ORP8對結直腸癌DLD-1細胞增殖、遷移和侵襲能力的影響
2021-07-02 07:23:42潘琪偉陳韋璁裴冬生王慶苓
安徽醫(yī)科大學學報 2021年6期
潘琪偉,陳韋璁,裴冬生,王慶苓
結直腸癌(colorectal cancer,CRC)是常見的消化道惡性腫瘤,在全球范圍內(nèi),結直腸癌是男性和女性的第三大癌癥,每年超過69萬例死亡,且該病的患者群逐漸趨向年輕化。因此,深入探討結直腸癌發(fā)病及轉移的分子機制,尋找新的治療靶點,對治療結直腸癌具有重要的意義。氧化固醇結合蛋白相關蛋白8(oxysterol binding protein-related protein 8,ORP8)隸屬于一類存在于真核細胞內(nèi)的參與脂質(zhì)代謝的非囊泡運輸?shù)鞍踪|(zhì)——氧化固醇結合蛋白(oxysterol binding protein,OSBP)家族,已有研究表明,ORP8在肝癌、胃癌、非小細胞型肺癌等腫瘤中介導的信號轉導在腫瘤細胞的增殖、凋亡的過程中起著重要作用,然而至今,結直腸癌患者中ORP8的表達水平及其對結直腸癌細胞的增殖、遷移和侵襲能力影響的研究尚無報道。該研究旨在通過從體外低表達或過表達 ORP8在結直腸癌細胞中的水平來探討其對結直腸癌細胞增殖、遷移和侵襲的影響及其分子機制。
1 材料與方法
1.1 實驗細胞
人結直腸癌DLD-1細胞購自中國科學院細胞研究所。1.2 ORP8表達質(zhì)粒和ORP8 siRNA片段
針對人ORP8 mRNA的表達質(zhì)粒pcDNA3.1-ORP8由本科室保存。針對人ORP8 mRNA的siRNA片段5′-GAGUGGUCUUGCAAAUUAUTT-3′及無關序列siRNA購自吉瑪生物科技有限公司。1.3 實驗試劑
兔抗人細胞外蛋白調(diào)節(jié)激酶(extracellular regulated protein kinases,ERK1/2)多克隆抗體、兔抗人磷酸化細胞外蛋白調(diào)節(jié)激酶(phosphorylated-extracellular regulated protein kinases,p-ERK1/2)多克隆抗體、兔抗人Vimentin多克隆抗體購自美國Cell Signaling Technology公司;鼠抗人ORP8單克隆抗體、鼠抗人p-ERK1/2單克隆抗體購自英國Abcam公司;鼠抗人β-actin、山羊抗鼠二抗、山羊抗兔二抗、鼠抗人E-cadherin多克隆抗體、兔抗人N-cadherin多克隆抗體購自美國proteintech公司;轉染試劑Lipofectamine 2000購自美國Invitrogen公司;CCK-8試劑盒購自美國APExBio公司;Transwell小室和Matrigel膠購自美國Corning公司;RIPA裂解液和BCA蛋白濃度測定試劑盒購自江蘇凱基生物技術有限公司;ECL購自美國Millipore公司。1.4 實驗方法
1
.4
.1
細胞培養(yǎng)及分組 人結直腸癌細胞系DLD-1細胞用含10%小牛血清的DMEM于37 ℃、5% CO恒溫培養(yǎng)箱內(nèi)培養(yǎng)。細胞轉染的操作按照Lipofectamine 2000試劑盒操作說明書進行,轉染48 h后收樣。將細胞分為轉染空載質(zhì)粒的Vector組、轉染ORP8表達質(zhì)粒的ORP8組、轉染無關序列siRNA的siCtrl組以及轉染ORP8干擾序列siRNA的siORP8組。1
.4
.2
Western blot檢測ERK1/2、p-ERK1/2以及上皮-間質(zhì)轉化 (epithelial-mesenchymal transition, EMT)相關分子標志物的蛋白的表達 提取總蛋白,進行SDS-PAGE電泳,然后轉印到PVDF膜,5% 脫脂牛奶封閉2 h,一抗4 ℃孵育過夜,二抗室溫孵育2 h,然后用 ECL 試劑檢測目的條帶。1
.4
.3
CCK-8檢測細胞增殖能力 取對數(shù)生長期的細胞,調(diào)整細胞密度為3×10個/孔,接種至96孔板,培養(yǎng)24、48、72 h,每孔加入10 μl CCK-8試劑,37 ℃孵育2 h,酶標儀檢測450 nm處的吸光度值(OD值)。1
.4
.4
Transwell法檢測細胞遷移和侵襲能力 取對數(shù)生長期的細胞進行細胞計數(shù),每孔加入6×10個細胞到Transwell小室的上層,小室下層加入0.6 ml的10%血清的培養(yǎng)液,每組3個復孔,其中,侵襲組實驗前小室內(nèi)加入基質(zhì)凝膠(無血清培養(yǎng)基 ∶Matrigel膠=9 ∶1),置于37 ℃培養(yǎng)箱中過夜。遷移組培養(yǎng)24 h、侵襲組培養(yǎng)48 h后取出 Transwell 小室,用100%甲醇固定30 min,結晶紫染液染色 15 min,于倒置顯微鏡10倍鏡下每個小室隨機取10個視野拍照計數(shù)。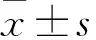
2 結果
2.1 ORP8表達質(zhì)粒和siRNA轉染DLD-1細胞后ORP8蛋白的表達
ORP8表達質(zhì)粒、siRNA轉染 DLD-1細胞 48 h后,采用 Western blot法檢測轉染效果,ORP8組ORP8表達高于Vector組,siORP8組ORP8表達低于siCtrl組,如圖1中所示。
圖1 ORP8過表達質(zhì)粒和siRNA轉染DLD-1細胞后ORP8蛋白表達情況
2.2 過表達ORP8對DLD-1細胞增殖、遷移和侵襲能力的影響
2
.2
.1
過表達ORP8對DLD-1細胞增殖的影響 CCK-8法檢測結果如圖2所示,72 h 時ORP8組的細胞 OD450值低于Vector組,差異有統(tǒng)計學意義(72 ht
值為5.709,P
<0.01)。以上實驗結果提示,過表達ORP8能夠抑制DLD-1細胞的增殖。
圖2 過表達ORP8對DLD-1細胞增殖能力的影響與Vector組比較:*P<0.05,**P<0.01
2
.2
.2
過表達ORP8對DLD-1細胞遷移和侵襲的影響 Transwell法檢測結果如圖3所示,與Vector遷移組相比,ORP8遷移組DLD-1細胞的遷移細胞數(shù)量減少(t
值為5.508,P
<0.01);與Vector侵襲組相比,ORP8侵襲組DLD-1細胞的侵襲細胞數(shù)量減少(t
值為3.823,P
<0.05)。以上實驗結果提示,過表達ORP8能夠抑制結直腸癌細胞的遷移和侵襲。
圖3 過表達ORP8對DLD-1細胞遷移和侵襲能力的影響 結晶紫染色 ×100
2.3 干擾ORP8對DLD-1細胞增殖、遷移和侵襲能力的影響
2
.3
.1
干擾ORP8對DLD-1細胞增殖的影響 CCK-8法檢測結果如圖4所示,72 h時 siORP8組的細胞 OD450值高于siCtrl組,差異有統(tǒng)計學意義(72 h時t
值為7.394,P
<0.01)。以上實驗結果提示,干擾ORP8能夠增強結直腸癌DLD-1細胞的增殖能力。
圖4 干擾ORP8對DLD-1細胞增殖能力的影響
2
.3
.2
干擾ORP8對DLD-1細胞遷移和侵襲的影響 Transwell法檢測結果如圖5所示,與siCtrl遷移組相比,siORP8遷移組DLD-1細胞的遷移細胞數(shù)量增多(t
值為7.103,P
<0.01);與siCtrl侵襲組相比,siORP8侵襲組DLD-1細胞的侵襲細胞數(shù)量增多(t
值為7.646,P
<0.01)。以上實驗結果提示,干擾ORP8能夠增強DLD-1細胞的遷移和侵襲能力。
圖5 干擾ORP8對DLD-1細胞遷移和侵襲能力的影響 結晶紫染色×100
2.4 過表達、干擾ORP8表達對DLD-1細胞ERK1/2、p-ERK1/2以及EMT相關分子標志物的蛋白表達情況的影響
Western blot檢測結果顯示:相較于Vector組,ORP8組上皮標志物 E-cadherin蛋白表達增高,間質(zhì)標志物Vimentin、N-cadherin 蛋白以及p-ERK1/2蛋白表達下降;相較于siCtrl組,siORP8組上皮標志物 E-cadherin蛋白表達下降,間質(zhì)標志物Vimentin、N-cadherin 蛋白以及p-ERK1/2蛋白表達增高。見圖6。
圖6 過表達和干擾ORP8對DLD-1細胞ERK1/2、p-ERK1/2及 EMT 相關分子標志物蛋白表達情況的影響
3 討論
ORP8主要存在于內(nèi)質(zhì)網(wǎng)與質(zhì)膜或者線粒體的膜接觸位點上,其主要功能目前被證明是從內(nèi)質(zhì)網(wǎng)到細胞膜或者線粒體運輸磷脂酰絲氨酸,并從細胞膜或者線粒體向內(nèi)質(zhì)網(wǎng)反方向運輸磷脂酰肌醇4-磷酸,導致磷脂酰肌醇4-磷酸被磷脂酰肌醇3-磷酸酶水解。Zhong et al報道了ORP8在臨床人類肝癌組織中下調(diào),而miR-143則被報道上調(diào)ORP8的表達,ORP8可能通過介導細胞表面死亡受體的易位參與調(diào)控了原發(fā)性肝癌細胞的凋亡。Guo et al報道了過表達ORP8在體內(nèi)抑制胃癌細胞的增殖和移植瘤的生長。內(nèi)質(zhì)網(wǎng)應激的誘導、Wnt信號的抑制和凋亡細胞的死亡與ORP8誘導的胃癌細胞增殖的抑制有關。最新的研究結果顯示,ORP8過表達通過釋放細胞色素c從線粒體進入細胞質(zhì)誘導非小細胞肺癌細胞凋亡。上述研究提示ORP8在腫瘤中可能具有重要作用。本實驗旨在探討ORP8表達水平的改變對結直腸癌細胞DLD-1增殖、遷移和侵襲的影響,結果顯示,過表達ORP8可抑制結直腸癌DLD-1細胞增殖、遷移和侵襲的能力,反之,干擾了結直腸癌DLD-1細胞ORP8之后,癌細胞的增殖、遷移和侵襲能力增強,提示ORP8對結直腸癌細胞增殖、遷移和侵襲的能力具有重要的抑制作用。
本研究顯示,在與細胞遷移侵襲實驗條件相同的情況下,干擾ORP8可促進上皮標志物 E-cadherin蛋白含量減少,而間質(zhì)標志物 Vimentin、N-cadherin 蛋白含量增高;過表達ORP8后結果與之相反。上述研究也進一步證實了ORP8參與調(diào)控EMT,影響結直腸癌細胞遷移侵襲。
已有多項研究表明,在結直腸癌中,通過影響ERK信號傳導可以調(diào)控EMT,另外也有研究表明,ERK信號通路激活與否影響著細胞的遷移能力,而且在Kattan et al的報道中,提及了ORP8與ERK信號通路的聯(lián)系。本研究檢測了ORP8對ERK1/2磷酸化水平的影響,結果顯示,ORP8的過表達和沉默確實引起了ERK1/2磷酸化水平的變化,提示ERK信號通路在ORP8抑制結直腸癌細胞增殖、遷移和侵襲的過程中扮演了一個重要的角色,然而其具體機制仍有待探索。
綜上所述,ORP8抑制了結直腸癌細胞DLD-1的增殖、遷移和侵襲,其作用機制可能與抑制EMT和ERK1/2磷酸化水平有關。
