超高壓輔助脫殼對蝦夷扇貝肌原纖維蛋白生化特性及結構的影響
2021-07-01 09:08:48陳啟航文麗華陳小娥方旭波凌建剛宣曉婷
食品科學 2021年11期
陳啟航,文麗華,陳小娥,,方旭波,凌建剛,宣曉婷
(1.浙江海洋大學食品與藥學學院,浙江 舟山 316022;2.寧波市農業(yè)科學研究院農產品加工研究所,浙江 寧波 315000)
蝦夷扇貝(Patinopecten yessoensis)味道鮮美、營養(yǎng)豐富,其貝肉是食用的主要部位[1-2]。傳統的扇貝加工工藝主要采用手工脫殼和加熱脫殼,其中手工脫殼存在脫殼效率低、貝肉完整性差的缺點,而加熱脫殼易導致蛋白變性[3-4]。超高壓技術高效方便、能耗低,不僅脫殼率較高,且不會影響產品的品質,是當前食品加工領域研究的熱點[5]。目前國內外學者對超高壓技術在扇貝中應用的研究,主要集中在脫殼工藝[6-8]、保鮮包裝[9]、質構[10]等方面。鞏雪等[8-9]對扇貝的得肉率、持水率、總揮發(fā)性鹽基氮含量及菌落總數等指標進行研究,發(fā)現當壓強300 MPa、保壓時間180 s時,扇貝的脫殼率和得肉率較高,同時還發(fā)現通過超高壓處理并存放8 d后,扇貝的保鮮指標與未處理組相比,保鮮效果達到預期。孫少斌[10]通過比較超高壓處理與熱處理對扇貝的影響,發(fā)現扇貝經過超高壓處理后口感方面比熱處理組鮮嫩。
肌原纖維蛋白屬于鹽溶性蛋白,是貝肉的重要組成部分,其生化特性和結構的變化會對扇貝的食用品質以及商品價值產生影響[11-13]。超高壓在提高脫殼效果的同時也能作用于蛋白等生物大分子,使其肌原纖維蛋白的生化特性發(fā)生一定程度變化,導致蛋白變性。葉韜等[14]利用超高壓技術處理小龍蝦蝦仁,結果發(fā)現超高壓處理后其肌原纖維蛋白Ca2+-ATPase活性下降,表面疏水性上升。周果等[15]采用超高壓技術處理梭子蟹蟹肉,結果發(fā)現經過超高壓處理,蟹肉的肌原纖維蛋白總巰基含量呈現下降趨勢。因此,采用超高壓技術處理蝦夷扇貝貝肉的同時,除了研究超高壓工藝對扇貝脫殼效果的影響外,關注超高壓對其肌原纖維蛋白生化特性和結構的影響也十分重要。
本實驗以蝦夷扇貝為研究對象,通過超高壓處理,研究不同壓力對扇貝脫殼效果的影響,依據肌原纖維蛋白的含量、總巰基含量、羰基含量、表面疏水性、Ca2+-ATPase活力等指標,分析超高壓對貝肉肌原纖維蛋白生化特性產生的影響,并利用圓二色光譜探討肌原纖維蛋白二級結構的變化,完善超高壓技術在貝類脫殼中的應用研究,為扇貝加工和相關產品的進一步開發(fā)提供理論參考。
1 材料與方法
1.1 材料與試劑
鮮活蝦夷扇貝購于寧波路林水產品交易市場;牛血清白蛋白 北京Solarbio公司;Ca2+-ATPase活力測定試劑盒 南京建成生物工程研究所;Tris-馬來酸、5,5'-二硫代雙(2-硝基苯甲酸)、2,4-二硝基苯肼 上海Aladdin公司;其他試劑均為國產分析純。
1.2 儀器與設備
CQC2L-600型全液相超高壓設備 北京速原中天股份有限公司;MS105DU電子分析天平、Delta320 pH計瑞士Mettler Toledo儀器有限公司;H1850R型臺式高速冷凍離心機 湖南湘儀離心機儀器有限公司;ST-752s紫外-可見光分光光度儀 上海光譜儀器有限公司;MOS-450圓二色光譜儀 法國Biologic公司。
1.3 方法
1.3.1 超高壓處理樣品
將清洗干凈的蝦夷扇貝分裝在聚乙烯袋中,分別在150、200、250、300、350 MPa下處理3 min,溫度為20~25 ℃。另以手工脫殼(直接用手術刀手工脫殼)和蒸煮脫殼(鮮活扇貝100 ℃水蒸煮3~5 min)為對照組。將樣品標記后,置于4 ℃下用于后續(xù)各項指標的測定。
1.3.2 脫殼效果指標測定
1.3.2 .1 脫殼時間的測定
參考汪蘭等[16]的研究方法,對扇貝分別進行手工脫殼、蒸煮脫殼和超高壓脫殼,記錄扇貝脫殼所用的時間,并計算出平均每只扇貝的脫殼時間。
1.3.2 .2 得肉率的測定
參考王敏[17]的研究方法,在超高壓處理后,貝肉用濾紙吸干并稱質量,按式(1)和式(2)分別計算脫殼后貝肉與貝殼的質量比(Rm-s)和脫殼后貝肉與整個扇貝的質量比(Rm-t),通過Rm-s和Rm-t反映貝肉產出率。
式中:mm為脫殼后貝肉質量/g;ms為脫殼后貝殼質量/g;mt為脫殼前整個扇貝質量/g。
1.3.3 肌原纖維蛋白特性及結構的測定
1.3.3 .1 肌原纖維蛋白的提取與質量濃度的測定
參照崔燕等[18]的方法對貝肉肌原纖維蛋白進行提取與質量濃度的測定。
1.3.3 .2 肌原纖維蛋白總巰基含量的測定
參考Benjakul等[19]的方法測定總巰基含量,并略作修改。取0.25 mL肌原纖維蛋白溶液,加入2.5 mL 0.1 mol/L磷酸鹽緩沖液(含8 mol/L尿素,pH 8.0),充分混勻后,分別加入50 μL 0.01 mol/L的5,5'-二硫代雙(2-硝基苯甲酸)溶液,在40 ℃條件下水浴15 min,測412 nm波長處吸光度。使用摩爾消光系數13 600 L/(mol·cm)計算總巰基含量。
1.3.3 .3 肌原纖維蛋白羰基含量的測定
參考Jia Guoliang等[20]的方法,對貝肉肌原纖維蛋白的羰基含量進行測定。
1.3.3 .4 肌原纖維蛋白表面疏水性的測定
參考崔燕等[18]的方法,對貝肉肌原纖維蛋白的表面疏水性進行測定。
1.3.3 .5 Ca2+-ATPase活力的測定
參考閆春子等[21]的方法,對貝肉肌原纖維蛋白的Ca2+-ATPase活力進行測定。
1.3.3 .6 圓二色光譜分析
取0.1 mg/mL提取的肌原纖維蛋白樣品,經0.45 μm微孔濾膜過濾后進行圓二色光譜掃描,以超純水作為空白。測定條件:0.1 cm石英樣品池,光徑0.1 cm、帶寬1 nm、測定溫度25 ℃、波長范圍190~250 nm、掃描速率0.5 nm/s,用平均殘基橢圓值[θ]表示圓二色光譜數據,單位為deg·cm2/dmol。通過儀器自帶的軟件計算出各條件下肌原纖維蛋白的二級結構單元的相對含量。
1.4 數據處理與分析
每組實驗平行3 次,結果以平均值±標準偏差來表示。應用Origin Pro 8.5軟件繪圖,通過SPSS 17.0進行單因素方差分析,以P<0.05表示差異顯著。
2 結果與分析
2.1 超高壓對蝦夷扇貝脫殼效果的影響
經過超高壓脫殼后得到的貝肉產出率可通過得肉率Rm-s和Rm-t反映,其中Rm-s表示脫殼后貝肉與貝殼的質量比,反映貝肉的完整性,而Rm-t則表示脫殼后貝肉與整個扇貝的質量比,反映貝肉的占比[17]。扇貝脫殼效率由脫殼時間反映,脫殼時間直接對扇貝的脫殼成本和生產效率產生影響[16]。表1展示了不同壓力條件下超高壓對扇貝脫殼效果的影響,可以看出,與蒸煮脫殼和手工脫殼相比,超高壓處理組脫殼時間顯著縮短(P<0.05),并且得肉率相對較高。在實驗條件范圍內,隨著超高壓壓力的增加,脫殼時間先縮短后延長,而扇貝的得肉率則先提高后下降。當壓力增加至250 MPa時,扇貝的脫殼時間最短,僅為7.25 s,而得肉率Rm-s和Rm-t最高,分別達88.55%和65.86%。分析原因可能是殼肉之間的組織蛋白因超高壓作用產生變性,在一定范圍內,隨著壓力的不斷增大,蛋白變性程度越大,殼肉分離更容易,得肉率也隨之提升。超過250 MPa,扇貝得肉率呈下降趨勢,脫殼時間延長。原因可能是過高的壓力造成貝肉肌纖維蛋白結構被破壞,肌肉出現破裂損失,從而導致扇貝得肉率下降[8]。綜上所述,適當的壓力條件能夠提高扇貝的得肉率和縮短脫殼時間。
2.2 超高壓對蝦夷扇貝貝肉肌原纖維蛋白氧化的影響
2.2.1 超高壓對肌原纖維蛋白質量濃度的影響
如圖1所示,與手工脫殼相比,超高壓和蒸煮處理均使貝肉肌原纖維蛋白質量濃度顯著下降,其中蒸煮脫殼組貝肉肌原纖維蛋白質量濃度下降最為顯著(P<0.05),僅為手工脫殼組的49.73%。說明蒸煮處理嚴重破壞了蛋白空間構象,使蛋白變性。與蒸煮處理相比,超高壓處理相對溫和,能夠降低貝肉肌原纖維蛋白的損失。且隨著壓力的增加,貝肉肌原纖維蛋白質量濃度逐漸降低,超過250 MPa時,肌原纖維蛋白質量濃度下降趨勢加快。王芝妍等[7]研究了超高壓對中華管鞭蝦蝦仁肌原纖維蛋白質量濃度的影響,結果表明當超高壓壓力大于200 MPa時,肌原纖維蛋白質量濃度逐漸下降,這與本實驗結果類似。葉韜等[14]在研究超高壓對小龍蝦蝦仁肌原纖維蛋白質量濃度影響時亦得到類似結果,認為超高壓會引起蛋白質變性和聚集,使肌原纖維蛋白的鹽溶性降低,導致肌原纖維蛋白質量濃度下降。

圖1 不同處理條件下蝦夷扇貝貝肉肌原纖維蛋白質量濃度的變化Fig.1 Changes in myofibrillar protein content in the muscle of Patinopecten yessoensis subjected to different treatments
2.2.2 超高壓對肌原纖維蛋白總巰基含量的影響
由圖2可知,與手工脫殼組相比,蒸煮脫殼組的總巰基含量急劇下降,為35.3 nmol/mg,降低了29.54%,可能是加熱破壞蛋白分子之間的作用力,巰基暴露后易被氧化為二硫鍵,使蛋白發(fā)生聚集和變性,引起巰基含量下降[22]。當壓力為150 MPa時,總巰基含量與手工脫殼組差異不顯著(P>0.05)。壓力超過250 MPa時,其總巰基含量發(fā)生顯著變化(P<0.05)。說明隨著壓力的增大,原本位于分子內部的巰基在壓力的作用下暴露出來,從而導致氧化,因此總巰基含量減少。隨著壓力的增大,扇貝貝肉肌原纖維蛋白總巰基含量總體呈下降趨勢。這與周果等[15]的研究結果相似。

圖2 不同處理條件下蝦夷扇貝貝肉肌原纖維蛋白總巰基含量的變化Fig.2 Changes in total sulfhydryl content of myofibrillar protein in the muscle of Patinopecten yessoensis subjected to different treatments
2.2.3 超高壓對肌原纖維蛋白羰基含量的影響
羰基的形成由自由基氧化修飾蛋白質所引起,其含量變化可表征蛋白質的氧化變性程度,含量越高表示蛋白氧化程度越高[23]。由圖3可知,超高壓和蒸煮脫殼處理后肌原纖維蛋白的羰基含量均顯著高于手工脫殼組(P<0.05),蒸煮脫殼組羰基含量最高,比手工脫殼組升高了2.67 倍。羰基含量隨著壓力的增大逐漸上升,說明貝肉肌原纖維蛋白氧化變性程度也隨之更高。這與郭麗萍[24]的研究結果相似。
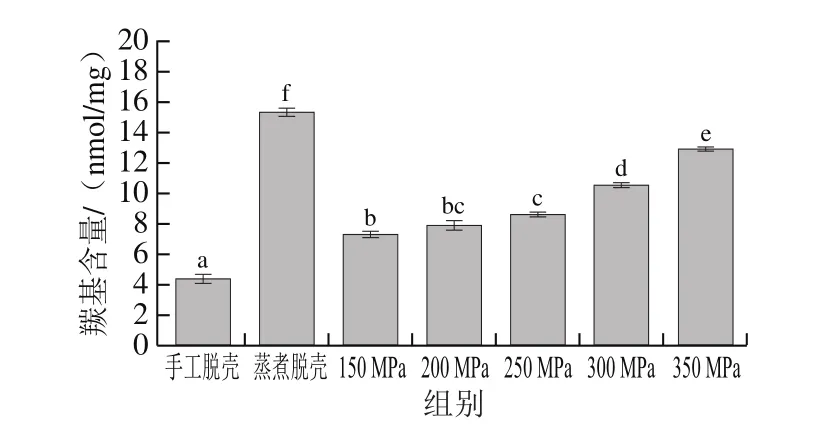
圖3 不同處理條件下蝦夷扇貝貝肉肌原纖維蛋白羰基含量的變化Fig.3 Changes in carbonyl content of myofibrillar protein in the muscle of Patinopecten yessoensis subjected to different treatments
2.2.4 超高壓對肌原纖維蛋白表面疏水性的影響
蛋白質分子內部的疏水基團暴露情況可由表面疏水性反映,表面疏水性越大說明基團暴露越多[25]。由圖4可知,與手工脫殼組相比,150、200、250、300、350 MPa處理后扇貝貝肉肌原纖維溴酚藍結合量分別增加了0.30、0.34、0.39、0.51、0.70 倍(P<0.05),說明扇貝的疏水基團多數位于蛋白質分子內部,表面疏水性低,在超高壓作用下,蛋白質分子伸展,基團逐漸外露,從而使表面疏水性上升。蒸煮脫殼組相較手工脫殼組,其表面疏水性增加了0.91 倍。這與李長樂[26]的研究結果相似。

圖4 不同處理條件下蝦夷扇貝貝肉肌原纖維蛋白表面疏水性的變化Fig.4 Changes in surface hydrophobicity of myofibrillar protein in the muscle of Patinopecten yessoensis subjected to different treatments
2.2.5 超高壓對肌原纖維蛋白Ca2+-ATPase活力的影響
肌球蛋白結構的完整性可由Ca2+-ATPase活性體現,肌球蛋白的穩(wěn)定程度會對肌原纖維蛋白的穩(wěn)定性產生一定影響[15]。由圖5可知,超高壓處理組隨著壓力的升高,其肌原纖維蛋白的Ca2+-ATPase活力呈下降趨勢。可能原因是超高壓處理使貝肉肌球蛋白頭部空間構象發(fā)生變化,造成Ca2+-ATPase活力降低。超高壓處理組在150 MPa時Ca2+-ATPase活力與手工脫殼組無顯著差異(P>0.05),但超過250 MPa時Ca2+-ATPase活力與手工處脫殼相比顯著降低(P<0.05),其中350 MPa時Ca2+-ATPase活力比手工脫殼組降低了37.35%,說明壓力過高的情況下Ca2+-ATPase易失活。蒸煮脫殼組的Ca2+-ATPase活力急劇下降,比手工脫殼組降低了45.78%。這與閆春子等[21]的研究結果相似。

圖5 不同處理條件下蝦夷扇貝貝肉肌原纖維蛋白Ca2+-ATPase活力的變化Fig.5 Changes in Ca2+-ATPase activity of myofibrillar protein in the muscle of Patinopecten yessoensis subjected to different treatments
2.3 超高壓對蝦類扇貝貝肉肌原纖維蛋白二級結構的影響
圓二色光譜作為一種研究蛋白質二級結構的方法,不僅快捷有效,而且能夠顯示生物大分子主鏈構象信息[27-28]。由圖6可知,手工脫殼組扇貝貝肉肌原纖維蛋白在209 nm、221 nm波長附近出現兩個明顯的負峰,其主要由α-螺旋結構引起,是α-螺旋結構的特征吸收峰[29-30],說明α-螺旋結構豐富。蒸煮脫殼組在209、221 nm波長附近出現的兩個負峰強度較弱,說明α-螺旋含量較低,蒸煮處理對肌原纖維蛋白二級結構產生影響較大。超高壓組在192 nm波長附近出現正峰,在209 nm、221 nm波長附近出現兩個負峰。在150~350 MPa處理3 min條件下,蛋白圓二色光譜中α-螺旋對應的負峰強度降低,說明超高壓破壞了蛋白的有序結構,α-螺旋結構部分喪失。
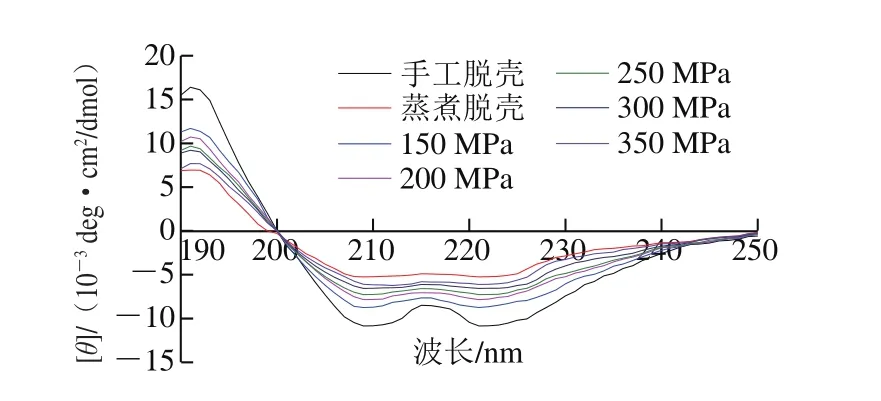
圖6 不同處理條件下蝦夷扇貝貝肉肌原纖維蛋白圓二色光譜Fig.6 Circular dichroism spectra of myofibrillar protein in the muscle of Patinopecten yessoensis subjected to different treatments
蛋白質二級結構是多肽鏈利用氫鍵進行折疊或盤旋,形成規(guī)則性和周期性的結構,包括α-螺旋、β-折疊、β-轉角和無規(guī)卷曲結構這幾種[31-32]。由圖7可知,與手工脫殼組相比,超高壓脫殼和蒸煮脫殼處理均對扇貝貝肉肌原纖維蛋白的構象單元組成產生一定程度的影響。在超高壓處理組中,150 MPa處理組肌原纖維蛋白中β-轉角、β-折疊與手工脫殼組相對含量基本接近。隨著施加壓力由150 MPa增至350 MPa,肌原纖維蛋白中α-螺旋和β-轉角相對含量逐漸降低、無規(guī)卷曲和β-折疊相對含量逐漸增加,說明壓力的增加使其結構緊密程度下降,增加了其柔性,同時維持蛋白質分子內氫鍵結合作用減弱,其蛋白質分子展開程度增加,從而對肌原纖維蛋白的二級結構產生一定影響[33-34]。無規(guī)卷曲相對含量的增加可能與蛋白分子中α-螺旋的展開有關[35]。當壓力達到250 MPa后,隨著壓力繼續(xù)加大,α-螺旋相對含量下降,β-轉角相對含量降低的幅度較小,基本無明顯變化。相比而言,蒸煮脫殼組肌原纖維蛋白中的α-螺旋相對含量最低,無規(guī)卷曲相對含量高于手工脫殼組,表明蛋白緊密的螺旋結構向無規(guī)則狀態(tài)變化,引起蛋白質的變性,蒸煮處理增強了扇貝貝肉肌原纖維蛋白的無序程度,對貝肉肌原纖維蛋白二級結構產生較大的破壞,超高壓狀態(tài)下其二級結構相對更為有序。綜上所述,超高壓處理會對蛋白構象產生一定影響。

圖7 不同處理條件下蝦夷扇貝貝肉肌原纖維蛋白二級結構含量的變化Fig.7 Changes in secondary structure contents of myofibrillar protein in the muscle of Patinopecten yessoensis subjected to different treatments
3 結 論
超高壓處理對蝦夷扇貝閉殼肌肌原纖維蛋白的生化特性和二級結構造成了一定影響。綜合分析超高壓處理后扇貝的脫殼效率以及貝肉肌原纖維蛋白的質量濃度、總巰基含量、羰基含量、表面疏水性、Ca2+-ATPase活力等指標,可以看出,當壓力處于150 MPa和200 MPa時,超高壓處理對貝肉肌原纖維蛋白的影響相對較低,在一定程度上維持了蛋白的功能特性。隨著壓力的升高,圓二色光譜圖中的兩個負峰逐漸消失,α-螺旋相對含量逐漸降低,當壓力超過250 MPa時,α-螺旋相對含量呈下降趨勢。壓力增大時蛋白從緊密的螺旋結構向無規(guī)則狀態(tài)變化,蛋白分子內疏水基團暴露,導致蛋白質發(fā)生一定程度的氧化、變性及降解,影響了蛋白構象。綜上所述,適宜的超高壓條件為250 MPa保壓處理3 min。本實驗為研究扇貝貝肉肌原纖維蛋白的生化特性及結構提供了一定的參考,進一步拓寬了蝦夷扇貝及相關產品研發(fā)思路。
