基于CRISPR/Cas9技術的水稻OsARF12突變體的構建
2021-07-01 08:07:10杜想想趙亞帆趙晨云趙帥兵趙全志杜彥修孫紅正孫虎威
華北農學報 2021年3期
關鍵詞:水稻
杜想想,趙亞帆,趙晨云,趙帥兵,李 源,程 遠,趙全志,杜彥修,孫紅正,孫虎威,彭 廷
(河南糧食作物協同創新中心,河南省水稻生物學重點實驗室,河南農業大學,河南 鄭州 450046)
生長素作為最早發現的一類植物激素,參與調控植物多種生長發育過程,如根的發育[1]、頂端優勢、向性反應和形態建成[2]等。生長素調控通路中關鍵蛋白主要包括生長素/吲哚乙酸蛋白(Aux/IAAs)、SCF復合系統和生長素響應因子(ARFs)[3]。生長素響應因子是能與生長素應答元件(AuxRE)TGTCTC序列特異結合,調節生長素反應基因的一類轉錄因子,最先在模式植物擬南芥中被鑒定出來[4],該家族共有23個基因[5]。其中ARF1/ARF2功能的缺失通過增加擬南芥中Aux/IAA基因的轉錄進而調控葉片衰老和開花時間[6];ARF3/ARF4主要調控側生器官的結構發育[7],ARF3直接與細胞分裂素基因AtIPT5啟動子結合,負調控AtIPT5的表達,介導生長素與細胞分裂素相互作用,影響新生芽的再生[8];ARF5通過調控AMP1的表達影響胚和維管組織的形成[9];arf6、arf8突變體雄蕊發育遲緩,而arf6/arf8雙突變體則不能形成成熟的花,表明ARF6、ARF8共同參與花的形態建成[10],同時,ARF8還負調控果實的起始發育[11];ARF7/ARF19可直接激活下游基因LBD/ASLs的表達來影響側根形成[12];ARF10/ARF16則是在miR160下游調控根冠發育及根的向地性[13]。
單子葉植物水稻ARF家族中有25個成員[14]。OsARF1是水稻中第一個被發現的ARF基因[15],它與胚芽鞘的向性有關;之后的研究表明,OsARF1對營養生長和種子發育至關重要[16];OsARF4能與OsGSK41/OsGSK互作且被后者磷酸化,調控水稻籽粒大小及千粒質量[17];OsARF16參與細胞分裂素介導的水稻磷酸鹽轉運和信號傳遞通路,且敲除株系對外源細胞分裂素不響應[18]。OsARF17和OsARF19通過調節生長素和BR信號來控制水稻葉夾角大小[19-20]。然而,目前對水稻ARF家族的研究大多集中在籽粒發育和葉夾角等方面,ARFs其他成員對水稻農藝性狀的影響尚不明確。
CRISPR/Cas9是第3代基因編輯技術,相比其他編輯技術具有成本低、快速高效等優點,已經成為目前最主流的基因編輯系統[21]。近年來,大量研究人員通過CRISPR/Cas9技術對水稻基因編輯,研究水稻基因的功能,也取得了重要進展[22]。本研究利用CRISPR/Cas9系統對水稻生長素響應因子OsARF12第1個和第2個外顯子靶位點進行編輯,獨立轉化粳稻品種日本晴,通過對2個外顯子不同突變位點的KO-ARF12-1和KO-ARF12-2突變體進行測序、表達量鑒定、表型性狀調查,研究OsARF12對水稻農藝性狀的影響。
1 材料和方法
1.1 試驗材料
轉基因受體材料為:粳稻品種日本晴(Oryzasativaspp.japonicacv. Nipponbare)。CRISPR/Cas9載體為:pOs-gRNA、pH-Ubi-CAS9。試驗所需引物(表1)合成與測序分析均由上海生工生物工程股份有限公司完成。

表1 試驗所用引物Tab.1 Primers used in this test
1.2 試驗方法
1.2.1OsARF12靶位點設計和表達載體構建 根據CRISPR/Cas9原理,在RAP-DB網站(https://rapdb.dna.affrc.go.jp/)上獲取水稻OsARF12外顯子序列,分別選取第1個外顯子PAM序列(CGG)前20 bp(5′-GAACGATGCGTACCTTCCCG-3′)及第2個外顯子PAM序列(GGG)前20 bp(5′-TTCACACCCGGATATCGGTT-3′)為靶位點(圖1)。分別在2個靶位點5′端前加上BsaⅠ限制性內切酶的黏性末端接頭GGCG,即為ARF12Crispr1_F、ARF12Crispr2_F(表1);將選取的2個靶序列分別反向互補并在其5′端前加上BsaⅠ限制性內切酶的黏性末端接頭AAAC,即為ARF12Crispr1_R、ARF12Crispr2_R(表1)。將2對加過接頭的序列送往上海生工生物工程股份有限公司合成后,經磷酸化修飾和退火形成雙鏈,用T4DNA連接酶與經過限制性內切酶BsaⅠ酶切過后的中間載體sgRNA連接,轉化大腸桿菌感受態DH5α,隨后進行菌落PCR檢測與測序,并用陽性質粒與Cas9終載體進行LR重組,轉化DH5α,經菌落PCR和測序檢測,將陽性質粒轉化農桿菌EHA105。利用農桿菌介導法導入粳稻品種日本晴,獲得轉基因植株。
1.2.2 植株DNA和總RNA的提取 在野生型和各轉基因植株抽穗前取葉片樣品,并立即置于液氮中,-80 ℃冰箱保存。采用CTAB法提取葉片DNA;TRIzol法提取總RNA。
1.2.3 反轉錄 使用天根生化科技有限公司FastKing RT Kit(KR118-02)反轉錄試劑盒將1 μg總RNA反轉錄成cDNA。
1.2.4OsARF12表達量分析 將cDNA稀釋10倍,使用天根生化科技有限公司定量檢測試劑盒SYBR Green(FP209),Actin基因作為內參,采用Bio-Rad CFX96熒光定量PCR儀進行實時熒光定量PCR分析。相對表達量采用2-ΔΔCt法計算。
1.2.5 數據分析 采用Microsoft Excel 2016對試驗數據進行整理和作圖,利用SPSS 24進行差異顯著性分析。
2 結果與分析
2.1 KO-ARF12-1/-2 T0突變體鑒定
將攜帶CRISPR/Cas9-ARF12-1和CRISPR/Cas9-ARF12-2質粒的農桿菌侵染水稻品種日本晴,得到KO-ARF12-1 T0再生苗5株,KO-ARF12-2 T0再生苗15株。利用CTAB法提取各單株葉片的DNA,并根據OsARF12的序列分別設計2個不同靶位點引物:CrisprARF12-1_F、CrisprARF12-1_R和CrisprARF12-2_F、CrisprARF12-2_R(表1),用對應的引物對轉基因植株的DNA進行擴增(圖2),并將擴增產物送往公司測序。通過對T0測序結果比對發現:KO-ARF12-1-1為1條鏈缺失1 nt、互補鏈缺失7 nt的雙等位突變體;KO-ARF12-1-2和KO-ARF12-1-3為1條鏈缺失4 nt、互補鏈缺失7 nt的雙等位突變體,KO-ARF12-1-4和KO-ARF12-1-5 2條鏈均未突變(表2)。
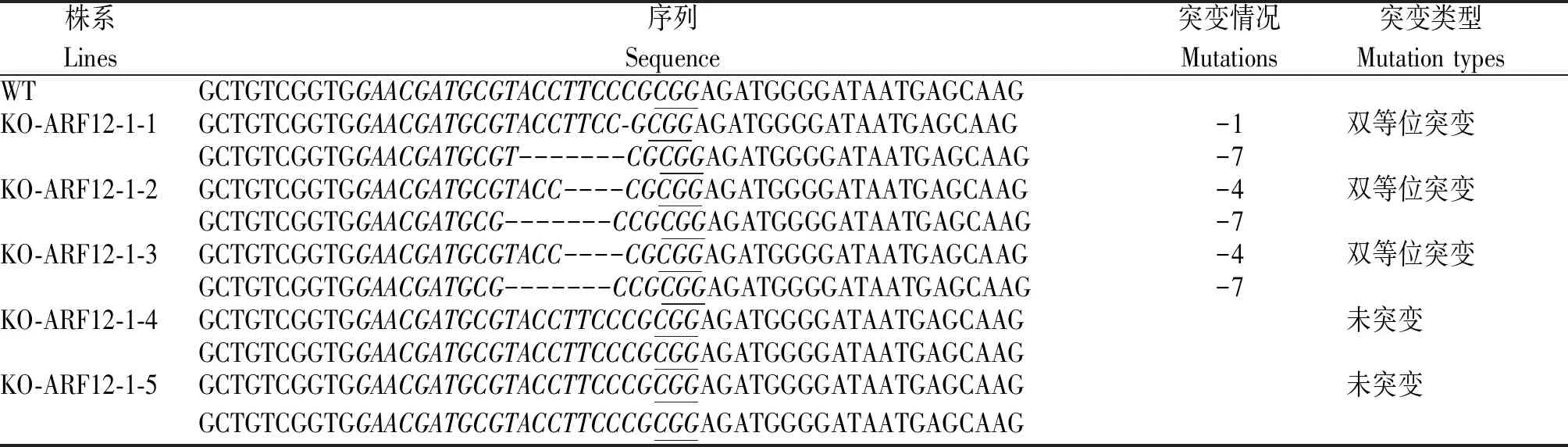
表2 KO-ARF12-1 T0轉基因植株突變類型鑒定Tab.2 Identification of mutation types of KO-ARF12-1 T0 transgenic plants
KO-ARF12-2各轉基因植株共有11種突變類型:KO-ARF12-2-12和KO-ARF12-2-15為2條鏈分別缺失了6 nt與5 nt的純合突變體;KO-ARF12-2-5為1條鏈缺失2 nt,KO-ARF12-2-8為1條鏈插入1 nt,互補鏈均沒有突變的雜合突變體;而其余8株均為雙等位突變:KO-ARF12-2-1為1條鏈插入1 nt,互補鏈缺失4 nt;KO-ARF12-2-3為1條鏈缺失1 nt,互補鏈缺失6 nt;KO-ARF12-2-6、KO-ARF12-2-9和KO-ARF12-2-11為1條鏈插入1 nt,互補鏈缺失1 nt;KO-ARF12-2-7為1條鏈插入1 nt,互補鏈缺失7 nt;KO-ARF12-2-13為1條鏈缺失4 nt,互補鏈缺失5 nt;KO-ARF12-2-14為1條鏈缺失1 nt,互補鏈缺失5 nt;KO-ARF12-2-2、KO-ARF12-2-4和KO-ARF12-2-10 2條鏈均未突變(表3)。
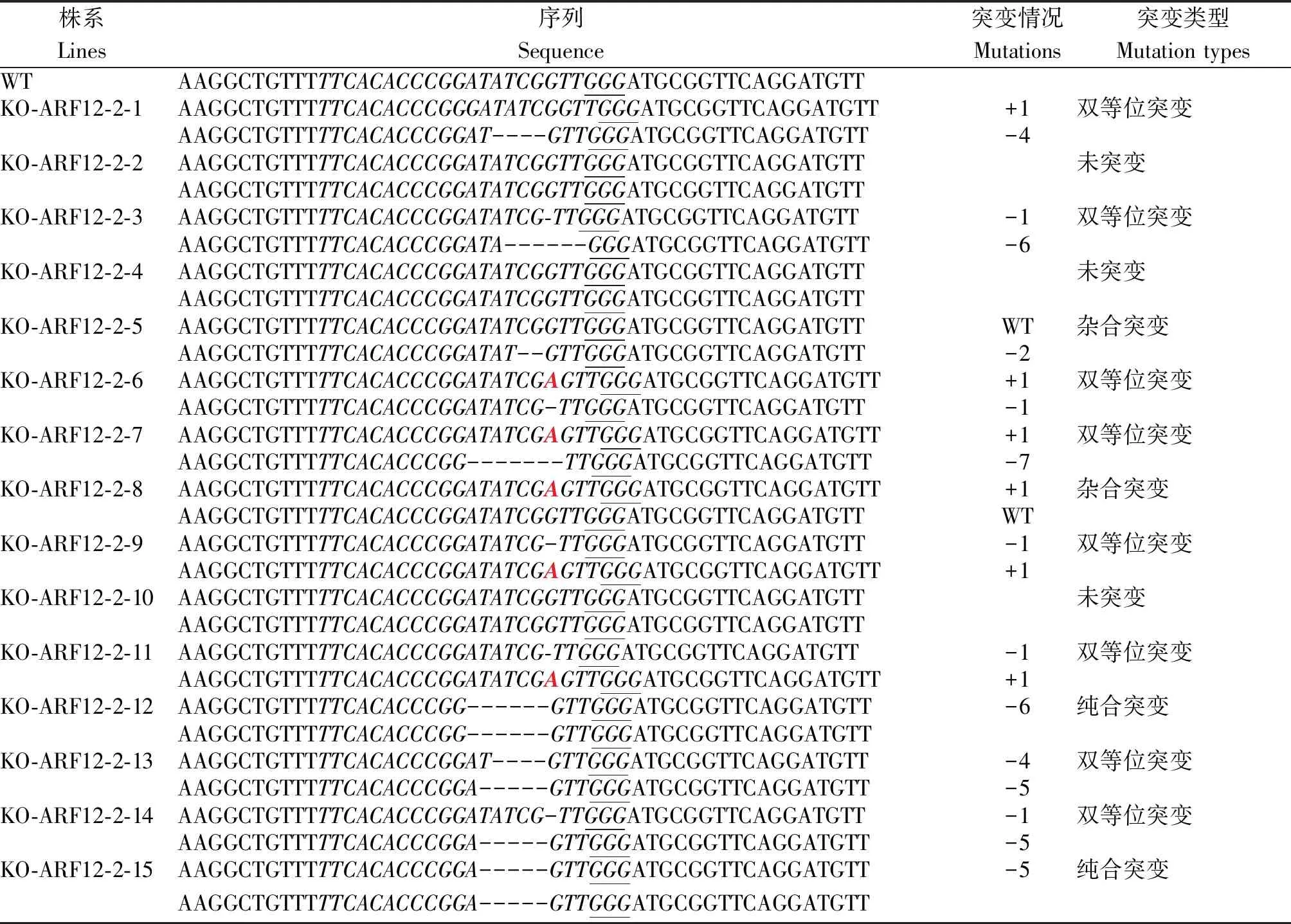
表3 KO-ARF12-2 T0轉基因植株突變類型鑒定Tab.3 Identification of mutation types of KO-ARF12-2 T0 transgenic plants
2.2 KO-ARF12-1/-2 T1突變體鑒定
采用CTAB法提取OsARF12在T0發生突變的轉基因株系葉片DNA,并用CrisprARF12-1_F、CrisprARF12-1_R和CrisprARF12-2_F、CrisprARF12-2_R對相應的轉基因株系DNA進行擴增(圖3,4),將產物送往公司測序。測序結果進行比對發現,KO-ARF12-1各突變轉基因株系T120個單株中,有3種純合突變、2種雙等位突變,共5種突變類型,分別為:1條鏈缺失1 nt、互補鏈缺失7nt的雙等位突變體有4株;缺失1 nt的純合突變體有1株;缺失7 nt的純合突變體有3株;缺失4 nt的純合突變體有7株;1條鏈缺失4 nt、互補鏈缺失7 nt的雙等位突變體有5株(表4)。

表4 KO-ARF12-1 T1轉基因植株突變類型鑒定Tab.4 Analysis and identification of mutation types of KO-ARF12-1 T1 transgenic plants
KO-ARF12-2各突變轉基因株系T160個單株中,有6種純合突變、1種雜合突變和4種雙等位突變,共11種突變類型,分別為:缺失1,6,2,7,4 nt和插入1 nt的純合突變體,分別為12,1,6,3,2,10株;1條鏈插入1 nt、互補鏈未突變的雜合突變體3株;1條鏈插入1 nt、互補鏈缺失4 nt,1條鏈插入1 nt、互補鏈缺失1 nt,1條鏈缺失4 nt、互補鏈缺失5 nt和1條鏈缺失1 nt、互補鏈缺失5 nt的雙等位突變體分別為6,9,4,4株(表5)。
2.3 KO-ARF12-1/-2 T2突變體表達量分析
以T22種不同靶位點的6種不同基因型純合突變體KO-ARF12-1-1-3(缺失1 nt)、KO-ARF12-1-2-1(缺失4 nt)、KO-ARF12-1-2-6(缺失7 nt)及KO-ARF12-2-5-2(缺失2 nt)、KO-ARF12-2-11-4(插入1 nt)、KO-ARF12-2-15-5(缺失5 nt)(圖5-A)為研究材料,選取大田正常生長條件下葉片樣品提取總RNA,反轉錄并定量分析發現,除KO-ARF12-2-11-4,其他各突變體株系中OsARF12表達量與野生型相比均顯著下降(P<0.05)(圖5-B)。

表5 KO-ARF12-2 T1轉基因植株突變類型鑒定Tab.5 Analysis and identification of mutation types of KO-ARF12-2 T1 transgenic plants

表5(續)
2.4 OsARF12對水稻株高的影響
為研究OsARF12對水稻農藝性狀的影響,于灌漿期將突變體株系和野生型株系進行比較,結果表明,KO-ARF12-1/-2的株高均顯著降低(圖6-A、C),各突變體株系株高與野生型相比分別減少了16.59%,16.63%,16.06%和14.94%,13.15%,18.39%(圖6-C)。進一步對水稻各莖節間長度進行統計分析發現,與野生型相比,KO-ARF12-1/-2顯著降低了第3和第4莖節間長度,其中第4莖節間降低幅度最大(圖6-B、D),降低的幅度為23.46%(P<0.05),其次為第3莖節間,降低的幅度為21.91%(P<0.05),而第1和第2莖節間長度與對照無顯著差異(P>0.05)。
3 結論與討論
現代農業發展中,對突變體進行基因功能研究已經成為一種重要的研究手段。2013年CRISPR/Cas9技術成功應用于定點突變水稻基因[23],2014年CRISPR/Cas9技術被證實可高效編輯水稻特異基因,并且基因突變能夠穩定遺傳[22],極大地促進了水稻突變體庫的建立。本研究利用CRISPR/Cas9技術分別對OsARF12的第1、第2個外顯子靶序列進行定點編輯,獲得了OsARF12多種類型的突變體。其中,KO-ARF12-1 T03種突變類型,T15種突變類型;KO-ARF12-2 T011種突變類型,T111種突變類型,為研究OsARF12對水稻農藝性狀影響奠定基礎。
通過對KO-ARF12-1/-2 2種突變體材料中OsARF12的表達量進行鑒定發現,CRISPR/Cas9基因編輯技術顯著降低了OsARF12的表達量。且與對照相比,KO-ARF12-1/-2株高顯著降低(P<0.05),表明OsARF12在一定程度上正向調節水稻的株高性狀。已有研究借助T-DNA插入或者是Tos17插入的arf12突變體研究發現,OsARF12可能介導根形態和磷誘導的生長素信號反應;Qi等[24]發現,OsARF12改變鐵調節蛋白(OsMIR)和鐵調節轉運蛋白1(OsIRT1)的豐度,導致鐵含量的改變。本研究通過CRISPR/Cas9基因編輯技術創制了OsARF12突變體,研究了其在調節水稻株高方面的重要作用,豐富了水稻生長素響應因子OsARF12的生物學功能。
水稻的株高受眾多激素以及它們之間的相互作用調控,OsMADS57功能的缺失降低了GA的活性,導致株高降低[25];OsMED14_1與轉錄因子YABBY5、TDR和MADS29互作,參與調節生長素的動態平衡,進而調控水稻株高[26];miR1848靶向OsCYP51G3調節水稻中植物甾醇和BR的生物合成,影響水稻株高[27]。本研究通過CRISPR/Cas9技術對生長素響應因子OsARF12進行編輯,可能通過影響其生長素調控路徑,進而導致株高降低。水稻株高由各莖節間長度組成,水稻抗倒伏能力與近基部莖節間長度呈負相關關系[28]。本研究表明,各突變類型的突變體與野生型相比,第1和第2莖節間長度無顯著差異,第3和第4莖節間長度顯著(P<0.05)降低,且第4莖節間降幅最大,表明突變體植株越靠近基部,其莖節間長度縮短幅度越大,這對提高水稻的抗倒伏能力有重要作用,但KO-ARF12突變體中不同節間長度改變的內在機制需要進一步研究。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
