植物木酚素生物轉化研究進展
2021-07-02 13:30:38馮程程張雯蔡子哲李穎汪勇張寧
食品與發酵工業 2021年11期
關鍵詞:植物
馮程程,張雯,蔡子哲,李穎,汪勇,張寧*
1(暨南大學 食品科學與工程系,廣東 廣州,510632) 2(暨南大學-薩斯喀徹溫大學“油料生物煉制與營養”聯合實驗室,廣東 廣州,510632) 3(宜春職業技術學院 護理學院,江西 宜春,336000)
植物雌激素是從植物中發現或源自植物前體的多酚類物質,結構與內源性雌激素類似,具有雌激素和抗雌激素功能[1]。根據其化學結構的不同,主要分為異黃酮類、香豆雌酚類、木酚素類、二苯乙烯類、鞣花丹寧類[2]。其中木酚素是一類由兩分子苯丙素衍生物(C6-C3)聚合而成的化合物[3],按其來源可分為植物木酚素和哺乳動物木酚素。植物中含量最多的植物木酚素是開環異落葉松樹脂酚(secoisolariciresinol,SECO)及其前體開環異落葉松樹脂酚二葡萄糖苷(secoisolariciresinol diglucoside,SDG)、羅漢松脂酚(matairesinol,MAT)。另外,松脂酚(pinoresinol,PIN)、皮樹脂醇(medioresinol,MED)、丁香脂素(syringaresinol,SYR)、異落葉松脂素(isolariciresinol,ISOLAR)、羥基羅漢松脂酚(hydroxymatairesinol,HYMAT)、落葉松樹脂醇(lariciresinol,LARI)、牛蒡子苷(arctiin,ART)、牛蒡苷元(arctigenin,ARG)、脫水開環異落葉松脂酚(anhydrosecoisolariciresinol,AHS)、芝麻素酚(sesaminol,SES)及前體芝麻素酚三糖苷(sesaminol triglucoside,STG)也是在植物中發現的木酚素[1]。植物木酚素因具有抗真菌、抗細菌、抗病毒甚至殺蟲的特性,可以保護植物免受病原體和害蟲侵害,如MAT及其相關代謝物可抑制云杉真菌生長[4]。哺乳動物木酚素也叫腸道木酚素,由植物木酚素經過腸道微生物的作用形成,主要是腸二醇(enterodiol,END)和腸內酯(enterolactone,ENL),存在于血清、血漿、尿液和糞便中。
木酚素因在預防結腸癌、乳腺癌、前列腺癌、心血管疾病和更年期綜合癥[1]等疾病中發揮作用,近年來越來越受到關注,然而植物木酚素由于其生物利用度低而具有有限的生物學特性[5-6],必須將其轉化為腸道木酚素才能在人體內發揮最大有益作用。盡管已經有了化學合成END的方法[7],但因其使用的化學試劑價格昂貴、步驟復雜且對環境有污染,實際生產中越來越傾向于生物轉化法,生物轉化法具有操作簡單、對設備要求低、反應快速等優點,具有重要的應用前景以及巨大的經濟價值和現實意義。因此,本文將主要綜述木酚素的結構、種類和分布、木酚素腸道微生物代謝機制,以及體外木酚素微生物和酶法轉化的研究進展。
1 木酚素生理功能
END和ENL與雌二醇化學結構相似,進入機體后與雌激素競爭性結合雌激素受體,產生一系列生理活性[8]。體內外試驗表明,腸道木酚素是哺乳動物芳香酶抑制劑,它通過與競爭底物雌烯二酮的結合、抑制雄性激素的合成、與性甾結合蛋白競爭性結合、促進性激素結合蛋白的合成、抑制二氫睪酮與促進性激素結合蛋白的結合及抑制人組織中5α-還原酶和17β-羥基類固醇脫氫酶的活性,進而降低血漿中雌二醇和睪丸激素的水平達到防治性激素依賴性疾病的目的。現有研究表明,腸道木酚素還具有內源性洋地黃樣及降低冠心病發作的生理活性[9]。除此之外,腸道木酚素能影響結腸、前列腺和乳腺的體外培養細胞的增殖,在體外比木酚素前體具有更強的抗氧化性能[10]。
2 木酚素分布
木酚素主要存在于植物中,在根、莖、葉、種子及果實中都能發現,軟木樹的結中存在大量的木酚素,主要是以HYMAT的形式[3]存在,如挪威云杉結中含有高達20%的木酚素(干重),其中HYMAT為主要成分。亞麻籽是最廣泛的植物木酚素來源,含量約300 mg/100 g鮮重,以SECO的糖苷形式SDG為主[11](表1)。谷物類如黑麥、小麥、蕎麥和燕麥含有中等到高等含量的木酚素,全麥亞麻籽面包和雜糧面包中含量也較高,羽衣甘藍、卷心菜、蘆筍、花椰菜、大蒜、杏、柚子和棗等蔬菜水果都含有適量的木酚素,紅茶、綠茶、咖啡、啤酒等常見飲料中也有少量木酚素[12-16]。

表1 植物性食品中植物木酚素的含量 單位:μg/100 g(濕重)
3 木酚素代謝
SETCHELL等[17]用人糞便菌培養亞麻籽可產生END和ENL,但是無菌糞便培養不能產生,且抗生素處理降低了結腸中END和ENL的產生,表明腸道微生物群負責將植物木酚素轉化為腸道木酚素,主要部位是盲腸和結腸。腸道菌群主要包括擬桿菌門、厚壁菌門、放線菌門和變形桿菌門,有超過1 000種不同的細菌種類,腸道菌群扮演著基因編碼酶的儲存庫的角色,這些酶可用于不同的代謝活動,而這是宿主本身所不具備的[2]。植物雌激素的潛在健康益生功能和生物活性很大程度上依賴于腸道微生物群,通過研究人體腸道菌群的代謝活動,可以更好地了解人類腸道菌群代謝木酚素的機制。
表2整理了能使植物木酚素轉化為腸道木酚素及其前體的各種特定腸道細菌。因為亞麻籽是木酚素最豐富的來源,大多數實驗都是使用亞麻籽為底物,亞麻籽中主要含有SECO,一般以其糖苷形式SDG存在。SDG的生物轉化通過腸道菌群中占優勢、亞優勢組之間復雜的相互作用進行,一般要經過4個步驟(圖1),轉化的第一步是從SDG中除去葡萄糖基產生SECO,這種生物轉化可以通過產β-葡萄糖苷酶的菌株進行[6]。SDG釋放葡萄糖基是益生菌增加SECO釋放、提高其結腸黏膜吸收的生物利用度、腸道微生物向END和ENL的生物轉化的重要特征[18]。水解SDG的細菌有擬桿菌,如吉氏擬桿菌(Bacteroidesdistasonis),脆弱擬桿菌(B.fragilis),卵形擬桿菌(B.ovatus)和梭菌,如耳蝸形梭菌(Clostridiumcocleatum)、多枝梭菌(C.ramosum)、噬糖梭菌(C.saccharogumia)[19-21]。RONCAGLIA等[22]證明了雙歧桿菌可將SDG水解為SECO,發現了10株能夠水解SDG的雙歧桿菌菌株,尤其是假鏈狀雙歧桿菌WC401,對水解SDG和其他糖綴合的多酚類物質具有較高的效率。

表2 使植物木酚素轉化為腸道木酚素及其前體的各種特定腸道細菌Fig.2 Various specific intestinal bacterias of transforming plant lignans into intestinal lignans and precursors
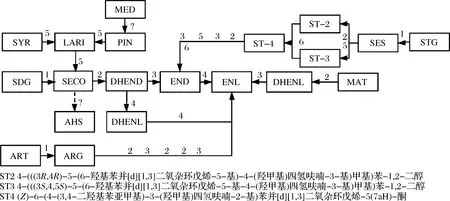
圖1 常見的植物木酚素轉化為腸道木酚素END和ENL的途徑Fig.1 Conversion pathway of common plant lignans to enterolignans END and ENL注:實線表示已知路徑,虛線表示理論路徑;反應為:1-去糖基;2-脫甲基;3-脫羥基;4-脫氫;5-還原;6-氧化;?-未知
轉化的第二步是SECO去除甲基,產生二羥基腸二醇(dihydroxyenterodiol,DHEND)。能夠使SECO去甲基的菌株有食甲基丁酸桿菌(Butyribacteriummethylotrophicum)、卡蘭德真桿菌(Eubacteriumcallanderi)、粘液真桿菌(Eubacteriumlimosum)、布勞特氏菌(Blautiaproducta)、消化鏈球菌(Peptostreptococcusproductus)[19-20]。P.productus不僅催化SECO去甲基化,而且還催化LAR、MAT、PIN和其他各種甲基化芳香化合物的去甲基化[19]。Eubacterium.sp.ARC-2可使牛蒡苷元脫甲基為二羥基腸內脂(dihydroxyenterolactone,DHENL),還可以通過去甲基化將SECO轉化為DHEND[23]。JIN等[24]發現梭菌科細菌(Clostridiaceae) END-2具有使SECO脫甲基的能力。
轉化的第三步是脫羥基,產生END。能去除芳香環上的羥基的細菌有遲緩埃格特菌(Eggerthellalenta)和梭狀芽苞桿菌(C.scindens)[20]。Lactonifactorlongoviformis可從END脫氫產生ENL[20-21]。這些反應似乎是對映體特異性的,SDG與腸道細菌共孵育后分離出的END和ENL的形式為(+)-END和(+)-ENL。而ART或松脂酚二葡萄糖苷為底物則生成(-)-END和(-)-ENL[25-26]。E.lentaSDG-2可以將(+)-DHEND轉化為(+)-END,而不能將(-)-DHEND轉化為(-)-END,人類糞便中分離出的ARC-1,不僅能將(-)-DHEND轉化為(-)-END,也能將(+)-DHENL轉化為(+)-ENL[24]。另外,DHEND可以通過脫氫作用形成內酯環,從而形成中間體DHENL,然后再脫氫生成ENL[27]。
膳食木酚素PIN也通過4種化學反應被腸道微生物轉化:首先是經過2次還原反應生成SECO,然后脫去2個甲基成為DHEND,再脫去2個羥基成為END,最后內酯化成為ENL[19,28]。BESS等[29]通過比較基因組學和轉錄組研究了4種細菌在厭氧環境生長過程中進行每種反應的能力。E.lentaDSM2243T發生催化還原和脫甲基反應,菌株產生的二芐醚還原酶(benzyl ether reductase,ber),由基因ber編碼,足夠使PIN發生還原和脫甲基反應;B.productaDSM3507把SECO代謝為DHEND,培養過程中表達最高的基因guaiacol lignan methyltransferase(glm);Gordonibacterpamelaea3C使DHEND脫羥基,生成END,培養過程中catechol lignan dehydroxylase(cldh)基因顯著上調;L.longoviformisDSM 17459T催化END的2個羥基內酯化為ENL,在END的存在下,1個包含3個基因的單一基因組位點的基因轉錄水平顯著上調,其中1個基因與主要協助轉運蛋白超家族轉運體有同源性,另外2個基因與4Fe-4S NADP-依賴型氧化還原酶基和NAD (P)依賴型短鏈脫氫酶/還原酶基因(END lactonizing enzyme,edl)具有同源性,可能是這些酶基因簇使END發生到ENL的轉化。其他分離菌株的基因簇和酶還有待鑒定,雖然目前已經分離并鑒定了大量的木酚素生物轉化和產生腸道木酚素的菌株,但對其基因及編碼酶的詳細研究并不普遍,比較基因組學和轉錄分析已被證明是鑒定木酚素生物轉化過程中所涉及的酶的有效工具。
芝麻種子中最豐富的木酚素為芝麻素酚三葡萄糖苷(sesaminol triglucoside,STG),是一種有亞甲二氧基苯基的四氫呋喃類木酚素,去糖基后生成芝麻素酚(sesaminol,SES),腸道菌群將四氫呋喃木酚素轉化為哺乳動物木酚素,涉及到糖苷的水解、亞甲基的去甲基化、二芐基丁二醇氧化為二芐基丁內酯、呋喃環的還原裂解[30]。對轉化SES的生物體的研究沒有涉及SDG轉化的生物體那么廣泛,在人類和大鼠體內以及體外培養中,SES也能轉化為END和ENL[30]。ART的苷元ARG轉化為ENL需要5個反應步驟,包括3個去甲基化反應和2個去羥基化反應[1]。
END和ENL生成后被腸道吸收,出現在血漿中,然后以葡萄糖醛酸鹽或硫酸鹽偶聯物的形式隨尿液和膽汁排出體外。QUARTIERI等[6]用5名連續7 d有亞麻籽膳食補充的受試者的糞便接種含SDG的培養物。在糞便培養物中鑒定出了與SECO、MAT和AHS有關的新型中間體,這些代謝物也以天然形式和/或以葡萄糖苷酸鹽或硫酸鹽復合物的形式隨尿液和膽汁排出體外。GAYA等[31]通過腸道菌群分析木酚素的代謝,體外發酵實驗采用14例健康成年志愿者的糞便樣品,所有受試者中均發現ENL,而END僅出現5例,木酚素代謝中沒有發現性別差異。雖然在腸道木酚素的產生中可以觀察到一些個體間的差異,但成年人類的腸道微生物在木酚素的代謝中表現出類似的行為。
4 體外木酚素微生物和酶法轉化研究進展
體外木酚素的微生物和酶法轉化研究以生產應用為目的,表3整理了與體外木酚素代謝有關的微生物和酶。杜木香等[37]利用生物酶水解SDG,結果顯示,3 mg/mL來源于綠色木霉的纖維素酶在60 ℃下反應48 h時,對SDG的水解效果最佳,此時產物SECO的得率為43.6%,相較于化學法水解得率更高,且反應后處理更為簡便、快捷。HWANG等[38]從人類糞便中分離出一株蠟樣芽孢桿菌(BacilluscereusH62L),可以釋放香豆酸葡萄糖苷(coumaric acid glucoside,CAG)、阿魏酸葡萄糖苷(ferulic acid glucoside,FAG)和SDG的糖基,但是其將SDG轉換為SECO的過程是有限的。由于蠟樣芽孢桿菌H62L葡萄糖苷酶的可逆性,SDG向SECO的生物轉化的產率極其有限,影響產率的參數包括SDG的純度和β-葡萄糖苷酶的可逆性。為了提高腸道木酚素的產量,有研究提出在較高濃度下使用高純度SDG底物,或使用變構抑制劑延緩SECO向SDG的反向反應,或對蠟樣芽孢桿菌株H62L進行轉基因處理。之后,HWANG等[39]研究了從臭豆腐水中分離出的高地芽孢桿菌(BacillusaltitudinisHK02)產生的SDG-β-葡萄糖苷酶對亞麻籽糖苷SDG、CAG和FAG 3種糖苷的去糖作用。這些酚類糖苷的脫糖率取決于糖苷元上的糖基數目。3種糖苷的酶解糖效率可達100%左右。脫脂亞麻籽粉可以增強酶的活性,葡萄糖、蔗糖、果糖、甘露醇等碳源對酶活性有明顯的抑制作用,纖維素可能是產生β-葡萄糖苷酶的良好誘導劑。ZHOU等[40]發現1株從人類腸道菌群中分離出來的菌株S1,與克雷伯氏菌屬(Klebsiella)親緣關系相近,該菌株可以通過轉化脫脂亞麻籽中前體產生SECO。

表3 與體外木酚素代謝有關的微生物和酶Table 3 List of microbial and enzyme implicated in the metabolism of lignans in vitro
GAYA等[41]研究了90種乳酸菌和雙歧桿菌中木酚素的代謝,發現大多數菌株有將SDG脫糖基為SECO的能力。還發現在一株青春雙歧桿菌(BifidobacteriumadolescentisINIA P784)作用下,能從木酚素提取物中產生1.12~4.18 μmol/L的END。然而,純木酚素SECO、MAT和SES并不能產生最終產物。BRAVO等[42]分析了70株乳酸菌、乳球菌和腸球菌產END和ENL的情況。在唾液乳桿菌(LactobacillussalivariusINIA P183、L.salivariusINIA P448)和格氏乳桿菌(L.gasseriINIA P508)中發現存在END和ENL,濃度分別約為46和6 μmol/L,且這些菌株產END的量是陽性對照B.adolescentisINIA P784產END量的10倍左右,這是首次發現單個菌株同時產生END和ENL。在此基礎上,PEIROTN等[43]在20種雙歧桿菌菌株中發現3種雙歧桿菌(BifidobacteriumbifidumINIA P466、B.catenulatumINIA P732和B.pseudolongumINIA P2)能夠從木酚素提取物中產生低水平END(2~11 μmol/L),而另一種假鏈狀雙歧桿菌(B.pseudocatenulatumINIA P946)則使SECO產生量大大增加。隨后,這3種產END的雙歧桿菌和另外3種先前被確定能產腸道木酚素的乳桿菌(L.gasseriINIA P508、L.salivariusINIA P448和L.salivariusINIA P183)分別與純SECO共同培養,得到END和ENL,而沒有代謝MAT。其中B.catenulatumP732和L.gasseriINIA P508是轉化SECO效率最高的菌株,產生濃度為2 mmol/L以上的ENL。此外,所有菌株的發酵過程中都觀察到中間化合物DHEND的形成。這項工作首次證明了雙歧桿菌和乳桿菌能夠進行完全的腸道木酚素代謝,轉化純木酚素SECO為END和ENL。
ZHU等[44]通過反復傳代和平板培養,從活性菌群中分離鑒定出3個形態各異的菌落,分別稱為ZL-I、ZL-II和ZL-III,在ZL-(I-II-III)菌株的聯合作用下,可直接從脫脂亞麻籽中產生END。3種菌株分別具有去糖基(ZL-I)、去甲基(ZL-II)和脫羥基(ZL-III)的功能。通過16Sr RNA序列分析發現它們分別與單行擬桿菌(B.uniformis)、粘液真桿菌(E.limosum)和遲緩埃格特菌(E.lenda)密切相關,這些物種都常見于人的腸道。用單行擬桿菌(B.uniformisZL-I)和粘液真桿菌(E.limosumZL-II)轉化亞麻木酚素的試驗中,在2 L的發酵體系中于24 h內約59.1%的SDG轉化為DHEND,到第6天時,發酵液中DHEND濃度達最大值,此時SDG轉化率達94.4%[45]。而與ZL-I菌株將脫脂亞麻籽中SDG轉化為SECO的比例(約80%)[46]相比,SDG轉化為END的轉化率大大提高,達到90%以上。
MUNOZ等[47]通過完全體外消化全亞麻籽和亞麻籽粉來評估干酪乳桿菌和嗜酸乳桿菌對亞麻木酚素生物可給性(bioaccessibility)的影響。干酪乳桿菌在體外消化過程中提高了亞麻籽中SDG和END的生物可給性。然而,嗜酸乳桿菌在開始的12 h內增加了END的生物可給性,48 h后無增加。2種益生菌對亞麻籽粉的消化均無明顯影響。
5 結論
木酚素廣泛存在于亞麻籽、黑麥麩皮等各種植物中,是一種植物雌激素,具有預防結腸癌、乳腺癌、前列腺癌、心血管疾病和更年期綜合癥等作用。植物木酚素需經腸道微生物多步代謝轉化為哺乳動物木酚素才能被人體吸收利用,各種木酚素的腸道代謝途徑、參與轉化的腸道微生物已有許多報道。其中,催化木酚素各轉化步驟的微生物酶系仍然有待進一步明確,這將為指導木酚素體外生物轉化提供重要理論依據。由于木酚素的生物利用度對腸道菌群健康狀況的高度依賴,木酚素體外生物轉化,尤其是體外益生菌轉化對開發高活性木酚素產品具有重要意義。對植物木酚素分布、生理功能、體內代謝吸收和體外生物轉化的研究將推動以亞麻籽粕、麩皮等農副產品為原料開發木酚素高附加值產品的深加工產業發展,實現農業廢棄物的資源化利用,具有重要社會經濟價值。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
