維氏氣單胞菌ΔexsA1減毒株對宿主免疫原性研究
2021-07-16 05:49:54盛強龍徐一軻劉超倫唐燕瓊
熱帶生物學報 2021年2期
盛強龍,徐一軻,劉超倫,李 宏,馬 香,唐燕瓊,劉 柱
(海南大學 生命科學與藥學院,海口 570228)
維氏氣單胞菌(Aeromonas veronii)是一種人魚共患的革蘭陰性病原菌[1],其分布廣泛、致病性較強,給水產養殖帶來巨大的經濟損失[2]。人感染后會出現菌血癥、腦膜炎等癥狀,甚至有死亡的危險[3]。在革蘭氏陰性菌中,III型分泌系統(Type III secretion systems, T3SS)通過分泌胞外蛋白和毒力蛋白介導毒力作用的發揮[4],是細菌向宿主細胞分泌毒力因子的重要途徑,如銅綠假單胞菌、沙門氏菌及福氏志賀菌對宿主的毒害作用皆由T3SS介導[5]。在急性感染期間,銅綠假單胞菌中T3SS特別活躍,感染者的死亡率增加[6-7]。T3SS在細菌病原體中十分保守,其表達由exoenzyme S transcriptional regulator ExsA激活[8]。在副溶血性弧菌(Vibrio parahaemolyticus)中,HlyU正調控exsA基因啟動子,激活T3SS的表達[9];在銅綠假單胞菌中,ExsA可調控細胞外金屬蛋白酶ImpA的表達,后者通過T3SS分泌到宿主體內后,可以裂解巨噬細胞表面蛋白CD44,以抑制巨噬細胞對細菌細胞的吞噬作用[10]。免疫球蛋白M(IgM)是一種普遍存在于脊椎動物中,由效應B淋巴細胞特異性分泌的免疫球蛋白[11-12]。根據Ig重鏈(heavy chain,H)恒定區的差異,硬骨魚類被劃分為IgM、IgD、IgZ/T、IgM和IgZ[13]等類型。在硬骨魚中,IgM較其他類型的免疫球蛋白在含量上占絕對優勢[14-15],檢測IgM成為判斷特異性免疫形成的標志之一。
疫苗主要分為傳統疫苗和基因工程疫苗。傳統疫苗分為滅活疫苗和減毒活疫苗(live attenuated vaccine, LAV),基因工程疫苗分為DNA疫苗、重組亞單位疫苗及活載體疫苗[16]。傳統疫苗制備簡易、成本較低、免疫效果優異,是市場主流。滅活疫苗較減毒活疫苗更加安全,且便于保存;減毒活疫苗接種量更小,但免疫持續時間長,也有一定的開發優勢。常規滅活疫苗雖然激發了顯著的抗體水平,但因為抗體不能有效進入感染細胞殺滅病原菌,無法提供有效保護;而減毒活疫苗進入宿主體內后,到達靶器官的方式與自然感染途徑類似,且持續表達抗原,能有效刺激宿主細胞免疫應答,提高宿主清除感染細胞的能力,是該菌疫苗發展的重要方向[17]。
尼羅羅非魚(Oreochromis niloticus)是重要的經濟養殖魚類,海南是我國主要的羅非魚養殖地,曾發生過以維氏氣單胞菌為主要病原菌的爆發疾病案例[18]。目前針對維氏氣單胞菌的疫苗相對空白,并且現有的嗜水氣單胞菌疫苗和殺鮭氣單胞菌疫苗對維氏氣單胞菌的免疫效果不佳,制備維氏氣單胞菌特異性疫苗迫在眉睫。為了展開針對維氏氣單胞菌疫苗的研發,本實驗構建了維氏氣單胞菌ΔexsA1敲除株,檢測了其毒力表達與免疫原性,為維氏氣單胞菌疫苗的制備奠定實驗基礎。
1 材料與方法
1.1 菌株、質粒維氏氣單胞菌野生株及ΔexsA1敲除株、大腸桿菌WM3064,具體見表1。

表1 實驗菌株及質粒Tab. 1 Bacterial strains and plasmids used for test
1.2 實驗動物尼羅羅非魚(Oreochromis niloticus)魚苗購自海南寶路水產科技有限公司文筆峰養殖基地 ,養殖至實驗用大小,規格為(10±1)g和(50±1)g。
1.3 主要試劑限制性核酸內切酶及其Buffer緩沖液購自NEB公司;T4 DNA Ligase及其Buffer緩沖液購自Monad公司; Rapid Taq Master Mix (2×)、Phanta Max Master Mix、FastPure Gel DNA Extraction Mini Kit、FastPure Gel DNA Extraction Mini Kit等皆購自Vazyme公司;羅非魚免疫球蛋白M(IgM)酶聯免疫分析試劑盒測定購自江蘇酶免實業有限公司。培養基(酵母提取粉、胰蛋白胨、瓊脂糖、瓊脂粉)購自Oxoid公司;二氨基庚二酸(Dap)購自Sigma公司;氯霉素購自北京索萊寶科技有限公司;各種無機鹽等 購自西隴科學股份有限公司;氨芐青霉素(Amp)購自上海源葉生物科技有限公司。
1.4 維氏氣單胞菌ΔexsA1基因敲除株的構建分析exsA1基因及其上下游基因,選擇上下游同源臂并設計其敲除擴增引物exsA1-F1/R1、exsA1-F2/R2和敲除驗證引物exsA1-F0/R0;在敲除擴增引物中添加內切酶BstXI酶切位點,并在該位點中間添加限制性內切酶KpnI和SacI的識別序列,兩同源臂之間添加EcoR I的酶切位點(表2)。提取維氏氣單胞菌基因組,用敲除引物擴增上下游同源臂并純化回收。將上下游同源臂片段用BstXI和EcoR I酶切,pRE112質粒用KpnI和SacI酶切,并連接得到重組質粒,再將連接產物電轉進入大腸桿菌WM3064感受態細胞中復蘇培養,菌落PCR驗證,篩選出陽性菌株。將篩選到的陽性菌株和維氏氣單胞菌野生株分別培養至對數期,再按照(1∶2)、(1∶3)和(3∶1)的比例混合為1 mL,6 000 r·min-1離心3 min后,留30 μL菌液滴入無抗的Dap的LB平板上,30 ℃培養30 h,將重組質粒利用雙親接合的方式轉入維氏氣單胞菌野生株中。取50 μL接合后,將菌液涂布于50 μg·mL-1氨芐青霉素、50 μg·mL-1氯霉素的LB平板上,30 ℃培養過夜。菌落PCR驗證是否是維氏氣單胞菌、是否有重組質粒。將陽性菌株通過8%蔗糖篩選,挑取篩選后的單菌落,使用敲除驗證引物exsA1-F0/R0菌落P CR驗證,對得到的陽性菌株用敲除引物進行PCR擴增,對產物進行了切膠回收后送樣測序。

表2 敲除株構建引物Tab. 2 Nucleotide sequences of primers used for construction of deletion mutants
1.5 細菌生長曲線測定分別挑取維氏氣單胞菌野生株和ΔexsA1敲除株單菌落,置于含有5 mL LB液體培養基的試管中,30 ℃、200 r·min-1過夜培養。紫外分光光度計中測定其OD600值,然后將各菌株菌液稀釋至106CFU·mL-1,分別吸取100 μL稀釋液于96孔板相應位置,無菌新鮮的LB液體培養基作為對照。將96孔板置于多功能酶標儀中,設置程序:溫度恒定30 ℃,持續振板,共測定36 h,每30 min測定1 次OD600值。
1.6 羅非魚的感染將維氏氣單胞菌野生株和ΔexsA1敲除株單菌落接種于5 mL LB液體培養基中,30 ℃、200 r·min-1過夜培養,紫外分光光度計中測定其OD600值后,取OD600=1的菌液,6 000 r·min-1離心3 min后去上清,用無菌的0.9%生理鹽水重懸,定容至2×108CFU·mL-1,即為感染液。感染方法:將羅非魚養殖到(10±1)g后,每組平均分20條。接種感染液,按照10 μL·g-1體質量的注射量對羅非魚進行腹腔注 射攻毒。持續觀察1周,統計各組每天的死亡數。
1.7 血清IgM含量檢測選取長勢和大小相近(50±1)g的羅非魚,平均分為3組,即空白對照組、野生株感染組和ΔexsA1敲除株感染組,并適應性飼養1周。將維氏氣單胞菌野生株和ΔexsA1敲除株單菌落接種于LB液體培養基過夜培養后,配置成上述感染液并稀釋10倍,得2×107CFU·mL-1菌液。對羅非魚進行腹腔注射攻毒實驗,無菌0.9%生理鹽水為對照。9天后再次攻毒同一組羅非魚,分別在攻毒后第2天、第4天和第6天抽取羅非魚血液。將血液室溫靜置30 min,2 000 r·min-1離心20min,吸取上清(羅非魚血清)裝入干凈1.5 mL EP管中,-20 ℃保存備用。采用羅非魚免疫球蛋白M(IgM)酶聯免疫分析試劑盒測定羅非魚血清中IgM含量。試劑盒應用雙抗原夾心法測定標本中IgM水平。固相抗原先與IgM結合,再與HRP標記的抗原結合,形成抗原-抗體-酶標抗原復合物,最后加底物TMB顯色。用酶標儀 在450 nm波長下測定吸光度(OD值),通過標準曲線計算樣品中IgM濃度。
1.8 減毒株對羅非魚的免疫保護率測定將羅非魚養殖到(10±1)g后,分為攻毒對照組、減毒株接種組和接種對照組,每組40條。培養維氏氣單胞菌ΔexsA1敲除株,配置成濃度為2×107CFU·mL-1的注射液。按照10 μL·g-1體質量的注射量對羅非魚進行腹腔注射,接種對照組注射無菌0.9%生理鹽水。9 d后重復上述接種步驟。2次免疫注射結束后,用2×109CFU·mL-1的野生菌株攻毒減毒株接種組和接種對照 組,同等接種量的無菌生理鹽水接種攻毒對照組,記錄各組羅非魚每天的死亡個數,計算累計死亡率。
2 結果與分析
2.1 維氏氣單胞菌ΔexsA1敲除株的構建通過同源雙交換法,設計特異性引物、重組載體構建以及細菌接合和篩選,成功構建維氏氣單胞菌exsA1敲除株,使用敲除驗證引物exsA-F0/R0進行對敲除菌株進行PCR驗證(圖1),由圖1可見,野生菌株使用exsA-F0/R0引物進行克隆時,PCR產物大小為1 100 bp;由于在維菌中存在2個拷貝的exsA, ΔexsA1缺失突變體的PCR產物除了有600 bp的exsA1缺失片段外,還有1 100 bp的野生型克隆產物。將敲除片段切膠回收后,送樣品至生工生物工程(上海)股份有限公司進 行測序,測序結果顯示exsA1已成功缺失。因此,將該菌株命名為維氏氣單胞菌ΔexsA1敲除菌株。
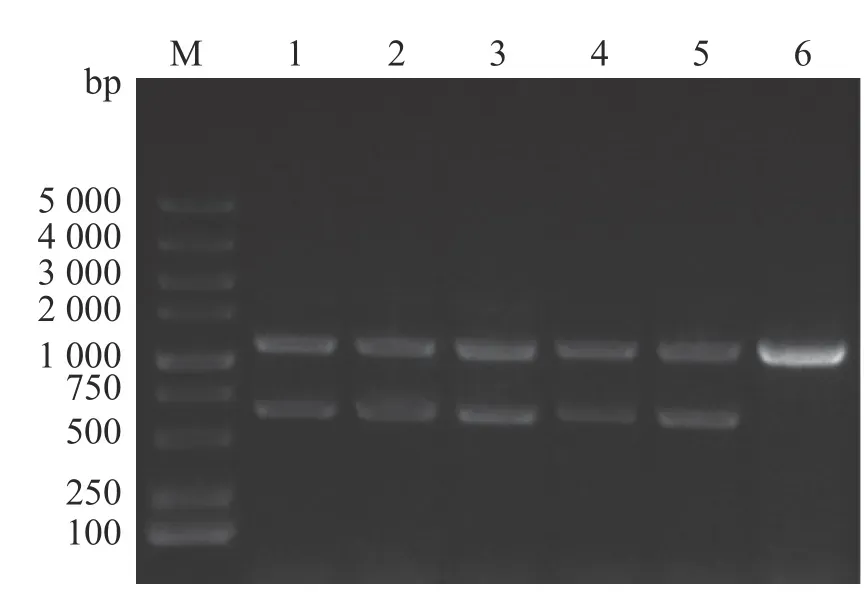
圖1 維氏氣單胞菌ΔexsA1敲除株PCR驗證M:DL 5000 DNA marker;1,2,3,4,5:ΔexsA1敲除株;6:野生株。Fig. 1 PCR verification of Aeromonas veronii ΔexsA1 mutant strainsM: DL 5000 DNA Marker; Lanes 1, 2, 3, 4, 5: ΔexsA1 mutant strains; Lanes 6: Wild strains.
2.2 維氏氣單胞菌野生株和ΔexsA1敲除株生長曲線測定維氏氣單胞菌C4野生株和ΔexsA1敲除株豐富培養下的生長曲線并作圖(圖2),由圖2可見,在LB液體培養基中,維氏氣單胞菌ΔexsA1敲除株與C4野生株在潛伏期時無差異,在對數期和穩定期敲除株的生長量略微低于野生株,但整體相比生長速度不 存在顯著差異。這說明exsA1基因對維氏氣單胞菌的生長沒有明顯影響。
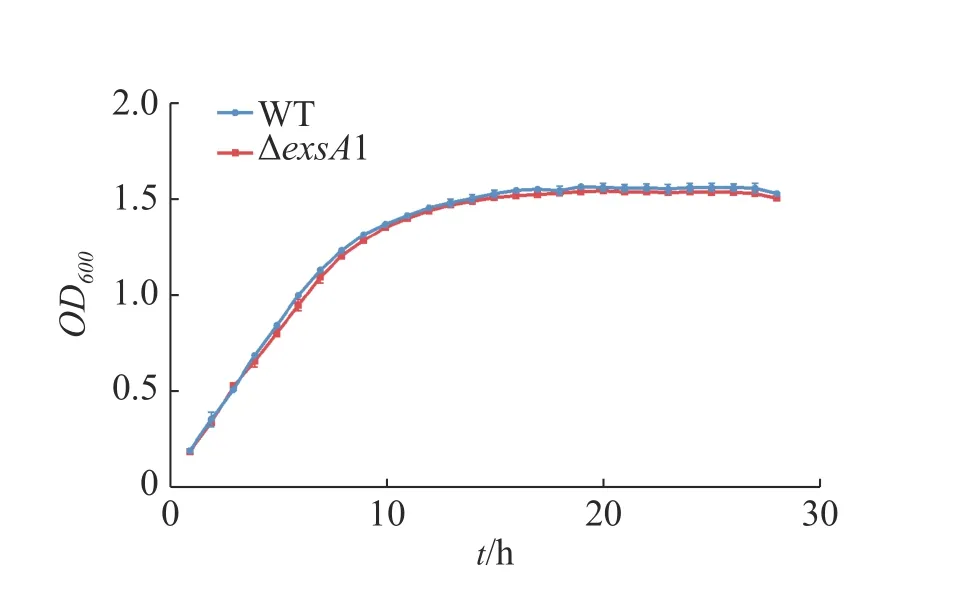
圖2 維氏氣單胞菌野生株和ΔexsA1敲除株生長曲線Fig. 2 Growth curves of Aeromonas veronii wild strains and ΔexsA1 mutant strains
2.3 exsA1基因影響維氏氣單胞菌毒力采用腹腔注射的方法對羅非魚進行攻毒,統計結果見圖3。由圖3可知,ΔexsA1敲除株的感染能力相對于野生株的感染能力更弱。敲除株感染后羅非魚在第1天的死亡率為25%,7天內的死亡率為30%;而野生株感染后羅非魚的第1天時,死亡率就已達到65%,而第3天的死亡率達到85%;空白對照組在觀察期內未出現死亡情況。這說明exsA1基因敲除后,維氏氣單胞菌的毒力減弱了,即為維氏氣單胞菌ΔexsA1減毒株。
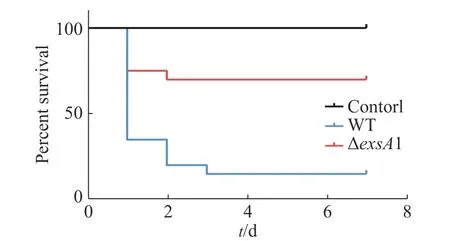
圖3 羅非魚感染維氏氣單胞菌野生株和敲除株后的生存情況Fig. 3 Survival of tilapia infected with Aeromonas veronii wild strains and mutant strains
2.4 羅非魚接種維氏氣單胞菌ΔexsA1減毒株后的IgM檢測采用非致死劑量的維氏氣單胞菌ΔexsA1減毒株,對羅非魚進行腹腔注射,以驗證ΔexsA1減毒株對羅非魚的免疫原性。羅非魚IgM的標準曲線見圖4-A,結果表明,接種ΔexsA1減毒株后,羅非魚體內的IgM得到累積。第4天時,IgM的濃度達到3.81 μg·mL-1,與野生株感染組的IgM累積量沒有顯著差異(圖4-B)。上述結果表明,維氏氣單胞菌ΔexsA1減毒株可以在不致死的情況下,誘導尼羅羅非魚產生與野生型相同水平的免疫反應,具有免疫原性。
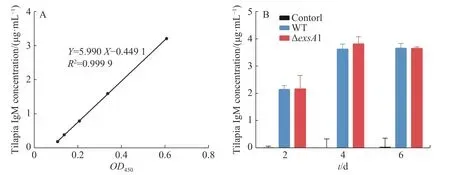
圖4 羅非魚接種exsA1減毒株后的免疫檢測A:羅非魚IgM標準曲線;B:羅非魚血清中IgM濃度。Fig. 4 Immunoassay of tilapia inoculated with the ΔexsA1 attenuated strainsA: IgM standard curve of Tilapia; B: IgM concentration in tilapia serum.
2.5 減毒株對羅非魚的免疫保護率測定第2次接種疫苗的9 d后,用高濃度的維氏氣單胞菌野生型菌株(2×109CFU·mL-1)攻毒,測定其對羅非魚魚苗的免疫保護能力,以驗證維氏氣單胞菌ΔexsA1減毒株對羅非魚的免疫保護效應。結果(圖5)表明,對照組在攻毒后第1天的死亡率高達95%,并且在第2天全部死亡;減毒株接種組第4天的死亡率為30%,整體下降了70%。這說明ΔexsA1減毒株有開發成疫苗的潛力。
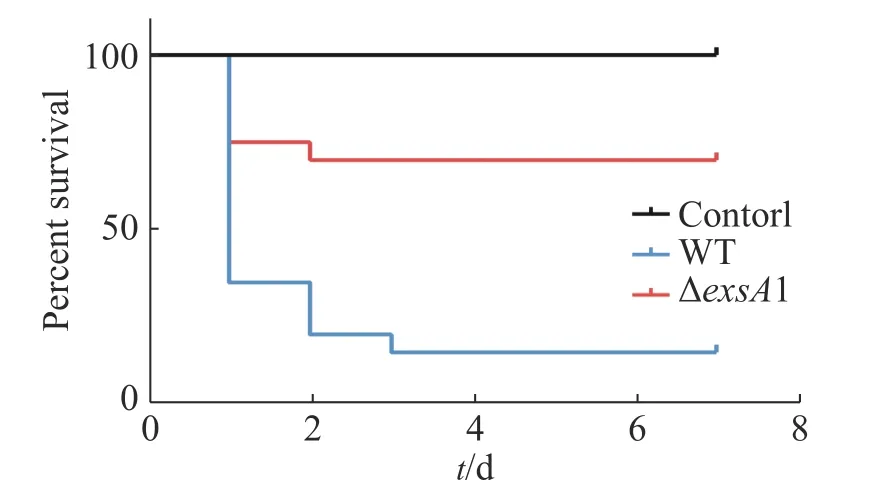
圖5 維氏氣單胞菌exsA1減毒株對羅非魚的免疫保護率Fig. 5 Immune protection rate of Aeromonas veronii ΔexsA1 attenuated strains against tilapia
3 討 論
氣單胞菌(Aeromonas)是革蘭氏陰性菌,是人、魚、獸共患病原菌,其廣泛存在于包括海水在內的各種水域環境甚至土壤環境中[19],給國內漁業生產造成了巨大的經濟損失。維氏氣單胞菌引起淡水漁類的病例在逐年增加[20],但是一直沒有得到足夠的重視。
隨著人們物質生活水平的不斷提高,對優質水產品的需求量快速增長,導致漁業規模迅速擴大,水產品產量得到迅速增加。不少養殖戶為了追求經濟利益,增加水產品的放養密度,導致養殖水質惡化,造成水體的嚴重污染,水產養殖病害大量爆發,對水產養殖產生了巨大的影響[21]。防治水產病害的方法主要是生態防治、藥物防治以及疫苗防治[22]。其中,生態防治對養殖環境要求很高,這會提高水產養殖的成本,我國目前的養殖環境還難以達到。藥物防治多以使用抗生素,初期效果十分顯著,長期使用會導致出現多重耐藥的水產病原菌,使得藥效越來越差,水產病害越來越難以治療。我國獲得商品化生產的魚用疫苗也已有7種,但是我國水產養殖區域廣,而且水產病害種類繁多,這些疫苗對不同病原菌的防治效果不一,水產疫苗的種類是遠遠不夠的,所以針對各種水產病原菌的疫苗研發非常重要,尤其維氏氣單胞菌疫苗的研發更是如此。
本研究用同源重組雙交換的方法,構建維氏氣單胞菌ΔexsA1敲除型菌株,并測定其生長曲線,發現exsA1基因不會影響維氏氣單胞菌的生長。羅非魚腹腔注射感染實驗,證實ΔexsA1敲除株的毒力確實下降。羅非魚攻毒后IgM濃度檢測,證明維氏氣單胞菌exsA1減毒株可以在非致死情況下引發羅非魚的免疫反應,并在體內產生較高濃度的特異性免疫因子IgM的累積。在高濃度野生株感染下,ΔexsA1減毒株對羅非魚的免疫保護率能達到70%,結果表明,ΔexsA1減毒株具有制備成疫苗的潛力。
一直以來,對維氏氣單胞菌感染的水產病害沒有有效的治療手段,面對這種多重耐藥的菌種,制備有效的疫苗刻不容緩。本研究成功構建了具有良好宿主免疫原性的ΔexsA1減毒株,為制備維氏氣單胞菌減毒活疫苗提供了可行的實踐依據。
