miR-223在甲型流感病毒誘導(dǎo)的肺上皮細(xì)胞炎癥中的作用機制
2021-07-02 07:23:40劉翠翠尚立群段進進陳瑞琳張永慶楊淑梅
安徽醫(yī)科大學(xué)學(xué)報 2021年6期
劉翠翠,尚立群,段進進,陳瑞琳,張永慶,楊淑梅
NOD樣受體蛋白(NOD-like receptor protein,NLRP)家族是NLR的重要成員,其中NOD樣受體蛋白3(NLRP3)炎癥小體是由NLRP3、凋亡相關(guān)斑點樣蛋白(apoptosis-associated speck-like protein containing CARD,ASC)和半胱氨酸天冬氨酸蛋白酶1前體(pro-cysteine-aspartic acid protease 1,Pro-Caspase-1)組裝成的多蛋白復(fù)合體,可將Pro-Caspase-1剪切為有活性的Caspase-1,進而剪切白細(xì)胞介素前體(pro-interleukin,Pro-IL)包括Pro-IL-1β、Pro-IL-18和Pro-IL-33形成有活性的成熟IL-1β、IL-18和IL-33,分泌至細(xì)胞外,介導(dǎo)炎癥反應(yīng)和誘導(dǎo)免疫病理損傷。
微小RNA(microRNA,miRNA)是一類長度約為20~24個核苷酸的內(nèi)源小RNA,其介導(dǎo)的轉(zhuǎn)錄后調(diào)控在基因表達的控制中發(fā)揮重要作用。研究表明,微小RNA-223(microRNA-233,miR-223)能夠特異性作用于NLRP
3 mRNA的3′-UTR區(qū),從而抑制NLRP3的翻譯表達。在病毒入侵時,為了避免引起免疫反應(yīng),機體會釋放miRNA抑制NLRP3炎癥小體的激活。目前,關(guān)于miR-223在流感病毒引起的肺部損傷中作用的研究報道較少。該研究旨在探究miR-223對流感病毒引起的肺部損傷的作用及其機制,為臨床上流感的治療提供新的理論依據(jù)。1 材料與方法
1
.1 材料
收集甲型流感患者32例和正常受試者32例的血清樣本,所有患者均知情同意,且本研究已獲國家科研倫理委員會批準(zhǔn)。正常人肺上皮細(xì)胞(bronchial epithelial cells, BEAS-2B)購自中國科學(xué)院細(xì)胞庫,以及甲型流感病毒(influenza A virus, IAV,A/Puerto Rico/8/34/H1N1)(于9~11 d的可育SPF雞蛋的尿囊內(nèi)繁殖)。RPMI 1640培養(yǎng)基、胎牛血清購自美國HyClone公司;TRIzol、反轉(zhuǎn)錄試劑盒和實時熒光定量聚合酶鏈反應(yīng)(RT-qPCR)試劑盒購自美國Promega公司;BCA蛋白質(zhì)定量檢測試劑盒購自美國Thermo Fisher Scientific公司;兔抗鼠NLRP3、兔抗鼠Bax、兔抗鼠Caspase-3、兔抗鼠Bcl-2和鼠抗GAPDH購自美國Abcam公司;轉(zhuǎn)染試劑Lipofectamine3000購自美國Invitrogen公司;PCR引物、miR-223 mimic以及陰性對照NC mimic由北京擎科生物科技有限公司設(shè)計、合成。1
.2 實驗方法
1
.2
.1
細(xì)胞培養(yǎng) 本研究所有細(xì)胞常規(guī)培養(yǎng)于RPMI 1640培養(yǎng)液(含10%胎牛血清,100 U/ml青霉素及0.1 mg/ml鏈霉素),置于37 ℃、5% CO的細(xì)胞培養(yǎng)箱,隔天換液,細(xì)胞融合至80%~90%時傳代,傳至第3代進行后續(xù)實驗。1
.2
.2
細(xì)胞處理 使用犬腎上皮細(xì)胞系MDCK來測定病毒的滴度,感染復(fù)數(shù)(MOI)為1。使用IAV病毒侵染BEAS-2B細(xì)胞,時間分別為12、24、36、48 h,測定細(xì)胞活力,后選取最適處理時間(36 h)進行后續(xù)實驗。使用RT-qPCR檢測不同處理組細(xì)胞中miR-223或NLRP3的表達情況。1
.2
.3
實驗分組 將細(xì)胞分為未處理組、IAV組、IAV+NC mimic組、IAV+miR-223 mimic組。IAV病毒侵染的BEAS-2B細(xì)胞為IAV組;未感染IAV病毒的BEAS-2B細(xì)胞為未處理組;IAV病毒侵染BEAS-2B細(xì)胞36 h后用Lipofectamine3000將陰性對照NC mimic以及miR-223 mimic分別轉(zhuǎn)染至BEAS-2B細(xì)胞中,分別為IAV+NC mimic組和IAV+miR-223 mimic組。1
.2
.4
RT-qPCR檢測mRNA表達 TRIzol法提取BEAS-2B細(xì)胞中總RNA,進行反轉(zhuǎn)錄合成cDNA。RT-qPCR檢測miR-223基因的mRNA表達,β-actin為內(nèi)參。引物序列如下:miR-223正向序列為5′-ACGTTAGCTACACTTGCGTT-3′,反向序列為5′-AGTGCCGTTGCATACAAAGG-3′;β-actin正向序列為5′-CTCACCATGGATGATGATATCGC-3′,反向序列為5′-AGGAATCCTTCTGACCACTGC-3′。以2法計算基因相對表達量。1
.2
.5
Western blot檢測蛋白表達 消化對數(shù)生長期的BEAS-2B細(xì)胞,接種于25 cm培養(yǎng)瓶中,細(xì)胞融合至80%~90%時進行細(xì)胞轉(zhuǎn)染。48 h后收集各組細(xì)胞,收集細(xì)胞上清液,并消化下貼壁細(xì)胞,加RIPA裂解液于冰上裂解10 min,4 ℃、12 000 r/min離心30 min(離心半徑9.5 cm)。上清液用BCA蛋白質(zhì)定量檢測試劑盒測定蛋白含量后,取30 μg蛋白進行SDS-PAGE凝膠電泳,130 V恒壓電泳2 h;濕轉(zhuǎn)法將蛋白轉(zhuǎn)移至PVDF膜上;室溫下以含5%脫脂奶粉的Tris緩沖液封閉PVDF膜1 h,沖洗后分別與一抗孵育,4 ℃過夜;TBST清洗膜8 min,重復(fù)4次,加相應(yīng)的辣根過氧化物酶(horseradish peroxidase,HRP)標(biāo)記的二抗,室溫孵育2 h;TBST重復(fù)清洗膜4次,ECL試劑盒檢測蛋白表達。1
.2
.6
細(xì)胞活力的檢測 將各組細(xì)胞以1×10個/孔的密度接種于96孔板中,于37 ℃、5% CO培養(yǎng)箱中培養(yǎng)24 h;向培養(yǎng)板中加入IAV病毒分別于37 ℃、5% CO培養(yǎng)箱中培養(yǎng)12、24、36、48 h,每個分組設(shè)3個復(fù)孔并設(shè)空白對照,每孔加入20 μl CCK-8溶液,孵育24 h;酶標(biāo)儀測定450 nm處的OD值。以上實驗重復(fù)3次,取平均值。1
.2
.7
細(xì)胞凋亡率的測定 將miR-223 mimic以及陰性對照NC mimic分別轉(zhuǎn)染BEAS-2B細(xì)胞,于37 ℃、5% CO條件下培養(yǎng)48 h;將細(xì)胞以1×10個/孔的密度接種在6孔板中。根據(jù)細(xì)胞凋亡檢測試劑盒的說明,用冷的PBS將細(xì)胞洗滌2次,然后用細(xì)胞凋亡檢測試劑盒中的膜聯(lián)蛋白V-FITC/PI染色。細(xì)胞凋亡率用流式細(xì)胞儀分析。以上實驗重復(fù)3次。1
.2
.8
活性氧(reactive oxygen species,ROS)的測定 將BEAS-2B細(xì)胞以1×10個/孔的密度接種于6孔板中,于37 ℃、5% CO培養(yǎng)箱中培養(yǎng)24 h;收集細(xì)胞,在37 ℃下用ROS指示劑DCFH-DA(10 μmol/L)在PBS中培養(yǎng)30 min。使用流式細(xì)胞儀分析熒光。實驗獨立重復(fù)3次。1
.2
.9
miR-223與NLRP3表達量的相關(guān)性分析 繪制散點圖來判斷miR-223與NLRP3表達量之間的關(guān)系形態(tài)。相關(guān)系數(shù)根據(jù)樣本數(shù)據(jù)計算的miR-223與NLRP3表達量之間線性關(guān)系強度的統(tǒng)計量。具體計算公式:r
=Cov
(x
,y
)/σ
σ
。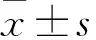
2 結(jié)果
2.1 miR-223在IAV感染的患者血清和細(xì)胞中的表達
RT-qPCR結(jié)果顯示,miR-223在流感患者血清中的相對表達量低于正常受試者中的表達,而NLRP
3在流感患者中的相對表達量高于正常受試者中的表達(圖1A、B)。進一步的研究表明,BEAS-2B細(xì)胞經(jīng)IAV處理后,細(xì)胞活力隨時間降低(圖1C)。此外,用IAV處理BEAS-2B細(xì)胞后,miR-223的相對表達量降低,而NLRP
3的相對表達量增加,且具有時間依賴性(圖1D、E)。
圖1 miR-223在IAV感染的患者血清和細(xì)胞中的表達
2.2 過表達miR-223對BEAS-2B細(xì)胞的炎癥和氧化應(yīng)激的影響
與未處理組相比,IAV處理組炎癥因子IL-6、IL-1β和TNF-α的水平顯著升高;而將IAV處理的細(xì)胞轉(zhuǎn)染miR-223 mimic后,炎癥因子的水平降低(F
=5.269,P
=0.031 1,圖2A~C)。此外,與IVA組或IVA+NC mimic組相比,IAV+miR-223 mimic組細(xì)胞內(nèi)ROS水平顯著降低(F
=7.358,P
=0.021 6,圖2D),同樣細(xì)胞內(nèi)NAD的水平也明顯下降(F
=15.872,P
=0.009 5,圖2E)。
圖2 過表達miR-223對BEAS-2B細(xì)胞炎癥和氧化應(yīng)激的影響
2.3 過表達miR-223對BEAS-2B細(xì)胞凋亡的影響
當(dāng)細(xì)胞經(jīng)IAV處理后,細(xì)胞活力降低,而轉(zhuǎn)染miR-223 mimic使IAV處理的細(xì)胞活力顯著升高(F
=3.125,P
=0.042 3,圖3A);此外,與IAV組相比,IAV+miR-223 mimic組細(xì)胞的凋亡率降低(F
=13.687,P
=0.006 5,圖3B),且細(xì)胞中Caspase-3和Caspase-9的蛋白表達水平和凋亡相關(guān)蛋白Bax/Bal-2的比值降低(F
=5.544,P
=0.026 8,圖3C~E)。
圖3 過表達miR-223對BEAS-2B細(xì)胞凋亡的影響
2.4 過表達miR-223對NLRP3的表達以及TLR4/NF-κB通路蛋白表達的影響
與IAV組相比,IAV+NC mimic組NLR3P蛋白表達水平降低(F
=6.766,P
=0.041 1,圖4A、B);此外,與IAV組相比,IAV+NC mimic組TLR4和p-p65蛋白水平也顯著下降(F
=10.264,P
=0.008 7,圖4C、D);通過對32個流感患者血清中miR-223和NLRP3表達量的相關(guān)性分析顯示,二者表達量呈負(fù)相關(guān)(F
=8.967,P
=0.025 4,圖4E)。
圖4 過表達miR-223對NLRP3的表達以及TLR4/NF-κB通路蛋白表達的影響
2.5 miR-223保護IAV誘導(dǎo)的BEAS-2B免受損傷的調(diào)控機制
CCK-8檢測結(jié)果顯示,與IAV組相比,IAV+miR-223 mimic組的細(xì)胞活力增加;IAV+pcDNA3.1-TLR4組和IAV+pcDNA3.1-NLRP3組細(xì)胞活力均減弱(F
=12.364,P
=0.0268,圖5A),且IL-1β和ROS的表達水平增加,且細(xì)胞凋亡率升高。IAV+pcDNA3.1-NLRP3組和IAV+ miR-223 mimic+pcDNA3.1-TLR4組IL-1β、ROS水平和細(xì)胞凋亡率顯著低于IAV+miR-223 mimic組,與IAV組無顯著性差異(F
=2.168,P
=0.048 7,圖5B~D);此外,與IAV組相比,IAV+miR-223 mimic組細(xì)胞中Caspase-3的表達水平明顯降低;IAV+pcDNA3.1-NLRP3組和IAV+ miR-223 mimic+pcDNA3.1-TLR4組Caspase-3的表達量顯著高于IAV+miR-223 mimic組,與IAV組無顯著性差異(F
=11.257,P
=0.013 6,圖5E)。
圖5 TLR4/NF-κB通路和NLRP3參與miR-223對IAV誘導(dǎo)的細(xì)胞損傷的保護機制
3 討論
IAV可引起反復(fù)流行,造成人類高發(fā)病率和病死率。因此深入研究IAV的感染致病及免疫反應(yīng)的機制,對研發(fā)流感病毒疫苗和抗病毒藥物具有深遠意義。該研究表明miR-223在IAV感染的患者血清細(xì)胞中的表達均下調(diào),提示miR-223可能與IAV感染過程具有一定的相關(guān)性。miR-223廣泛參與調(diào)控細(xì)胞的生物學(xué)過程和多種病理過程,如癌癥、自身免疫性疾病和炎癥性疾病。該研究表明過表達miR-223可抑制IAV誘導(dǎo)的BEAS-2B細(xì)胞炎癥,這與郝俊芳 等在LPS誘導(dǎo)的小鼠乳腺上皮細(xì)胞所得到的結(jié)論一致。李蕓蕓 等指出miR-223可顯著降低心臟缺血再灌注后心機細(xì)胞的氧化應(yīng)激反應(yīng)。此外,miR-223還可抑制糖基化終產(chǎn)物誘導(dǎo)的人脂肪間充質(zhì)干細(xì)胞凋亡。與前人的研究一致,該研究顯示過表達miR-223降低IAV誘導(dǎo)的BEAS-2B的細(xì)胞凋亡率。以上表明,IAV感染可使miR-223表達下調(diào),過表達miR-223可緩解 IAV誘導(dǎo)的BEAS-2B細(xì)胞損傷。
NLRP3炎癥小體信號通路在IAV感染后引起炎癥反應(yīng),并可釋放炎癥因子放大炎癥反應(yīng)。當(dāng)IAV感染引起炎癥小體激活時,大量的炎性因子會誘導(dǎo)嚴(yán)重肺臟免疫病理損傷,如急性肺損傷和急性呼吸窘迫綜合征。Chung et al證實NLRP3炎癥小體活化后釋放的IL-1β破壞上皮細(xì)胞,并進一步誘導(dǎo)IL-6、TNF-α的產(chǎn)生,進而誘發(fā)細(xì)胞因子引起嚴(yán)重的急性肺損傷。越來越多的研究表明,miR-223能夠抑制NLRP3炎癥小體活化,進一步抑制炎癥性疾病的進展。此外,激活miR-223的轉(zhuǎn)錄顯著抑制炎性介質(zhì)NLRP3的活化,并降低IL-1β產(chǎn)生來防止炎癥損傷。Sha et al的研究表明,NLRP3在miR-223敲除小鼠表達水平升高,導(dǎo)致IL-1β產(chǎn)生和腸道炎癥易感性增加。該研究顯示miR-223抑制了NLRP3的活化以及TLR4/NF-κB信號通路的激活,且TLR4/NF-κB通路和NLRP3參與了miR-223對IAV誘導(dǎo)的細(xì)胞損傷的保護機制,表明miR-223可能是通過抑制TLR4/NF-κB信號通路并抑制NLRP3炎癥小體的活化來抑制IAV感染造成的BEAS-2B細(xì)胞損傷。
