全外顯子組測序在自身免疫性疾病中的研究進展
2021-07-15 05:44:38馬曉敏王欣姚新生譚貴琴周芳宇魏文文于紅松
中國比較醫學雜志 2021年6期
關鍵詞:研究
馬曉敏王 欣姚新生譚貴琴周芳宇魏文文于紅松
(遵義醫科大學基礎醫學院,貴州遵義 563000)
自身免疫性疾病(autoimmune diseases,AIDs)是一類因免疫功能發生紊亂或機體自身發生免疫應答從而使得自身器官系統受損的疾病[1]。 正常機體的免疫系統具有區別“自己”和“非己”的能力,對非己抗原能夠發生免疫應答,對自身抗原則是處于無應答或微弱應答的免疫耐受狀態;如果正常的免疫耐受被打破,處于非正常免疫激活狀態的T 細胞就會持續遷延,對自身抗原產生異常的免疫應答,導致AIDs 的發生;AIDs 的主要特征是患者血清中可檢測到高濃度的自身抗體或是能夠與自身組織起反應的淋巴細胞。 研究發現,環境因素和遺傳因素可以增加普通人群患AIDs 的風險[2-4]。 大量遺傳學研究還表明,不同的 AIDs 有著共同的遺傳背景[5-7]。
全基因組關聯研究(genome-wide association studies,GWAS)、 全 外 顯 子 組 測 序 (whole exon sequencing,WES)與全基因組測序等技術的飛速發展,極大地加深了我們對AIDs 相關遺傳因素的理解[8]。 GWAS 的核心是研究分子變異和目標表型性狀之間的關聯,極大地擴大了對最常見形式AIDs 遺傳因素的理解,但只能部分解釋復雜性狀的遺傳性,不能找到真正的致病基因或者突變位點,對基因與環境因素交互作用缺乏足夠的關聯研究,GWAS 對低頻變異和罕見變異的檢測能力也十分有限[8-11]。 全基因組測序是針對個體所有基因序列進行的測序分析,其測序范圍廣、數據多、分析困難、專業性強、費用昂貴,因此目前難以大規模應用于臨床[12]。 而WES 則只是針對DNA 的外顯子區域,更簡便、經濟和高效,同時目標區域覆蓋度更高,便于變異檢測;雖然WES 捕獲探針僅覆蓋人類基因組的1%~1.5%的區域,但是該區域包含了大約85%的已知致病基因變異,可一次性精準檢測人類基因組20000 多個基因中180000 個外顯子的致病變異位點;WES 還解決了GWAS 對罕見突變和結構變異性不敏感的缺點,通過其高效的方法能鑒定新的稀有編碼變體[13-14]。 本文主要以系統性紅斑狼瘡、類風濕性關節炎、多發性硬化病、銀屑病、干燥綜合征、白塞病等常見AIDs 為例,對 WES 在 AIDs 中的應用及研究進展進行綜述。
1 WES 與系統性紅斑狼瘡
系統性紅斑狼瘡(systemic lupus erythematosus,SLE)是一種累及多系統的慢性AID,病程遷延反復[15]。 鄺少松等[16]通過小鼠動物模型證實其主要的病理特征是T 淋巴細胞的失衡,導致了機體免疫紊亂產生大量自身抗體引發炎癥和組織損害,常累及全身多系統或多器官病變,包括心臟、各個關節、皮膚、肺、血管、肝、腎,以及神經系統;在我國人群中的患病率為70/10 萬,全國患病總人數近一百萬,尚缺乏特異的有效治療手段[17]。 GWAS 發現SLE存在遺傳易感性,目前已發現SLE 發病與多個易感基因有關[18]。
SLE 的發病可能是機體免疫失衡所致,近年來,研究人員通過WES 技術已經新鑒定出多個遺傳變異可能通過打破免疫耐受參與SLE 的發生。 李國民等[19]對一例SLE 患兒及其父母進行了WES 和生物信息學分析,后使用Sanger 法驗證突變位點并在其家系其他成員中進行突變分析,篩選得到92 個變異位點,檢測到1 個可能致病基因,即SLC7A7 經典剪切區IVS4+1 G>A 純合變異,此基因突變可以引起巨噬細胞和淋巴細胞中氨基酸代謝異常,從而導致免疫耐受被破壞,出現自身抗體。 李國民等[18]對另一例SLE 患兒及其父母進行WES,發現了105 個變異位點,篩選到可能的致病基因NRASc.38G>A 雜合突變。 而Li 等[2]對7 例SLE 患者及其家屬進行WES,在4 例患者外周靜脈血中發現了雜合的NRASc.38A>G 突變,但其父母均未攜帶,故為新發突變;另 2 例患者檢測到TNFAIP3c.559C>T 和PIK3CDc.3061G>A 雜合突變;NRAS基因編碼蛋白是RAS/MAPK 信號通路中的信號轉導分子之一,該信號通路可將細胞外生長因子信號傳至胞內,對調節細胞周期和細胞的分化、生長、衰老及凋亡起重要作用,因此該基因雜合突變會使RAS/MAPK 信號通路調節異常[17];TNFAIP3 是一個系統的負調控因子,主要誘導免疫耐受,其雜合突變可能打破機體免疫平衡,導致AIDs 的發生[20];PIK3CD編碼的磷脂酰肌醇3-激酶(PI3K)在免疫細胞的激活和功能中起關鍵作用,基因突變引起的PI3K 活性增強會導致AIDs 的發生[21]。 此外,有研究發現相關基因的突變也可以通過調節B 細胞相關信號通路導致AIDs 的發生,Al-mayouf 等[22]對一例罹患 SLE 的六歲女童進行WES,分析確定了致病性TALDO1c.793Cdel 純合子變異和PTENc.518G>C 雜合變異;PTEN可通過B 細胞受體調節正常信號,并在建立外周免疫耐受中發揮作用,其雜合變異可能誘導AIDs 的發生。 Jiang 等[23]對 116 例 SLE 患者 行WES,對97 名沒有慢性病病史的健康老年人行GWAS,研究鑒定出BLK基因6 個罕見突變或新的錯義突變以及BANK1 基因的低頻突變參與了SLE的發病。
以上研究表明,通過WES 可以檢測到與SLE相關的基因變異位點,這些突變位點通過打破自身免疫耐受、影響信號通路等不同機制使得機體免疫失衡而導致SLE 的發生,這促進了我們對于SLE 發病遺傳因素的認識,也可能為SLE 的診斷與治療提供靶標,但尚有一些突變位點缺乏對其相應機制的闡釋,還需進一步研究來證實。
2 WES 與類風濕性關節炎
類風濕性關節炎(rheumatoid arthritis,RA)是一種病因不明的慢性多發性AID[24],其基本病理改變為滑膜炎、血管翳的形成,并逐漸出現關節軟骨和骨的破壞,最終可導致關節畸形和功能障礙,影響了高達1%的世界人口,由遺傳和環境危險因素共同作用導致其發生,其中遺傳率約為65%[25-26]。 最近的GWAS 和相關性研究已經確定了50 多個RA易感位點,GWAS 的模擬數據和直接測序數據表明,高頻、低頻和罕見變異體的混合導致了RA 患病風險的上升[27]。 近年來,大量研究人員已將WES 技術應用到對RA 遺傳因素的研究中。
Okada 等[28]通過 WES,在人類染色體 2q23 區域的PLB1 上發現了一個非同義的c.2263G>C(p.G755R)突變,該突變在顯性遺傳模式的家系成員中與RA 顯著共分離,隨后在1088 例RA 患者和1088名健康對照者(歐洲血統)中進行PLB1 基因的測序分析,證實該突變的存在及其顯著致病風險。 Wang等[29]對51 例不伴有間質性肺疾病(interstitial lung disease,ILD)的 RA 患者和 45 例合并 ILD 的 RA 患者(RA-ILD)進行 WES,研究發現在 RA-ILD 隊列中,攜帶MUC5B變異的患者急性加重或死亡的風險更高,而RA 患者MUC5B突變率為17.6%,因此其變異體的攜帶者狀態可作為RA-ILD 預后降低和病情加重的指標。
綜上,WES 研究發現了新的遺傳變異參與RA的發生,另外發現患者對相應變異體的攜帶與否和疾病的嚴重程度有關,這有助于臨床對患者預后的判斷,但由于WES 在RA 中的應用研究依然較少,更多變異位點尚需進一步的研究確定。
3 WES 與多發性硬化癥
多發性硬化癥(multiple sclerosis, MS)是一種炎癥性、自身免疫性脫髓鞘疾病,以軸突損傷、神經元丟失和中樞神經系統萎縮為特征[30-31]。 越來越多的證據表明,MS 的發生由環境和遺傳因素共同作用導致[32]。 近期的 GWAS 和 Meta 分析發現,人類白細胞抗原DR(HLA-DR)的多態性與歐洲和北美等地區的高加索人群MS 的易感性顯著關聯,而與包括中國人群在內的亞洲人群MS 的發生卻無顯著關聯[30],遺傳異質性表明MS 尚有其他關鍵的遺傳變異未被鑒定,而WES 的應用豐富了我們對MS相關遺傳因素的認識[33]。
Wang 等[30]對來自中國南方的8 例MS 患者和26 名健康對照者外周血單個核細胞中的基因組DNA 進行WES,共發現了41227 個可能的變異,有17 個變異體具有顯著的等位基因頻率,經Sanger 測序證實,3 例 MS 患者具有異質性變異,即位于TRIOBP第 7 外顯子的罕見變異(Chr22 ∶37723520G>T,rs201693690),導致了氨基酸替換,MS 患者的此等位基因頻率明顯高于健康對照組,因此TRIOBP可能是中國南方人群 MS 的新的危險基因,但TRIOBP如何影響免疫或炎癥的機制尚不清楚。Kattimani 等[31]對1 例復發性緩解性多發性硬化癥(relapse-remitting multiple sclerosis, RRMS)女性患者進行 WES 分析,發現miR-8485 攜帶 CA 堿基的移碼純合子缺失;而NRXN1 攜帶外顯子8 的CT 到TC 堿基的非移碼純合替換,絲氨酸替換為亮氨酸;這二者突變改變了鈣穩態和NRXN1/NLGN1 細胞粘附分子結合親和力,miR-8485 可誘導NRXN1 基因沉默,而NRXN1 在調節鈣通道活性和突觸處Ca2+觸發的神經遞質釋放中發揮重要作用,因此突變體miR-8485 對NRXN1 調節表達失調,會導致NRXN1的過度表達,觸發Ca2+內流增加,Ca2+持續升高可能引起氧化應激和細胞死亡,導致神經變性。
上述這些研究結果提示多個遺傳變異參與了MS 的發生,且在不同人群中有所差異,另外還發現這些變異可以通過相應機制影響到患者的具體癥狀,對于研究相應治療方式提供了新的思路,但是仍有變異位點的關鍵作用機制尚且不明,有待進一步的研究。
4 WES 與銀屑病
銀屑病是一種免疫介導的具有多基因遺傳性和復發性的慢性炎癥性皮膚病,以紅細胞斑塊、角質形成、細胞增生和淋巴細胞浸潤為特征,病程較長,易復發,有的病例甚至終生不愈,其發病率約為世界人口的2%,且該病發病以青壯年為主,對人類的身體健康和精神狀況均造成較大影響,常由感染、外傷、藥物等環境因素誘發;GWAS 已鑒定了多個與銀屑病相關的易感位點,但仍未能清楚解釋其發病機理,而WES 技術的應用加深了我們對銀屑病發病機制的理解[34-37]。
Signa 等[38]研究了一個以兒童期起病的紅皮病型銀屑病為特征的有3 對雙胞胎的大家庭,對該家族的5 名成員進行了WES 分析;在CARD14 第4 外顯子中發現新的雜合突變(C.446T>G),此突變可導致以彌漫性紅皮病型銀屑病為特征的不尋常的臨床表型和嚴重的兒童期銀屑病的發生,該變異的存在通過Sanger 測序得到證實;同時該研究發現不同的SNP 可對生物制劑的療效造成影響,這將有助于我們對銀屑病的精準治療。 此外,還有研究發現有些突變與患者治療療效相關,Kuang 等[34]采用WES對22 例尋常型銀屑病患者(11 例甲氨蝶呤治療有效和11 例甲氨蝶呤治療無效)進行了檢測,研究發現3 個錯義突變(SMG6/rs216195 T>C、IMMT/rs1050301 G>A 和UPK1A/rs2285421 T>C)可能與甲氨蝶呤的療效有關,但其具體作用機理還需進一步研究證實。 Carlsson 等[39]研究發現,UPK1A在銀屑病等炎癥性皮膚病中表達顯著升高,但是否由UPK1A/rs2285421 T>C 錯義突變所導致尚不清楚。
以上研究表明,WES 研究新發現了一些參與銀屑病發生的遺傳變異,這些變異不僅僅導致疾病的發生,還有可能與治療效果相關,但尚未闡明具體機制,亟待進一步更深入的研究。
5 WES 與干燥綜合征
干燥綜合征(Sj?gren’s syndrome, SS)是一種以口眼干燥為典型表現的AID,隨病情進展常累及全身各個系統器官[40]。 SS 可分為單獨發病的原發性干燥綜合征(primary Sj?gren’s syndrome, pSS)和伴有其他 AIDs 的繼發性干燥綜合征(secondary Sj?gren’ s syndrome, sSS),其病因復雜,具體發病機制尚不明確,目前普遍認為其發病機制涉及遺傳因素、感染因素和內分泌因素等[41]。 近期的WES 研究,進一步加深了我們對SS 發病機制的理解。
杜蒙[40]應用WES 分析了91 例pSS 患者和152名健康對照者,篩選到25 個罕見變異位點,進一步分析發現,CREBBP和OAS1 是pSS 的易感基因,但其具體作用機制尚不清楚。 王啟迪等[41]對11 例pSS 患者和6 名健康對照者進行WES,篩選獲得25個候選變異位點,后通過Sanger 測序驗證,結果表明,IFIH1c.2115A>C、SIDT1c.1216G>A 可能與 SS 發病相關;IFIH1 編碼胞漿RNA 解旋酶,可在病毒感染過程中提供識別dsRNA 的胞漿受體,通過線粒體抗病毒信號蛋白激活IFN-I 信號通路;SIDT1 是一種多次跨膜蛋白,其膜外區域可結合 dsRNA,并將dsRNA 轉運到細胞內,其作用可能主要與pSS 患者機體的多系統和器官累及有關;因此,推測IFIH1 可能是導致pSS 發生的主效基因,而SIDT1 僅為其中的一個微效基因,然而變異位點的影響、基因在疾病中的具體作用還需功能實驗的證實。 Shen 等[13]分析15 例SS 患者和100 名健康對照者的WES 數據,確定了PRAMEF13、TARBP1、UGT2B28、TRBV56和NAPB為 SS 新的易感基因,并發現UGT2B28/rs72552705 和MSH5/rs2075789 為 SS 風險基因位點;UGT2B28 在調節類固醇激素方面起著重要作用,具有結合某些膽汁酸和雌激素的固有能力,類固醇激素影響免疫細胞功能和炎癥,但其與AIDs 的關系缺乏數據驗證。
WES 的應用不僅鑒定了SS 相關的新的遺傳變異位點,還發現這些遺傳變異位點在疾病的發生發展中起到了不同程度的影響,這為進一步闡釋SS 的發病機制提供了新的思路和研究靶點。
6 WES 與白塞病
白塞病(Beh?et’s disease, BD)是一種累及多系統的由自身免疫介導的炎癥性疾病,以非特異性血管炎為主要病理損害,其典型的臨床特征包括口腔和生殖器潰瘍、葡萄膜炎和皮膚病變等,其發病機制尚不明確,目前普遍認為BD 的病因包括遺傳學、表觀遺傳學、免疫學及環境因素等[42-44]。 近年來WES 研究鑒定發現了多個遺傳變異參與BD 的發生發展。
Dimopoulou 等[42]對一例 BD 患者及其父母進行了WES,研究發現BD 患者ERAP1 基因啟動子區域出現罕見的純合子突變(1-bp 缺失),且BD 患者中ERAP1 的表達水平顯著低于正常對照;此研究也發現WES 可以補充基于DNA 微陣列的GWAS 研究,以調查稀有和非編碼變體。 Ognenovski 等[43]對14 例來自德國的 BD 患者進行了 WES,并通過Sanger 法測序驗證WES 結果,研究發現了與BD 發生相關聯的77 個變異體,其中2 個為新發現的關聯變異體(LIMK2/rs149034313 和NEIL/rs5745908);LIMK2/rs149034313 影響趨化因子和細胞因子信號通路介導的炎癥、T 細胞激活以及血管生成,還參與調控細胞骨架重組,而細胞骨架重塑是BD 中白細胞黏附和浸潤的中心;NEIL1/rs5745908 是一個剪切位點變異,可能引入有害的內含子保留,導致終止密碼子提前出現以及產生非編碼轉錄本,這種有害的效應可能會對氧化應激后的DNA 損傷修復產生不良后果;由于這些基因對BD 中白細胞浸潤和氧化應激的增加是至關重要的,因此所發現的罕見變異關聯可能會為未來的研究提供新的疾病靶點和關鍵途徑。 Shigemura 等[44]調查了一個有BD 病史的日本家庭,對6 例4 代以上BD 患者進行分析,對患者1 和他母親(患者2)單個核細胞提取的基因組DNA 進行WES,研究發現BD 患者均攜帶TNFAIP3雜合錯義突變(chr6_138197226_G>A,C243Y),該突變可能通過抑制NF-κB 的活化而增加人類炎性細胞因子(IL-1β, IL-6 和 TNF-α) 的產生,表明TNFAIP3 突變(chr6_138197226_G>A,C243Y)可能是常染色體顯性BD 的致病基因。
目前,盡管WES 在BD 中的應用較少,但現有WES 研究已經鑒定多個基因變異參與BD 的發生,這些變異通過影響炎性細胞因子的生成、調控細胞骨架系統或增加氧化應激等機制參與了BD 的發生發展,這加深了我們對BD 發生相關遺傳因素的認識,有利于將來對其具體發病機制的闡釋并應用于臨床診斷和治療。
通過以上所述可知,WES 可以檢測到與AIDs相關的基因位點變異(詳見表1),但某些位點的變異不僅僅局限于一種疾病,多種AIDs 在遺傳風險上也共享某些疾病易感基因(詳見表2)。 例如,IFIH1作為介導Ⅰ型干擾素產生的抗病毒解旋酶基因之一,可導致干擾素調節因子和NF-κB 等轉錄因子的激活,從而觸發先天免疫應答,這可能會增加AIDs的風險,經研究證實其遺傳變異與多種AIDs 易感性有關聯:rs1990760 為 SLE、RA、MS 等的共有風險變異,其與血清IL-18、顆粒酶B 水平呈顯著正相關,IL-18 可調節T 細胞分化,打破機體免疫平衡,導致嚴重的組織炎癥和自身免疫;顆粒酶B 通過促進抗原表位的產生和刺激自身反應性T 細胞在獲得性免疫應答和自身抗原的呈遞中起著重要作用,還可以促進 IL-18 的激活,而 IL-18 在 SLE、RA、MS 中均有異常增高。 此外,rs3747517 與 SLE、MS 的發病相關,獨立SNPs 的稀有等位基因rs10930046 變異的攜帶對銀屑病、MS、SLE 有保護作用[45-50]。 而多項研究發現,TNFAIP3 的遺傳變異也與 SLE、RA、MS、銀屑病、SS、BD 等 AIDs 相關,rs2230926 是 SLE 和SS 的共有變異位點,作為NF-κB 途徑的重要負反饋調節因子,rs2230926 異常導致NF-κB 通路的失控,可能參與AIDs 的發生發展[51-52]。 而BD 的易感因子ERAP1 的變異位點rs30187 與銀屑病和MS 的遺傳易感性有關聯,其能夠調控MHC I 類分子的表達,還有助于釋放炎性細胞因子的膜結合受體,通過對抗原的處理和提呈及影響肽修剪參與機體免疫應答[53-54]。MSH5 也被發現在 SLE、MS 及 SS 中發揮作用,其變異使得調節失調可能導致相關疾病易感性, 但具體機制還需更深入研究來證實[13,55-56]。 由此可見AIDs 共享的疾病易感基因可能對疾病的發生發展有相同的作用機制,對這些變異的鑒定將促進我們對AIDs 的進一步認識,為今后的研究提供新的思路。

表1 常見AIDs 中的外顯子變異情況Table 1 Exon variants in common AIDs
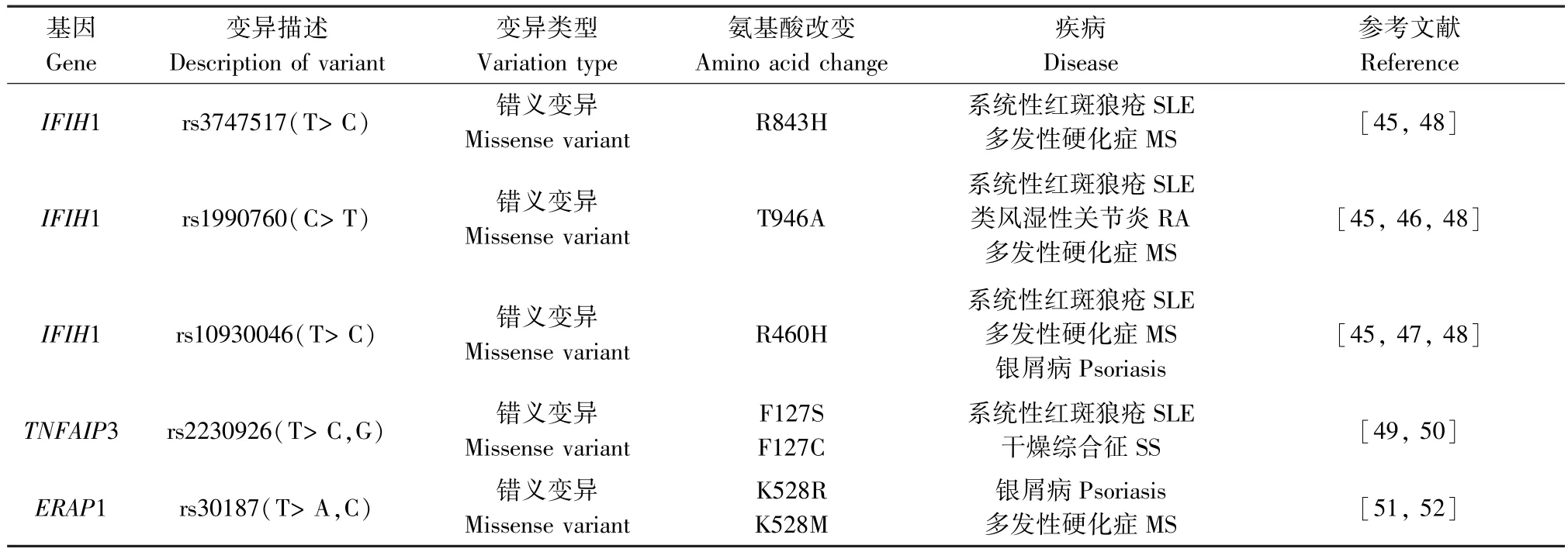
表2 不同AIDs 共性的外顯子變異情況Table 2 Common exon variants involved in different AIDs
7 小結與展望
綜上所述,WES 在多種AIDs 中的使用已新鑒定了多個疾病獨有或共有的遺傳變異,這些變異位點通過各種不同的途徑參與疾病的發生發展,對AIDs 的發生發展提出了新機制。 但是,這些相關聯的遺傳變異如何參與AIDs 發生發展的具體分子機制,尚未完全闡明,有待于進一步的研究。 隨著測序技術的進步及廣泛應用,WES 將來可能有望與蛋白組學、表觀遺傳學等多組學聯合測序,輔以細胞成像技術、原位雜交、免疫組織化學等技術,促進對復雜疾病中基因異常表達調控機制及后續修飾的認識,為AIDs 的預防、早期診斷、監測、治療及臨床預后提供重要的參考。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
