未來面臨諸多挑戰。首先,當前DNA存儲的寫入吞吐量約為每秒千字節級別,與主流云存檔存儲系統的每秒億字節讀寫能力仍有數量級差距。其次,DNA分子的物理保存方式也需斟酌[69-70]。體內和體外存儲各有其優勢:在備份成本方面,雖然體內存儲比體外寡核苷酸文庫的合成更復雜,但細胞自主的DNA合成和糾錯更具成本優勢;在長期儲存方面,體內條件下的DNA降解度慢于體外,因此更適合長期保存數據[71]。隨著DNA合成技術精度的不斷提高,體外合成比體內復制過程中由突變造成的誤差更低。目前仍缺乏時間讀取短且空間占用小的存儲方法,DNA數據的讀取過程仍耗時長。未來新一代合成、測序及檢索技術將為DNA數據存儲帶來質的飛躍。
4 總結與展望
半導體合成生物學利用材料科學的工具和方法去發展和調控生物系統,同時基于合成生物學思路開發新材料。生物學、材料學、電子學和計算機科學等多學科交叉可促進生物學與半導體技術的日益融合,衍生出具有重要價值的研究課題。
(1)半導體材料能高效吸收轉化光能、磁能、化學能,活細胞則將能量用于自身代謝并合成高附加值產品。用于生物催化的半導體-細胞雜合體系是目前研究熱點[72]。本課題組致力于工業微生物的遺傳改造和調控,其中包括將還原性三羧酸循環及C4模塊導入大腸桿菌,使其固定CO2而生產化學品。對于大腸桿菌等非光合細菌,該途徑提供的還原力及能量不足,而細胞與半導體材料耦合可彌補該缺陷,從而提高CO2利用率。材料與細胞表面的接觸對細胞間原有信號傳遞、生物被膜等造成干擾。如何減輕或平衡能量傳輸對細胞的負荷和不良刺激,加速能量傳遞,是發展半導體生物催化的關鍵。此外,經半導體材料傳輸的能量在進入細胞后會遵循胞內能量代謝方式,缺乏可控性。未來,利用合成生物學手段開發胞內多模塊間正交能量分配或許是新突破。
(2)半導體器件可放大信號、具有一定容錯性且響應速度快,適于調控復雜生物系統[44-45];而活細胞具有精密的基因表達及代謝能力,能夠執行無機材料無法完成的工作。二者結合將同時打破材料及生物系統等領域的技術瓶頸。例如,通過課題組之間的協作,以極端微生物為底盤細胞,將其與多重納米材料相結合,以便監測與修復環境。其中的工程細胞可感知環境中污染物的濃度,而半導體材料傳感器則負責信息的反饋;之后,信息集成模塊根據污染物的種類,通過不同信號輸入激活工程菌中相應的污染物降解途徑,從而實現“自感知-自反饋-多樣化處理”的目標。目前,生物傳感器的應用已經從基礎的物質檢測擴展到了復雜的生物醫療。然而,相對于半導體響應元件,人們對活細胞功能的開發程度還較低。今后,將外部傳感信號與胞內如群體感應(quorum sensing)等自主交流方式相耦合,同時結合人工智能手段賦予細胞“感應-分析-指令-行動”等能力,有望使生物傳感技術邁向新臺階。
(3)DNA數據存儲是未來信息存儲的發展趨勢,其優點在于存儲密度大、易保存且受外界條件影響小。然而,DNA合成成本高以及讀取速度慢等缺點仍是挑戰。盡管如此,現有物種DNA圖譜復雜多樣,包含信息量巨大,若將DNA存儲的編碼算法與之耦合即可將生物基因組作為天然存儲單元,節約合成成本。此外,新一代測序技術助力DNA的精確和快速讀取。
未來半導體合成生物學仍將遵循“設計-構建-測試-學習”(design-build-test-learn)的閉環思路,在細胞、組織和系統各個水平不斷發展和優化生物-非生物雜合體系并拓展其應用。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
小學科學(學生版)(2018年3期)2018-04-18 12:34:19
知識經濟·中國直銷(2017年10期)2017-11-07 02:39:52
知識經濟·中國直銷(2017年3期)2017-04-16 03:08:01
文學少年(有聲彩繪)(2017年2期)2017-03-06 08:10:54
