洱海濕地昆蟲群落網絡結構與自適應性研究
2021-06-30 16:42:00郝鴻俊祝遠波羅桂蘭
大理大學學報 2021年6期
關鍵詞:物種
郝鴻俊,王 熙,張 梅,劉 璇,祝遠波,羅桂蘭*
(1.大理大學工程學院,云南大理 671003;2.大理大學數學與計算機學院,云南大理 671003)
昆蟲是數量最多的動物,占所有動物總數的73%〔1〕。昆蟲能在自然界營養循環等生態系統服務區域中發揮重要作用〔2〕,并可以作為監測濕地生態環境和生物多樣性的指示性物種〔3〕。濕地昆蟲種類繁多,群落結構也很復雜。韓爭偉等〔4〕對太湖濕地昆蟲群落結構及多樣性進行研究,利用多樣性特征指數等參數找到了影響太湖濕地昆蟲群落的重要因子為濕地植被類型和人工干擾。何云川等〔5〕對銀川4種濕地生境的陸生昆蟲群落多樣性與穩定性進行研究,對4種生境群落特征指數聚類分析,結果表明昆蟲群落多樣性隨季節時序變化。可見昆蟲對環境的變化十分敏感,環境發生變化將會導致昆蟲的數量和群落結構變化〔6〕。
洱海是云南省第二大淡水湖,北起洱源,長約42 km,周邊有豐富的濕地資源。但是隨著洱海周邊旅游業的開發,洱海濕地生態環境遭到了一定的破壞影響。目前洱海生態保護已經受到人們的廣泛關注,濕地建設工作也在緊張進行。本文選取洱海3個典型的濕地作為研究對象,3個濕地擁有不同的植被類型,受人為干擾程度也大不相同。通過調查和分析2018年11月至2019年10月不同生境下的昆蟲群落多樣性、均勻度、優勢集中性指數等,探討昆蟲多樣性的季節性動態變化和適應能力。該研究將為洱海濕地生物多樣性保護與濕地建設方案提供理論支持和數據分析樣本。
1 研究區域與昆蟲環境樣本數據
洱海位于云南省大理市郊區,面積約251 km2,總蓄水量30億m3。大理屬亞熱帶高原季風氣候,是孟加拉灣西南季風帶的迎風區,所以風較大,紫外線較強,光照充足,年平均氣溫15.7℃,年均降雨量1 000~1 200 mm,降水充沛〔7〕。
本文選取的3個濕地為洱源東湖濕地、洱海月濕地和羅時江濕地。濕地信息如表1所示,各區域環境差異較大。
本文數據樣本由國家自然科學基金項目“洱海濕地昆蟲群落普適生境網絡生態行為研究”提供。該項目的主要研究目標是通過構建昆蟲生境關系網絡,探討各個生境因子之間、生境因子與昆蟲群落結構之間、人類行為與生境因子之間的相關性。目前已經完成了昆蟲生態網絡模型的定義,生成了3個濕地的昆蟲群落網絡拓撲圖,并對網絡特征參數進行了分析。本文將會在原有昆蟲生態網絡模型的基礎上進行改進,生成2018年11月至2019年10月的昆蟲生態網絡關系拓撲圖。樣本采集信息見表2。
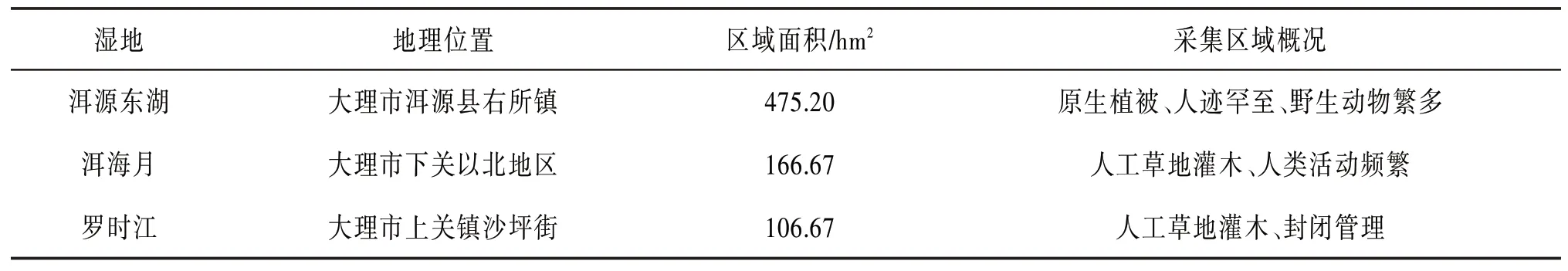
表1 洱海濕地信息
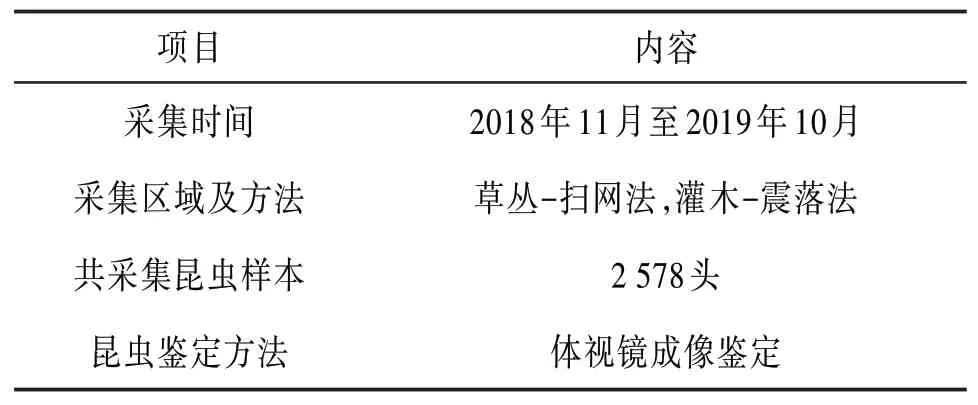
表2 昆蟲數據樣本采集信息
昆蟲捕捉采用自制捕蟲網,網口直徑約33 cm。草叢每5 m進行一次掃網,一共掃20網。采集到的昆蟲將其置于鑒定盤里進行初步分類。初步分類后將會對采集物種進行標號并置于離心管中保存。離心管中注入乙醇防止昆蟲尸體毀壞。最后,根據昆蟲分類法〔8〕利用體視鏡成像鑒定出昆蟲的綱、目、科、種等信息。鑒定信息匯總后得到各個濕地不同采集區域每個月的昆蟲樣本數據。體視鏡成像照片標號與昆蟲樣本標號對應,方便后期數據核查。
環境數據包括氣象數據和生境植被數據。其中氣象數據是通過放置生態環境監測點,周期性采集了溫度、濕度、降雨量、光照等10余項環境數據樣本。生境植被數據通過濕地調研、取樣鑒定,共鑒定出60余種植被種類。3個濕地的植被種類見表3。
洱源東湖濕地作為人為干擾最小的濕地,將其作為另外兩個濕地植被種類的對照組具有較強的意義。

表3 濕地植被種類
3個濕地鑒定出的植被種類數量相當,但是洱海月濕地的灌木、喬木種類及數量占比要高于洱源東湖濕地,其原因是洱海月濕地為人為建設的濕地保護區,目前作為濕地公園向游客開放。濕地內的大部分植被都為外來引進。灌木、喬木不僅具有生態功能,同時還具有為人類遮陽防風等功能,并且作為濕地生態景觀,其草本植物種類雖然較少,但草本植被分布均勻,空間結構較好。
羅時江濕地雖然也是人為建設的濕地保護區,但目前已封閉管理。該濕地最主要的功能為生態保護。植被類型多為草本植物,其數量與洱源東湖相當。相比另外兩個濕地,該濕地灌木、喬木分布較稀疏,所以視野較開闊,地面光照較充足。
洱源東湖濕地作為最原始的濕地保護區,其植被類型少有外來引進。由于沒有人為干擾,該區域植被分布較為緊密。
2 昆蟲群落結構參數描述與定義
參數1 相對多度Pi指第i個物種個體數占總個體數的比例。其中,M i表示第i種物種的個體數,M表示昆蟲群落內采集的物種總個體數。計算公式為

參數2 物種多樣性指數H′表征昆蟲對一定環境的適應性和物種在特定空間范圍里的演化程度,也表征昆蟲群落內所擁有物種種類數目多樣的程度〔9〕。計算公式為
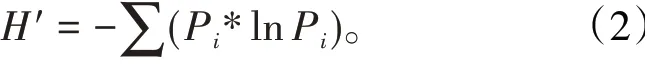
參數3 物種豐富度指數lnS,S為該區域采集到的物種數,豐富度指數lnS越大,表明該區域的物種種類越豐富,反之物種就越單一〔4〕。
參數4 物種均勻度指數J指所有昆蟲個體在群落中的分布情況。計算公式為

群落穩定性與物種均勻度指數密切相關,J越高表明昆蟲群落的穩定性越強〔10〕。
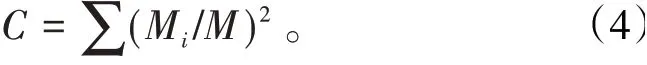
參數5 優勢集中性指數C用來描述昆蟲群落組成狀況。計算公式為C反映了昆蟲群落內物種發展的優勢,C越大,群落中的個體數越集中在少數類群上,H′就會越小〔11〕。
參數6 昆蟲群落穩定性指標,計算公式為

S s∕S i反映種間數量上的制約作用,比值越大,相對穩定性越強。S n∕S p反映食物網關系的復雜程度和相互制約的程度,比值越大,天敵物種數所占比例越大,群落穩定性越強〔12-13〕。
定義1 優勢類群指昆蟲群落中更適應環境的物種,即個體數占采集總個體數比例大于10%的物種〔14〕。
定義2 常見類群指昆蟲群落中常見的物種,即個體數占總采集個體數比例大于等于1%的物種〔14〕。
定義3 稀有類群指在昆蟲群落里很少采集到的物種,即個體數占總采集個體數比例小于1%的物種〔14〕。
定義4 昆蟲群落時序演化結構指昆蟲群落的物種數、個體數、均勻度等參數隨時間(如季節等)變化所表現出來的動態結構。
3 昆蟲群落網絡拓撲結構建模
在文獻〔3〕原有昆蟲生態網絡模型中,根據復雜網絡理論,以昆蟲種群為網絡節點,以每月采集的昆蟲物種的個體數量構建長度為12的時間序列,分別求出兩個物種間個體數量變化的皮爾遜相關系數,并根據相關系數的數值大小定義邊的權重,最后使用Gephi軟件〔15〕建立昆蟲生態網絡節點和邊的數據資料庫,以此生成昆蟲生態網絡關系拓撲圖〔3〕。
為了降低由于采集人員不同帶來的誤差,本文在原有昆蟲生態網絡模型的基礎上做出改進。以每個月昆蟲種群所占該區域采集昆蟲個體的數量比例構建時間序列,求出各物種間數量比例的相關性作為邊。構建出的昆蟲生態網絡模型拓撲圖的節點面積大小表征某區域昆蟲種群大小,節點顏色深淺表征節點度大小。邊表征昆蟲數量比例間的相關性。生成的3個區域的昆蟲生態網絡關系拓撲圖見圖1~3。
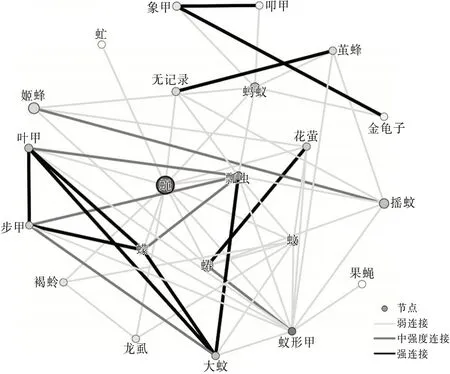
圖1 洱源東湖濕地昆蟲生態網絡關系拓撲圖
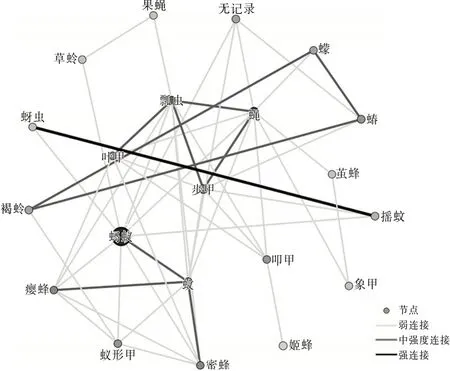
圖2 洱海月濕地昆蟲生態網絡關系拓撲圖
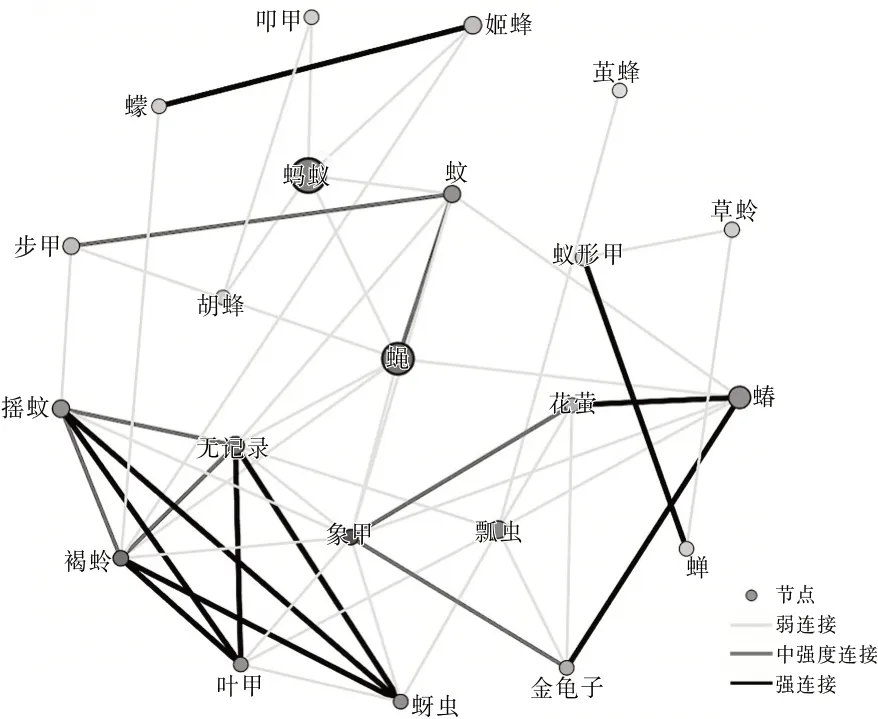
圖3 羅時江濕地昆蟲生態網絡關系拓撲圖
從3個區域的網絡關系拓撲圖可以看出蠅的節點顏色較深,可見蠅和其他物種的相關性比較強,是網絡中的中心(HUB)節點。這里的HUB節點的節點度值較高,表征該節點與其他節點連接緊密。果蠅在洱源東湖和洱海月濕地中都為低節點度節點,并且在羅時江濕地中沒有捕獲到該物種。草蛉在洱海月和羅時江濕地中都為低節點度節點,并且在洱源東湖濕地中沒有捕獲到該物種。這類型節點與其他物種的相關性較低。同時有明顯特點的還有螞蟻節點,洱海月和羅時江的螞蟻種群都比較大。可見蠅、螞蟻都是濕地昆蟲生態網絡的關鍵物種。
根據文獻〔3〕計算方法得到網絡特征參數。平均加權度:洱源東湖>洱海月>羅時江;平均聚類系數:羅時江>洱海月>洱源東湖。可見:洱源東湖濕地昆蟲群落的昆蟲節點連接最豐富,關系最密切,網絡的魯棒性較好。羅時江節點之間的相互影響力強,當網絡節點種類、數量發生變化時能夠迅速做出調整,具有較強的調節能力。
4 昆蟲群落結構及特征分析
4.1 昆蟲群落組成分析 3個濕地在一年內共采集到2 422頭昆蟲,隸屬于11目。3個濕地在一年內均采集到約30種昆蟲類群,其中約2∕3的昆蟲類群在3個濕地均有采集到,但是3個濕地昆蟲目個體數所占比例差異較大。3個濕地昆蟲群落結構組成成分見表4。
根據表4可知:洱源東湖濕地的雙翅目、膜翅目、半翅目和鞘翅目昆蟲類群,洱海月濕地膜翅目、鞘翅目和雙翅目昆蟲類群,羅時江濕地雙翅目、膜翅目、鞘翅目和半翅目昆蟲類群,均超過所在區域總數量的10%。
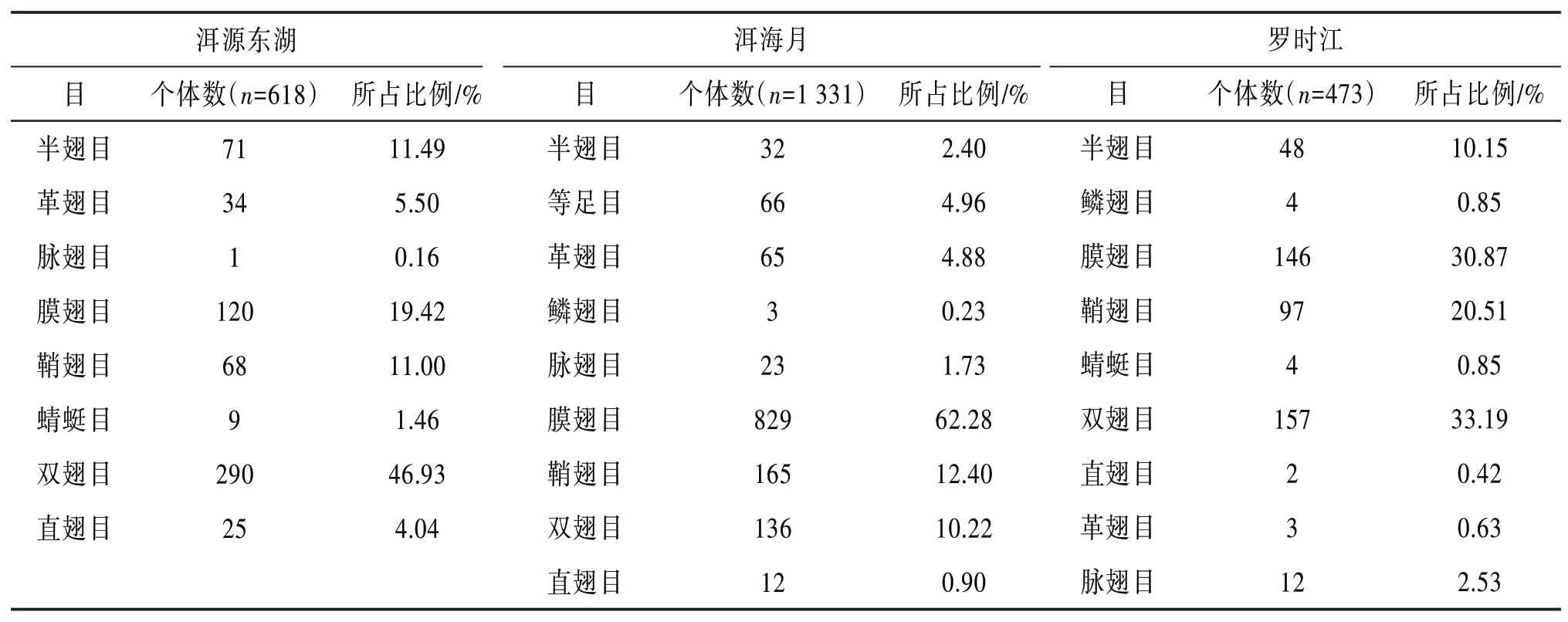
表4 昆蟲群落結構組成成分
表5為一年中3個濕地昆蟲群落的物種占比表。
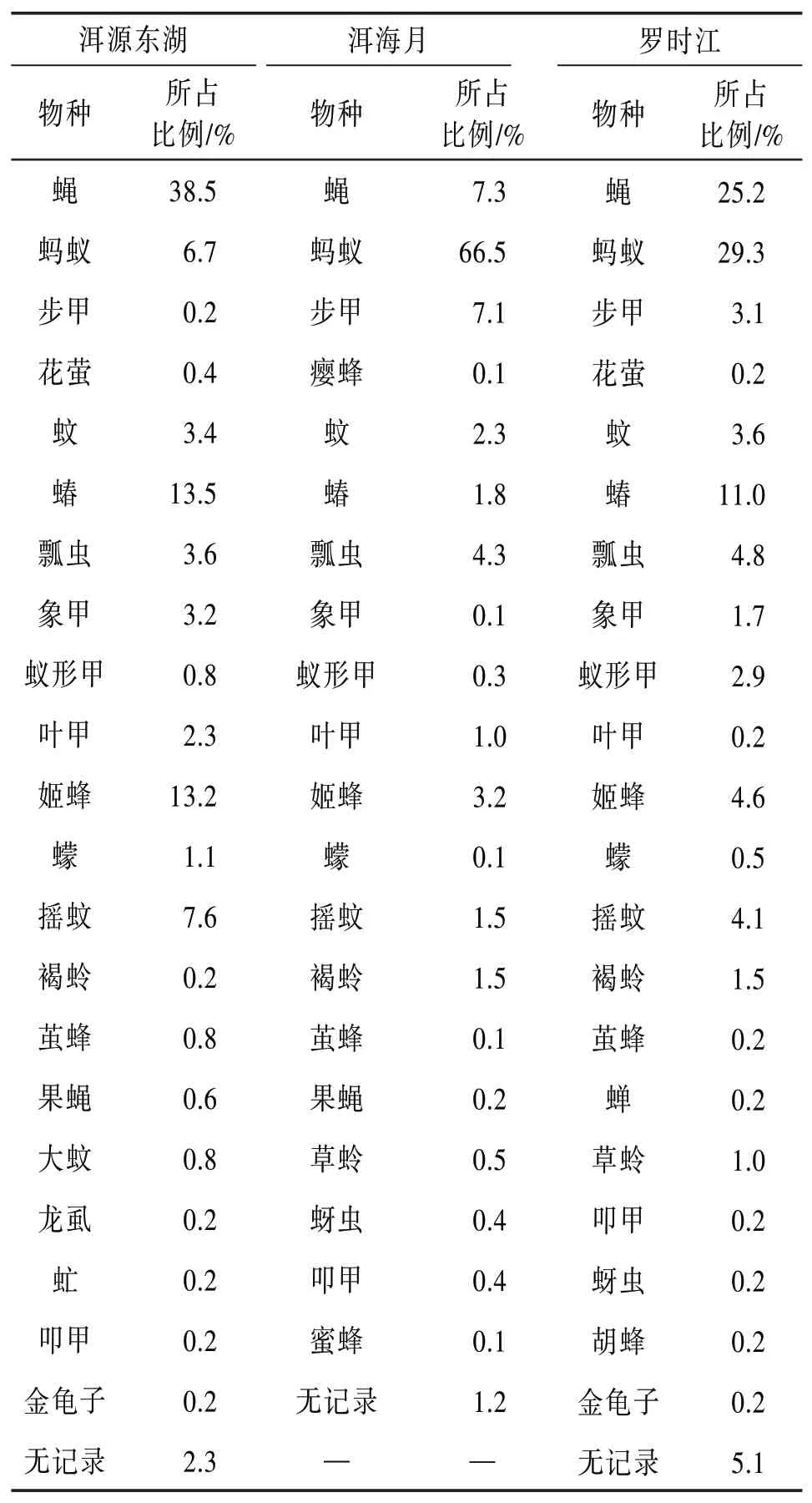
表5 洱海濕地昆蟲群落物種占比
根據定義1~3,再結合表5所示結果,整理出3個濕地的種群類別見表6。
由表6可知,蠅是洱源東湖和羅時江的優勢類群,但卻是洱海月的常見類群。常見蠅類多以腐敗的動植物以及人與動物的食物、排泄物等為食。洱源東湖濕地最接近原始生態,沒有人為清理腐敗植物,沒有占領野生動物的活動區域,區域內有大量動物的糞便,這為蠅的生存繁殖提供了很好的場所。而羅時江和洱海月的植被都有人為定期修剪清理,并且洱海月的人類活動頻繁,野生動物相對較少。這些因素導致蠅在3個濕地的數量占比差距懸殊。同理:螞蟻是洱海月和羅時江的優勢類群,但卻是洱源東湖的常見類群。螞蟻通常生活在干燥的區域,而大理降雨充沛,濕地多有積水。洱源東湖由于沒有人工干預,多為坑洼的泥濘區域,容易積水。而另外兩個濕地由于人工定期對草地灌木修整,陽光更容易照在地面上,地面水分蒸發較快,為螞蟻提供了更好的棲息場所。
綜上分析,人為干擾對昆蟲群落結構造成了巨大的影響。優勢類群決定了影響該區域生態環境的直接因素;常見類群反映出影響該區域生態環境的主要因素,該類群維持著區域生態環境的穩定性;而稀有類群表征了昆蟲的多樣性,反映出該區域生態環境在遭受到破壞以后的系統自我修復能力。
4.2 昆蟲群落結構特征參數比值分析 取3個濕地一整年的昆蟲群落數據樣本,根據式(2)~(4)計算出洱海濕地昆蟲群落的多樣性特征參數,對比分析濕地昆蟲群落的多樣性及穩定性。以洱源東湖特征參數為基準“1”,其余兩個濕地特征參數與洱源東湖特征參數比值的柱狀圖見圖4。

表6 昆蟲種群類型
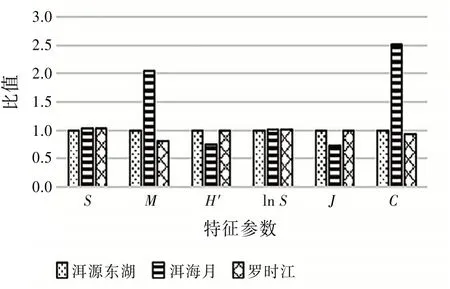
圖4 3個濕地特征參數比值柱狀圖
由圖4可見:物種多樣性指數H′和物種均勻度指數J規律保持一致,均為洱源東湖>羅時江>洱海月。而優勢集中性指數C表現為洱海月>洱源東湖>羅時江。其中洱源東湖和羅時江的各個參數相差較小,與洱海月的參數相差較大。由于各個濕地采集到的物種種類數目近似,所以物種豐富度指數lnS也近似。各參數表明洱源東湖和羅時江濕地的昆蟲群落物種豐富,物種數量也相對均衡,沒有極端優勢物種,與洱海月的昆蟲群落比較要相對穩定。
羅時江濕地經過人為建設保護,種植了特定的植物。但是由于后期封閉管理少有人員活動,所以昆蟲群落結構良好,與原生濕地洱源東湖相差無幾,可見人工修復濕地能起一定的作用。但是由于人類活動頻繁,導致洱海月濕地昆蟲群落結構較為脆弱,所以人為干預濕地更應重視昆蟲多樣性保護,提高生態系統的自我修復能力。
4.3 昆蟲群落時序演化結構分析 根據昆蟲生態環境數據樣本,按照式(1)~(4)計算,統計分析得到昆蟲群落結構特征參數隨時間動態演化趨勢。
由圖5可見:3個濕地全年的多樣性指數大多集中在1.0~2.0這個范圍內,其中最大值出現在9月(2.365 1)的洱源東湖濕地,最小值出現在5月(0.665 1)的洱海月濕地。洱源東湖濕地的多樣性指數變化趨勢相對平緩,從2018年11月(0.893 6)開始緩緩上升,說明該區域的昆蟲對生態環境的適應力強,只有在2019年4月(1.595 5)和2019年8月(1.538 9)發生明顯的下降趨勢。從物種來看,洱源東湖4月采集的物種以膜翅目和雙翅目為主,沒有采集到以蝽為主的半翅目。蝽的第一代成蟲一般出現在5月,第二代至第四代分別在6月上旬、7月中旬和8月中旬出現。蝽成蟲的壽命多為30~50 d。5月至8月采集到的蝽數量分別為3、13、12、31頭。這導致在8月蝽成了優勢類群。短期內對昆蟲群落的多樣性造成了一定的影響。
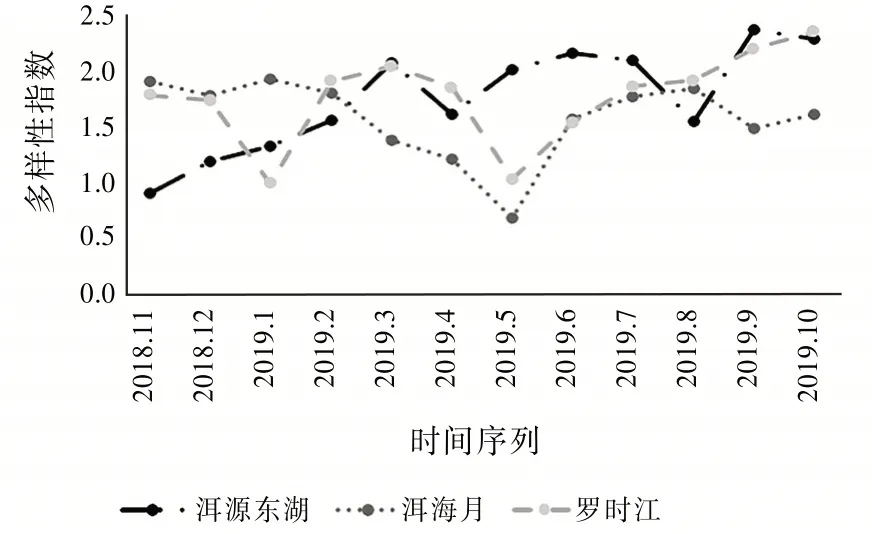
圖5 多樣性指數變化趨勢
洱海月全年的多樣性指數變化呈“V”形變化趨勢,在2019年5月(0.665 1)達到全年的最低值,但是在2019年6月之后逐步上升至上半年的平均水平,說明洱海月濕地具有一定的自我調節能力,昆蟲群落在受到一定的干擾破壞之后又能緩慢恢復。羅時江全年的多樣性指數呈“W”形變化趨勢,分別在2019年1月(0.976 3)和5月(1.013 5)到達谷底。說明羅時江濕地的昆蟲群落結構變化率較大,但即便如此在全年的大多數時間里,羅時江濕地的多樣性指數都高于洱海月濕地。羅時江濕地因為有更豐富的昆蟲物種,即使在外在因素導致群落發生變化時也能迅速做出自我調整,從而保持對該區域環境的自適應能力。
圖6的均勻度指數和圖5的多樣性指數的變化趨勢相似。其中,洱海月的均勻度指數常年都低于另外兩個濕地,說明該區域內的昆蟲群落穩定性相對較弱。具有自然生態環境的洱源東湖變化趨勢最平緩,可見原生的生態環境對昆蟲群落穩定性的維持起著重要作用。
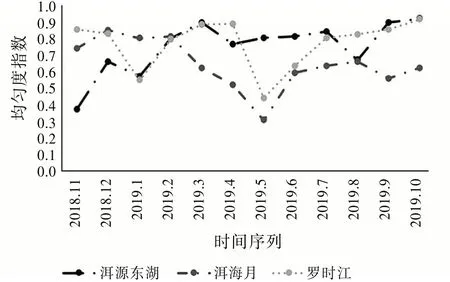
圖6 均勻度指數變化趨勢
圖7的豐富度指數能直觀反映采集到的物種數量變化。其中洱源東湖和洱海月的豐富度指數在2018年11月至2019年3月變化趨勢雷同,在該時間段豐富度指數波動明顯,并且整體有所下降。而羅時江的豐富度指數變化趨勢雖然和另兩個濕地近似,但是卻滯后了一個月,這可能與羅時江溫度低于其他兩個濕地有關。在2019年4月至2019年10月洱源東湖濕地和羅時江濕地變化趨勢近似,分別在6月至8月有一定的下降,而在8月之后又有所上升。可見,洱海濕地在冬季氣溫較低時物種豐富度指數較小,并且波動明顯,而在5月至9月溫度較高的季節,物種豐富度指數保持在較高的水平。該結果表明,豐富度指數反映了昆蟲群落受季節變化影響明顯。
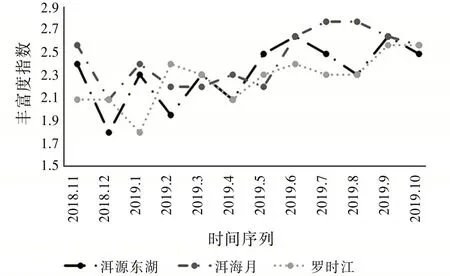
圖7 豐富度指數變化趨勢
同樣明顯受季節變化影響的還有昆蟲數量。從圖8可以發現:冬季溫度較低,在2018年12月3個濕地的溫度都達到全年的最低值。與此同時,3個濕地采集到的總昆蟲數量也達到全年的最低值。而在4月之后,3個濕地均超過時間序列年平均溫度,昆蟲的數量開始增多。特別地,洱海月濕地采集的昆蟲數量巨幅上升,這是因為采集到的螞蟻數量增多。從2月的14頭、3月的34頭、4月的53頭到5月的142頭、6月的94頭、7月的113頭。可見,螞蟻數量的激增是導致洱海月濕地昆蟲群落參數變化的主要原因。
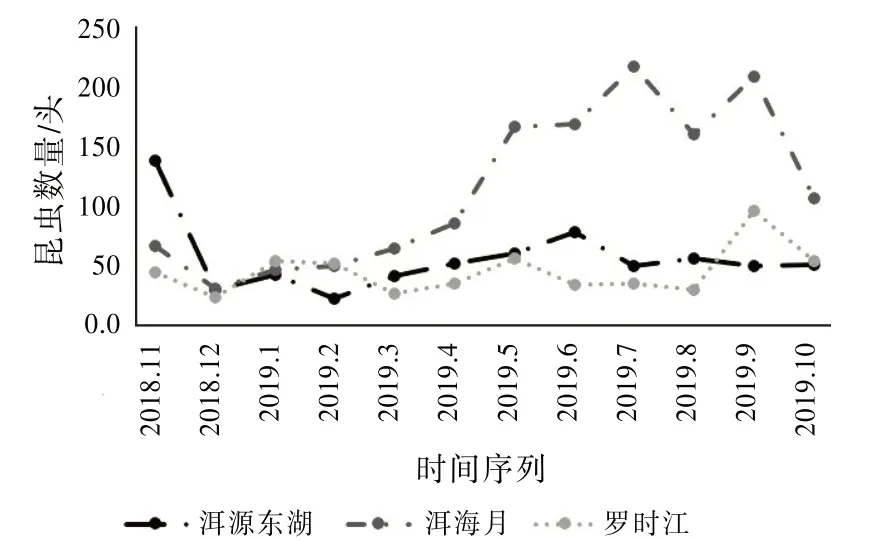
圖8 昆蟲數量變化趨勢
由式(4)知,優勢集中性指數和昆蟲群落結構穩定性成反比。由圖9發現:2019年5月洱海月濕地昆蟲群落優勢集中性指數達到最高0.729 0,這反映出該時間段內該區域出現了極端優勢物種,也就是圖8中提到的螞蟻,此時洱海月昆蟲群落穩定性最弱,正好對應圖6該月均勻度指數達到最小值。同理不難發現:羅時江濕地在該時間段的優勢集中性指數也達到全年最高0.598 2,其原因也是螞蟻數量的激增,但是增長率相對于洱海月濕地要小。對應圖6也反映了此時羅時江昆蟲群落穩定性較弱。
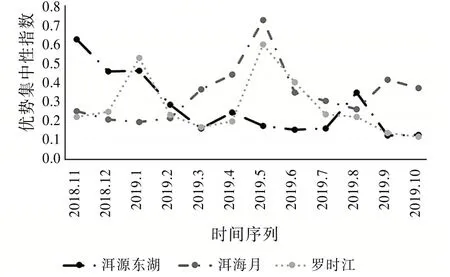
圖9 優勢集中性指數變化趨勢
4.4 昆蟲群落結構特征參數相關性分析 為進一步研究各個特征參數間的關系以及對昆蟲群落穩定性及多樣性的影響,這里引入相關系數R〔16〕,考察一年內各個特征參數時間序列間的關聯程度。相關系數值介于-1和1之間,其中越靠近1說明正相關性越強,越靠近-1說明負相關性越強。得出的相關系數見表7。
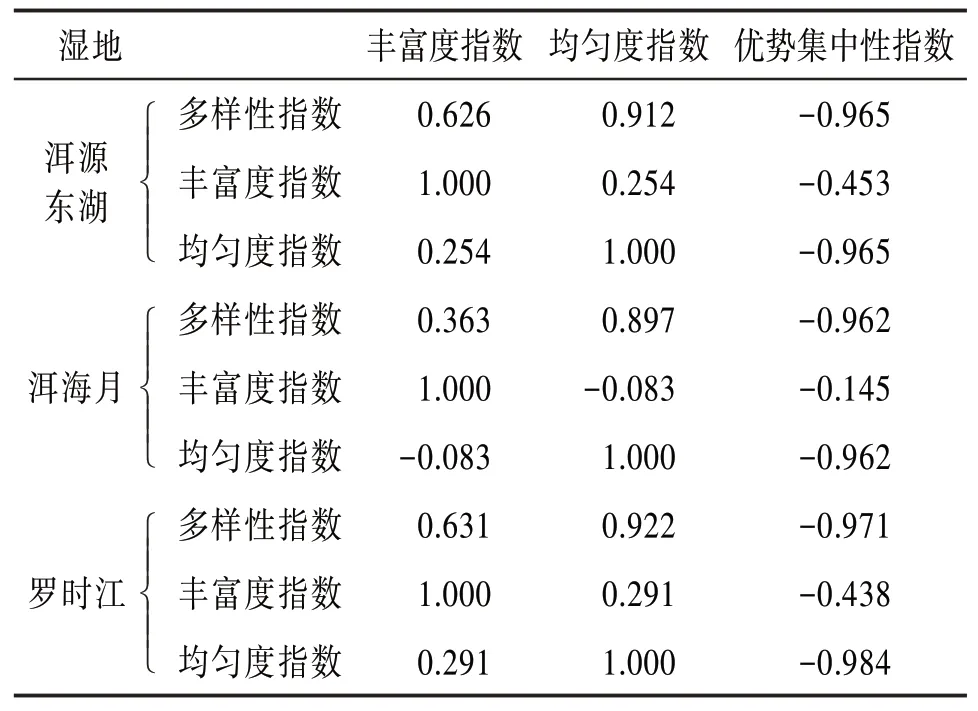
表7 特征參數相關系數表
由表7可見,3個濕地的多樣性指數與均勻度指數的相關性均為0.9左右,具有較強的正相關性,說明洱海濕地昆蟲群落的多樣性指數與均勻度指數關系密切。而3個濕地的優勢集中性指數分別與多樣性指數和均勻度指數的相關性均小于-0.9,具有較強的負相關性。
綜上分析,對于洱海濕地昆蟲群落保護,首先要保證物種的豐富度,特別是稀有類群,稀有類群的多少直接決定了該濕地的昆蟲生態網絡的大小。較大的昆蟲生態網絡具有較好的魯棒性。而優勢類群則很大程度上限制了昆蟲群落的多樣性。所以洱海濕地昆蟲群落的保護策略是:優先保護瀕臨消失的類群,重點監控優勢類群的發展。
4.5 生境因子對昆蟲群落結構特征參數影響分析根據3個濕地環境監測點采集的氣象數據,分別求出昆蟲采集前后3 d內的溫度、環境濕度、紫外線瞬時值、降雨量、風速的平均值,并構建時間序列。考察一年內生境因子與昆蟲群落結構特征參數的相關性。分別得出3個濕地生境因子與該濕地特征參數的相關系數的絕對值,并求出平均數。最后得到的相關系數見表8。
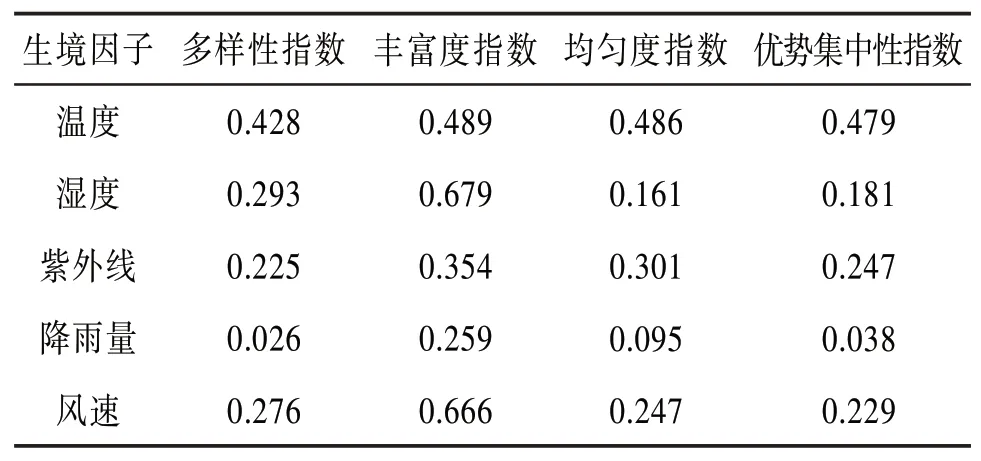
表8 生境因子與特征參數相關系數表
由表8可知,濕度和風速與豐富度指數的相關性達到了0.6以上,這可能與大理的地理因素有關,大理地處亞熱帶高原季風氣候區,常年日照充足,水分蒸發量大,氣候干燥;晝夜溫差大,常有大風天氣。其中,昆蟲群落豐富度指數隨著濕度的增加而降低,隨著風速的降低而增加。濕度越大物種越豐富,而風速過大會導致飛行類昆蟲的種類數量降低。
同時,溫度與昆蟲群落結構的特征參數更為密切,各相關系數都達到了0.4以上。圖10展示了昆蟲采集前后3 d內的日平均溫度。
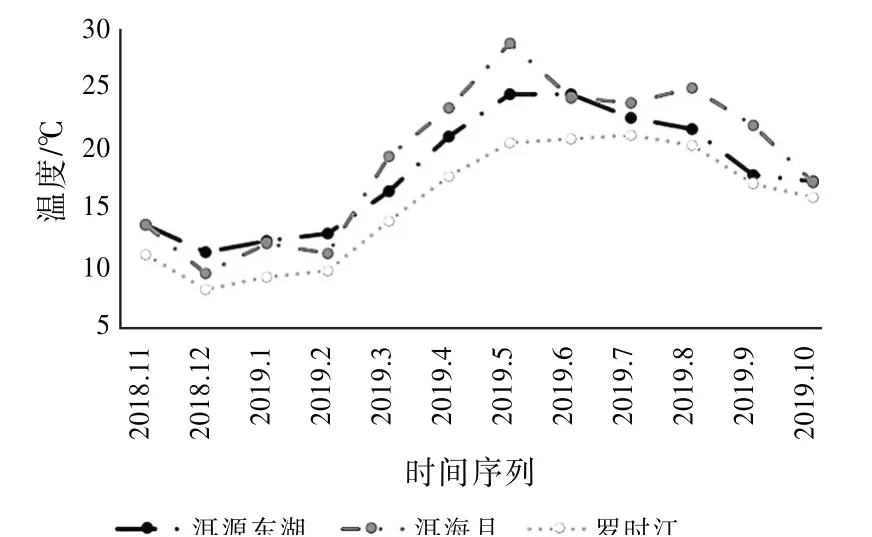
圖10 濕地溫度變化趨勢
可以看出羅時江濕地的溫度要低于另外兩個濕地,洱源東湖濕地的溫度在大部分時間介于洱海月濕地和羅時江濕地之間。洱源東湖濕地和羅時江濕地的溫度變化趨勢保持一致,溫度變化曲線相對平緩,而洱海月濕地的溫度變化較大,并且存在多個明顯拐點,最明顯的是2019年5月,平均溫度達到最高的28.8℃,明顯高于洱源東湖的24.5℃和羅時江的20.4℃。又因螞蟻生長活動最佳的溫度為25~35℃之間,所以洱海月濕地采集到的螞蟻數量大于另外兩個濕地,優勢物種的出現對群落結構的穩定性產生較大的影響。而羅時江常年溫度低于另外兩個濕地,采集到的物種種類與物種數量都低于另外兩個濕地。由此推測:洱源東湖濕地的溫度更適中,全年溫度變化更加平緩,更適合昆蟲群落均衡發展。可見,溫度是影響昆蟲群落結構參數的主要因素。
4.6 昆蟲群落穩定性分析 本文采用式(5)、(6)對昆蟲群落的相對穩定性進行研究。分別對3個濕地每個月的穩定性指標進行計算并求取平均值,得到3個濕地的昆蟲群落相對穩定性數值,見表9。
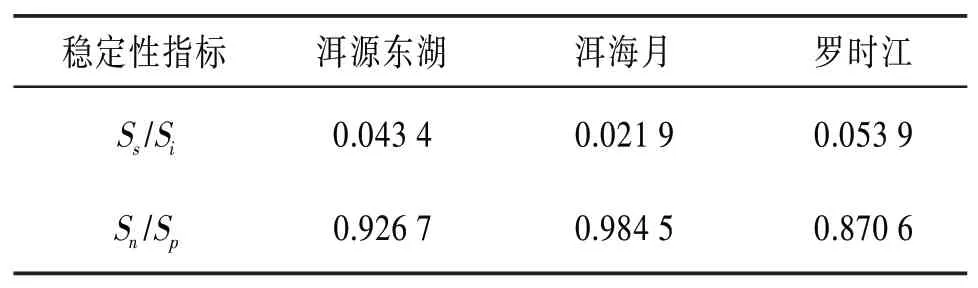
表9 昆蟲群落的相對穩定性數值表
由表9可知S s/Si的大小順序為羅時江>洱源東湖>洱海月,Sn/S p的大小順序為洱海月>洱源東湖>羅時江。其中洱源東湖濕地的相對穩定性指標數值都位于第二,且接近第一,而另外兩個濕地的穩定性指標數值都有偏小項。所以綜合分析可知,洱源東湖濕地的昆蟲群落網絡內部關系較為復雜,穩定性最高。另外兩個人為建設的濕地昆蟲群落穩定性雖然低于洱源東湖,但水平接近洱源東湖濕地。
5 結論
本文通過昆蟲群落結構及其特征參數研究,結果發現:對濕地昆蟲群落影響較大的除了四季變化帶來的溫度等自然生境變化,就是人類活動對其產生的干擾。3個洱海濕地的昆蟲群落結構具有相似性,物種較為豐富。其中,洱源東湖濕地的昆蟲群落結構最穩定,其余兩個濕地昆蟲群落也具有較強的自我調節修復能力。洱海濕地的昆蟲群落多樣性指數、豐富度指數以及均勻度指數越大,優勢集中性指數越小,反映出昆蟲群落物種豐富,結構穩定,所監測的棲息地生態環境越好。
同時,洱海月濕地昆蟲群落結構多樣性表明,通過人為建設改造后的濕地,從長遠來看并不一定會破壞濕地環境。相反,如果能夠維持昆蟲群落結構穩定性和自適應性,也能達到原生自然生境的生態標準。所以合理的建設規劃對于洱海濕地保護很有必要。如果人類活動過于頻繁,將導致極端優勢物種的產生,勢必將在一定程度上破壞昆蟲群落的穩定性。所以合理地劃分人類活動區域和原生濕地區域,對保護濕地生態環境具有重要意義。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
