脫落酸對水稻種子萌發期耐高溫脅迫的誘抗效應
2021-07-01 08:07:06楊雲雲陳啟洲楊洪濤蘇佩佩劉曉龍
華北農學報 2021年3期
關鍵詞:水稻
楊雲雲,陳 鑫,陳啟洲,盧 芳,徐 晨,楊洪濤,蘇佩佩,劉曉龍
(1.宜春學院 生命科學與資源環境學院,江西 宜春 336000;2.吉林省農業科學院 農業資源與環境研究所,吉林 長春 130033)
長江中下游地區是我國重要的水稻主產區,近年來,由于氣候變暖導致的夏季極端高溫天氣時有發生,最高溫度超過35 ℃的天氣持續10~15 d。高溫脅迫已對當地的水稻生產造成嚴重影響,并已成為制約當地水稻種植業發展的主要限制因素之一。高溫脅迫會對不同生育時期的水稻生長造成影響,在種子萌發期遭遇超過35 ℃的高溫脅迫會導致種子發芽遲緩、種子喪失活力、發芽率下降[1]。水稻在幼苗階段遭受高溫脅迫不僅會出現葉片卷曲,含水量降低,葉片顏色變淺、變白、變短和畸形等癥狀,還會致使植株生長發育緩慢、抑制根系生長[2-3],光合作用受阻,凈光合速率和氣孔導度降低,光合作用原初反應受到抑制[4-5]。水稻孕穗期遭遇高溫脅迫會導致穎花退化、花粉敗育、花粉活力下降,進而導致秕粒增加[6-7];在抽穗期和灌漿期遭受高溫脅迫會阻礙水稻的開花授粉,穗中秕粒增多、結實率下降,造成千粒質量降低,進而導致水稻減產[8-11]。研究表明,水稻幼苗在堿脅迫環境下,根系活性氧大量積累,破壞了抗氧化防御系統,進而損傷根系細胞,導致幼苗萎蔫和死亡[12]。高溫脅迫與堿脅迫類似,其抑制水稻生長發育的一個重要因素就是引起ROS過量積累,破壞ROS產生和清除的平衡體系,導致膜質過氧化作用加劇,進而損壞細胞結構[3,13-14]。高溫脅迫抑制水稻種子萌發,但其導致發芽率下降的主要原因尚不明確,本研究擬以ROS積累為線索探究高溫脅迫抑制水稻種子萌發的機制。
脫落酸(ABA)是植物體內的重要激素,常作為脅迫激素參與植物對多種逆境脅迫的響應,并發揮重要作用[15]。產生誘抗效應是ABA提高植物抗逆性的一個重要機制[16]。ABA對水稻耐鹽堿脅迫產生誘抗效應,能夠提高水稻對堿脅迫的抗性和蘇打鹽堿水田中的產量[17-18]。ABA在提高植物耐高溫脅迫中同樣具有促進作用,高溫處理可降低植物體內IAA、GA、自由脯氨酸及可溶性蛋白質的含量,增加脫落酸含量[19]。抽穗期噴施S-誘抗素或ABA溶液能夠提高水稻的結實率和千粒質量,稻米的碾磨品質和蒸煮品質也得到改良[20]。研究表明,外源ABA能夠通過增加蔗糖的轉運和加速蔗糖代謝來保持碳平衡和能量平衡,進而阻止花粉敗育[21]。以上研究結果為ABA提高水稻耐高溫特性的效果提供了堅實的科學依據,但目前對于其內在的生理機制及分子機理解析尚不明晰,尤其在ROS相關通路中的報道并不多見。研究表明,ABA預處理能夠提高堿脅迫下水稻幼苗的下游抗氧化防御能力,抑制ROS過量積累;且ABA緩解了外源二氯百草枯(Paraquat)對水稻幼苗引發的氧化脅迫,這是ABA誘導水稻耐堿脅迫的主要途徑之一[22]。基于以上研究結果,本研究以高溫為主要非生物脅迫因子,探究ABA對水稻耐高溫脅迫的誘抗效應,以ABA浸種的方式在水稻種子萌發期進行研究,并以ROS積累為出發點初步解析高溫環境下ABA對水稻種子萌發的影響機理。
1 材料和方法
1.1 供試材料
以江西省主推常規水稻品種黃華占和日本晴為試驗材料。
1.2 試驗設計
1.2.1 ABA溶液的配制 脫落酸(ABA:Sigma,Inc.,St,Louis,MO,USA)試劑先溶于少量的無水乙醇中,然后用去離子水定容至一定的濃度。本試驗用外源ABA浸種的方式探究ABA對水稻種子萌發的誘抗效應,參照前人研究結果,選用10 μmol/L作為試驗用ABA濃度[17-18,22]。
1.2.2 試驗設計 萌發試驗在培養皿中進行,設置2個處理,即非ABA處理組(-ABA)和ABA處理組(+ABA),每個處理5次重復。每個重復選取飽滿一致的50粒種子,ABA處理組用10 μmol/L的ABA溶液于黑暗條件下浸種24 h,非ABA處理組用去離子水于黑暗條件下浸種24 h。
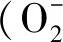
1.3 各項指標的測定方法
1.3.1 相對電導率和丙二醛含量的測定 相對電導率(Relative conductivity,RC)常用來評價細胞膜的損傷程度。用煮沸前和煮沸后的電導率(R1和R2分別表示煮沸前后溶液的電導率)來計算,公式為:RC(%)=R1/R2×100%。丙二醛(Malondialdehyde,MDA)含量采用硫代巴比妥酸顯色法測定,采用公式6.45×(A532-A600)-0.56×A450計算各樣品的MDA濃度,再根據樣品質量計算MDA含量。
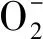
過氧化氫含量的測定:植物中的H2O2與硫酸鈦(或氯化鈦)反應生成黃色的過氧化物-鈦復合物沉淀,在H2SO4中進行溶解,在415 nm處有特征吸收峰,用分光光度計比色可測定H2O2含量[25]。
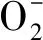
1.3.3 萌發相關基因的表達分析 利用TRIzol法提取水稻幼芽總RNA,并測定RNA濃度,用M-MLV反轉錄酶(TaKaRaBioInc.,Otsu,Japan)進行反轉錄,形成cDNA。在實時熒光定量PCR儀(Eco TM 48,Illumina,Saffron Walden,UK)上進行qRT-PCR反應(Quantitative real-time PCR),總反應體系為20 μL,包括:10 μL 2×SYBRPremixExTaqTM(TaKaRa Bio),1.6 μL cDNA,0.8 μL引物和7.6 μL ddH2O。qRT-PCR反應程序為:95 ℃ 5 min;95 ℃ 5 s,60 ℃ 30 s,30個循環;95 ℃ 1 min,55 ℃ 30 s,95 ℃ 30 s。確定cDNA模板和引物沒有基因組DNA污染后,以穩定表達的水稻看家基因β-actin作為內參,進行基因表達量的測定。每個處理設置3次生物學重復,每個模板2次技術重復,利用2-ΔΔCT的方法計算基因的相對表達量[26]。
ROS清除相關基因包括:OsCATB、OsAPX6、OsFe-SOD和OsCu/Zn-SOD;細胞死亡相關基因包括:OsBI1和OsKOD1;ABA應答基因有Salt和OsWsi18[12,22],用于qRT-PCR的基因及相關引物(5′-3′)如下:
OsACT1(內參,Os03g0718100)F:TTCCAGCCTT
CCTTCATA,R:AACGATGTTGCCATATAGAT;
OsBI1(Os02g0125300)F:CTACATCAAGCACGC
ACTC,R:ACCTCTTCTTCCTCTTCTTCTC;
OsKOD1(Os04g0507950)F:TCAAGCCATTCATC
TTCCAT,R:ATCAGCAACCTCGTCAAG;
Salt(Os01t0348900)F:CGAAATAATGTTCCATG
GTGTT,R:TGTACTACGGATCGGTGCAA;
OsWsi18(Os01g0705200)F:TGTGACTCGATCCA
GCGTAG,R:GTTCCTGCTGAGAAGCCATC;
OsCATB(Os06g0727200)F:GCTGGTGAGAGATACC
GGTCA,R:TCAACCCACCGCTGGAGA;
OsAPX6(Os12g0178100)F:CCCCAAGATCCCCA
TGATCTA,R:CCTCTGGCGGGCATTG;
OsFe-SOD(Os06g0143000)F:CGACGCCGAGGAAT
TTCTAG,R:AGGTGGTGTAAGTGTCTCTCATGC;
OsCu/Zn-SOD(Os06g0130900)F:TGTGACGGGA
CTTACTCCTGG,R:CACCCATTCGTAGTATCGCCA。
1.4 數據處理與分析
采用Excel 2013軟件進行數據統計,數據繪圖應用Origin 9.1繪圖軟件。利用SPSS 21.0(IBM Corp.,Armonk,NY)軟件,基于單因素方差分析(ANOVA)和Duncan方法進行數據的顯著性差異分析,顯著性差異水平為P<0.05。
2 結果與分析
2.1 ABA浸種預處理對高溫脅迫下水稻種子萌發的影響
不同處理下,種子萌發試驗結果如圖1所示。非高溫處理下,ABA浸種與否對種子萌發無明顯影響。高溫脅迫抑制了日本晴和黃華占的種子萌發,導致芽長和根長縮短。而ABA浸種能夠促進高溫脅迫下水稻種子的萌發和幼芽的生長。
發芽率統計試驗結果表明(圖2),ABA浸種在高溫脅迫初期促進了水稻種子的萌發。非ABA浸種條件下,與CK-ABA相比,高溫處理使日本晴在第2天和第3天的發芽率分別提高了11.60,7.60百分點,使黃華占在第2天,第3天的發芽率分別提高了14.00,4.00百分點。ABA浸種條件下,與CK+ABA相比,高溫處理使日本晴在第2天,第3天的發芽率分別提高了7.60,9.20百分點,使黃華占在第2天,第3天的發芽率分別提高了14.40,6.80百分點。隨著萌發時間的延長,不管ABA浸種與否,高溫脅迫均抑制了水稻種子的萌發。在萌發的第7天,高溫脅迫使ABA浸種和非ABA浸種的日本晴發芽率分別下降了12.00,22.80百分點,黃華占分別下降了10.40,20.00百分點。
對照條件下,ABA浸種的種子發芽率略低于非ABA浸種。而高溫脅迫下,ABA浸種對水稻種子的萌發起到了一定的促進作用,且脅迫時間越長,促進作用越明顯。從萌發的第5 天開始,高溫脅迫條件下,ABA浸種提高了2個水稻品種的發芽率;在萌發的第6天,ABA浸種使日本晴和黃華占的發芽率分別提高了4.40,6.80百分點;第7天,ABA浸種使日本晴和黃華占的發芽率分別提高7.60,8.00百分點。
ABA浸種促進了水稻芽和根的生長。試驗結果表明(圖3),對照條件下,ABA浸種對水稻幼芽和根的生長略有促進作用,ABA浸種顯著提高了黃華占的主根長(P<0.05)。高溫脅迫下,ABA浸種顯著提高了水稻的芽長和主根長(P<0.05),黃華占的芽長增加更明顯。高溫脅迫下,ABA浸種使日本晴和黃華占的芽長分別提高了21.96%,38.92%;主根長分別提高了48.00%,63.82%。
2.2 ABA浸種對高溫脅迫下水稻幼芽ROS積累的影響
研究表明,外源ABA預處理顯著上調了多個ROS清除基因的表達,提升抗氧化酶活性,進而抑制ROS過量積累來提高水稻幼苗的耐堿性。在這些ROS清除基因中上調幅度較大的有OsCATB、OsAPX6、OsFe-SOD、OsCu/Zn-SOD,分別為過氧化氫酶(CAT)、抗壞血酸過氧化物酶(APX)和超氧化物歧化酶(SOD)的主要調控基因[22]。本試驗繼續分析高溫脅迫下ABA浸種對以上4個ROS清除基因表達的影響。結果表明(圖5),高溫脅迫下,ABA浸種顯著上調了4個ROS清除基因的表達(P<0.05),且OsFe-SOD和OsCu/Zn-SOD的上調幅度更大,暗示著ABA浸種提高了抗氧化酶CAT、APX和SOD的活性以清除過多的ROS。
2.3 ABA浸種對高溫脅迫下水稻幼芽細胞死亡的影響
研究表明,逆境脅迫下ROS的過量積累是導致水稻細胞損傷和幼苗萎蔫的主要原因[12],本試驗進一步對高溫脅迫下水稻幼芽的細胞損傷狀況進行測定。由圖6可知,高溫脅迫條件下,ABA浸種處理使幼芽中細胞死亡抑制基因OsBI1的表達量顯著上調(P<0.05),而細胞死亡基因OsKOD1表達下調,進而降低了高溫脅迫導致的細胞過量死亡。高溫脅迫下,與非ABA浸種處理相比,ABA浸種使2個品種OsBI1的上調幅度分別為182.0%,156.0%,而細胞死亡基因OsKOD1則分別下調了33.5%,51.3%。
2.4 高溫脅迫下水稻種子萌發與幼芽生理指標的相關性分析
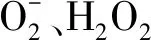
2.5 高溫脅迫下水稻幼芽ABA應答基因表達量的變化
進一步分析了2個ABA應答基因的表達變化,結果如圖7所示。高溫脅迫下,ABA浸種處理下水稻幼芽內ABA應答基因顯著(P<0.05)上調,說明ABA浸種激活了水稻內的ABA信號通路,使之在抵抗高溫脅迫中發揮作用。高溫脅迫下,與非ABA浸種處理相比,ABA浸種使日本晴和黃華占幼芽內Salt基因相對表達量分別提高了4.84,5.46倍(圖7-A),OsWsi18分別提高了5.87,7.06倍(圖7-B)。
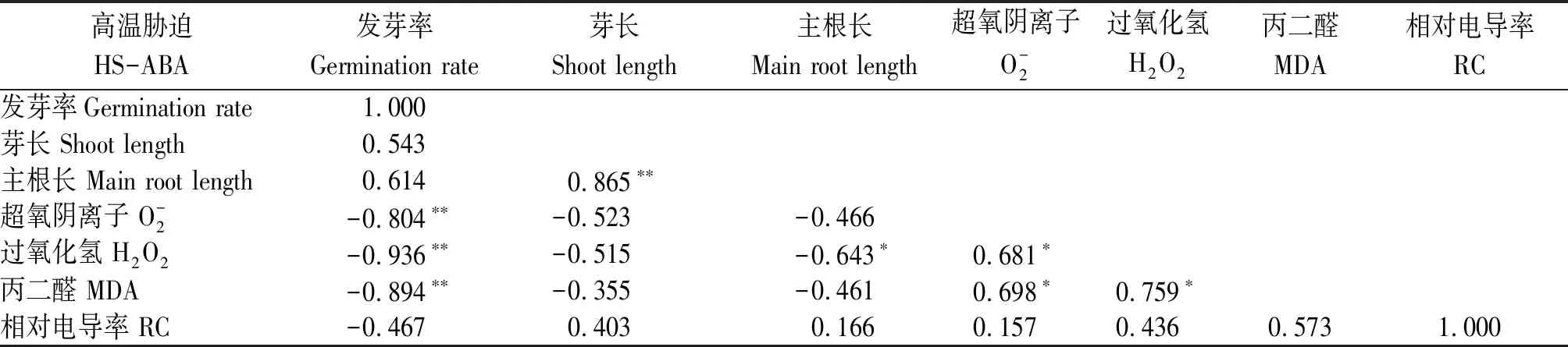
表1 高溫脅迫下非ABA浸種處理下水稻種子萌發進程中各指標的相關性分析Tab.1 Correlation analysis of various indexes in rice seed germination process of non-ABA soaking treatment under high temperature stress condition
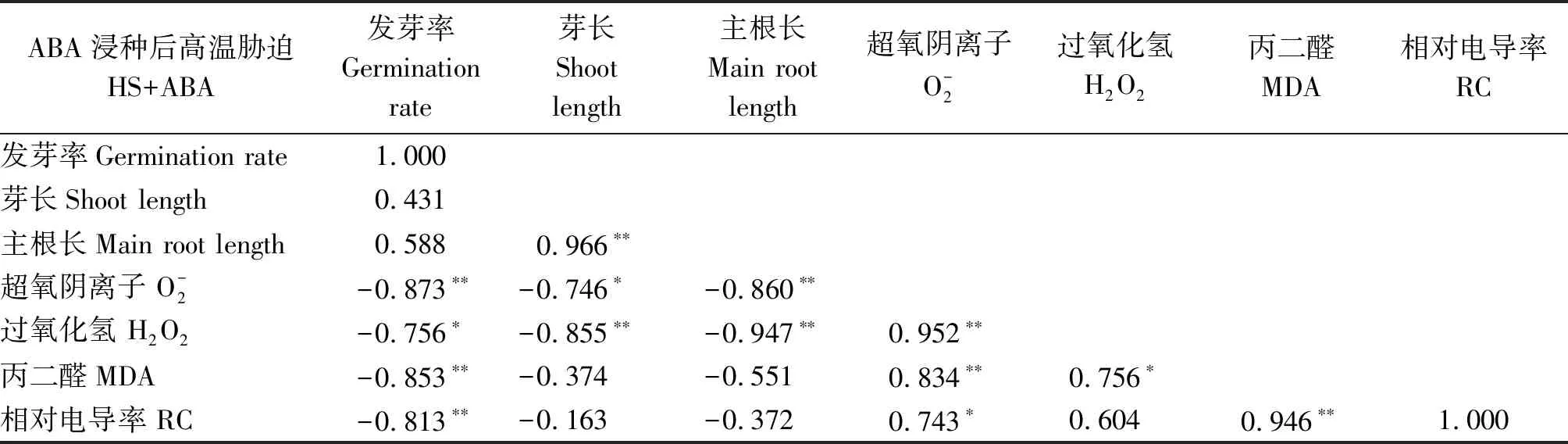
表2 高溫脅迫下ABA浸種處理下水稻種子萌發進程中各指標的相關性分析Tab.2 Correlation analysis of various indexes in rice seed germination process of ABA soaking treatment under high temperature stress condition
3 討論與結論
良好的種子萌發率是植物適應環境的先決條件。一般來說,水稻在種子萌發期對高溫脅迫具有較強的抵抗力,這是因為誘導了種子中小熱激蛋白的表達,進而能夠幫助種子抵御高溫;但超過35 ℃時水稻種子的萌發受阻,種子喪失活力,發芽率降低[27]。本研究結果表明,在萌發初期(前3 d),水稻種子在30 ℃的環境和40 ℃的高溫環境下均能夠正常萌發,且高溫處理對種子萌發具有促進作用,發芽率略高于對照處理,這可能是由于種子經24 h浸泡后,含水量增加,能夠在短期內抵抗高溫的緣故,這與前人研究結果基本一致[28]。在萌發初期,對照條件和高溫脅迫下,ABA浸種對種子萌發有抑制作用,2個品種的發芽率略低于非ABA浸種處理。隨著萌發進程的推進,不管ABA浸種與否,與對照相比,高溫脅迫均抑制了水稻種子的萌發;在萌發的第7 天,2個品種的發芽率在高溫處理下較對照均有不同程度的下降,且芽長和主根長顯著(P<0.05)低于對照處理,這與多數研究結果基本一致[29-31]。不管ABA浸種與否,在對照條件下,品種黃華占的芽長和主根長高于日本晴;且在高溫脅迫下的芽長和主根長的下降幅度以及幼芽內ROS積累和質膜損傷程度均低于日本晴,說明黃華占在種子萌發期對高溫的抗性略高于日本晴。
ABA不僅參與調控植物的多種生長發育進程,且在植物應對鹽、堿、低溫和高溫等非生物脅迫和多種生物脅迫中發揮重要作用[15,32]。ABA調控植物抗逆性的一種重要機制就是產生誘抗效應,通過浸種和浸根等多種方式賦予植物抵抗逆境的潛在能力[18,22-33]。研究表明,水稻種子或幼苗經ABA預處理后能夠提高鹽脅迫下幼苗的生長發育和最終產量[34-35];水稻幼苗經外源ABA浸根預處理后,能夠提高抗氧化清除能力,抑制ROS的過量積累,進而提高對堿脅迫的抗性和蘇打鹽堿水田中的產量[17-18,22]。在高溫脅迫下,外源ABA能夠通過多種途徑緩解高溫脅迫對水稻的傷害,包括調控能量平衡、ROS積累和糖代謝等途徑,促進水稻生長發育和產量形成[2,21,36]。本試驗結果表明,種子萌發初期,在對照和高溫處理下,外源ABA浸種較非ABA浸種在一定程度上抑制了種子的萌發。在長期高溫脅迫下,外源ABA浸種加快了水稻種子的萌發速度,促進了幼芽和幼根的生長,緩解了長期極端高溫脅迫對水稻種子幼芽生長的抑制作用。對其內在機制進一步研究發現,高溫脅迫下,ABA浸種顯著(P<0.05)上調了ABA應答基因Salt和OsWsi18的表達量,說明ABA信號途徑被激活,進一步提升了高溫脅迫下水稻幼芽抗氧化清除基因的轉錄表達,降低ROS的含量,進而緩解了水稻幼芽的質膜損傷和細胞過量死亡,促進幼芽的生長。
綜上所述,本研究探究了高溫脅迫下ABA浸種對水稻種子萌發的影響,進一步驗證了外源ABA對水稻抗逆性的誘抗效應。水稻種子在萌發初期,高溫處理能夠短暫的促進種子萌發;此后,高溫處理導致幼芽內ROS過量積累,并進一步導致細胞損傷,進而抑制了幼芽的生長。種子在萌發前利用外源ABA浸種能夠起到提高抗氧化清除能力的作用,降低由極端高溫脅迫而引起的ROS過量積累,從而緩解細胞損傷,促進高溫脅迫下水稻幼芽及幼根的生長。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
