維氏氣單胞菌flrB基因敲除株構建及其生物特性鑒定
2021-07-16 05:49:54王祖成唐鴻倩唐燕瓊
熱帶生物學報 2021年2期
王祖成,馬 香,李 宏,王 丹,唐鴻倩,唐燕瓊,劉 柱
(海南大學 生命科學與藥學院,海口 570228)
維氏氣單胞菌(Aeromonas veronii)是一類桿狀的革蘭氏陰性菌,具有極強的適應性,并廣泛存在于水環境中的常見致病菌。維氏氣單胞菌于1983年首次從病患機體中分離出來并命名[1],此后該菌株不斷在魚類、水體環境以及食物中被分離鑒定出來,是一類重要的食源型致病菌[2]。維氏氣單胞菌對人、魚以及陸生哺乳動物均具有極強的致病性。魚類感染維氏氣單胞菌后會使體表充血紅腫甚至潰爛、內臟出血、腹水等癥狀[3-4];人類感染維氏氣單胞菌后會引起嚴重的腸胃感染以及敗血癥,免疫力低下的人群甚至會引發更嚴重的腦膜炎以及尿路感染[5];有案例表明維氏氣單胞菌可感染一些哺乳動物,引發腸胃炎,并導致肝臟受損[6]。近年來,由維氏氣單胞菌引發的感染案例越來越多,這對魚類養殖業以及人類的生命安全構成了嚴重的威脅。因此,解構維氏氣單胞菌感染和致病機理將對維氏氣單胞菌的防治具有重要意義。
雙組分系統廣泛存在于細菌中,通過感知外界信號、信號轉導進而調控細菌內基因的表達,幫助細菌快速適應變化的環境。此外,有研究表明雙組分系統也與細菌致病性相關,熒光假單胞菌可通過RstA/RstB雙組分系統激活外排泵增加耐藥性[7],野油菜黃單胞菌通過RavA/RavR雙組分系統調控鞭毛合成使其更容易侵染宿主[8]。FlrB/FlrC被鑒定為調控細菌鞭毛合成的雙組分系統[9],FlrB作為傳感器激酶具有自磷酸化和傳遞磷酸基團至受體調節器的作用,進而調控下游鞭毛基因的表達,霍亂弧菌鞭毛合成受該雙組分系統調節[10]。FlrB作為感受器激酶是雙組分系統中不可或缺的因子,對鞭毛系統的合成至關重要,并且鞭毛是細菌成分中影響生物膜形成的主要因子[11-12]。研究表明在溶藻弧菌中敲除flrB基因后細菌的運動性和生物膜形成顯著降低[13],在霍亂弧菌中FlrC的磷酸化依賴于同源FlrB,磷酸化的FlrC能提高細菌的運動和定植能力[14]。實驗室前期對維氏氣單胞菌進行全基因組測序分析,發現FlrB/FlrC雙組分系統存在于維氏氣單胞菌中。雖然flrB基因被報道與致病性性狀相關,但在維氏氣單胞菌中的作用機理尚未見報道。因此,本研究旨在構建flrB基因敲除株,揭示維氏氣單胞菌中flrB基因的生物學作用,進一步闡明維氏氣單胞菌的致病機理,為維氏氣單胞菌的治療和防治提供理論基礎。
1 材料與方法
1.1 實驗材料、儀器維氏氣單胞C4菌株、大腸桿菌WM3064、pRE112質粒均為本實驗室自存。基因組提取試劑盒、質粒提取試劑盒、產物純化回收試劑盒、2×PCR MIX購自諾維贊生物科技有限公司;限制性內切為酶(KpnI、XbaI)以及T4連接酶購自NEB公司;氨芐青霉素(水溶,母液濃度50 g·L-1,工作濃度50 μg·mL-1,-20 ℃保存)、氯霉素(乙醇溶,母液濃度25 g·L-1,工作濃度25 μg·mL-1,-20 ℃保存)、二氨基庚二酸(水溶,母液濃度50 g·L-1,工作濃度50 μg·mL-1,-20 ℃保存)、LB培養基(1% NaCL、1%蛋白胨、0.5%酵母提取物)購自索萊寶有限公司。PBS緩沖液(100 mmol·L-1NaCL,8.1 mmol·L-1Na2HPO4,1.5 mmol·L-1KH2PO4,2.7 mmol·L-1KCL,PH 7.4),結晶紫染料、冰醋酸等均為國產分析純,購自海南正恒科技有限公司。Eppendorf Centrifuge 5418 高速臺式離心機(Eppendorf 中國有限公司)、Life ECO-PCR 基因擴增儀(杭州博日科技有限公司)、Bio-Rad MicroPulser 電穿孔儀(美國 Bio-Rad 公司)、HZQ-F100 振蕩培養箱(哈爾濱市東聯生化儀器有限公司)、ME204E 電子天平(梅特勒-托利多儀器上海有限公司)。本研究所用的flrB上下游同源臂擴增引物、flrB敲除驗證引物、pRE112載體驗證引物均合成于生工生物工程(上海)股份有限公司,序列見表1。

表1 基因敲除所用引物序列Tab. 1 Primer sequence for gene knockout
1.2 擴增flrB基因上下游同源臂片段以細菌總DNA為模板,分別用flrB-F1/flrB-R1和flrB-F2/flrBR2 進行PCR擴增目的片段。30 μL PCR體系:2×PCR MIX 15 μL,基因組DNA 1 μL,ddH2O 12 μL,上下游同源臂引物各1 μL。反應程序:95 ℃ 10 min,95 ℃ 30 S,58 ℃ 30 S,72 ℃ 1 min,35個循環,72 ℃ 總延伸10 min 4 ℃停止反應。使用產物純化試劑盒純化回收擴增的DNA片段并保存于4 ℃冰箱待用,同時使 用1%瓊脂糖凝膠電泳檢測擴增條帶大小是否正確。
1.3 pRE112-ΔflrB載體構建使用Overlap PCR連接上下游同源臂。Overlap PCR 體系(50 μL):2×PCR MIX 25 μL,引物flrB-F1 1 μL,引物flrB-R2 1 μL,上游擴增同源臂 1 μL,下游擴增同源臂1 μL,ddH2O 21 μL;PCR程序為:95 ℃ 10 min,95 ℃ 30 S,58 ℃ 30 S,72 ℃ 2 min,35個循環,72 ℃ 總延伸10 min 4 ℃停止反應。回收PCR產物至1.5 mL離心管中,加入30 μL 6×DNA上樣緩沖液混勻并加入到1%瓊脂糖凝膠孔中,120 V恒流電泳25 min。使用凝膠成像儀觀測并切膠回收正確條帶,使用產物純化試劑盒回收凝膠中的DNA片段并檢測濃度。以KpnI和XbaI酶切pRE112質粒和上下游同源臂連接片段,酶切體系50 μL(5 μL Cutsmart mix,KpnI和XbaI各1 μL,pRE112質粒和上下游同源臂連接片段1 500 ng,加水補齊至50 μL),37 ℃酶切4 h。用產物純化試劑盒純化回收酶切后產物。用T4連接酶進行連接酶切后的質粒與片段,連接摩爾比質粒(片段為1∶5,連接體系為:1 μL T4連接酶,1 μL T4連接酶buffer,酶切pRE112質粒100 ng,酶切片段50 ng,加水補齊至10 μL),16 ℃連接1 h。使用電擊轉化法,將連接產物轉化進入大腸桿菌WM3064感受態細胞中,轉化細胞在37 ℃孵育1 h后涂布于氯霉素、二氨 基庚二酸平板上過夜培養;挑取單菌落,使用pRE112引物進行驗證,成功重組載體送測序,作進一步驗證。
1.4 雙親接合及敲除菌株篩選接種野生型維氏氣單胞菌C4和攜帶有pRE112-ΔflrB的大腸桿菌WM3064,分別于30 ℃和37 ℃過夜培養,以2×106CFU· mL-1為起始菌量轉接并培養至OD值為0.4~0.6;將維氏氣單胞菌和大腸桿菌以(1∶1)、(1∶4)、(2∶3)、(3∶2)、(4∶1)的比例混合至1 mL,6 000 r·min-1離心3 min去上清,使用10 μL新鮮培養基重懸菌體并點于DAP平板上,置于30 ℃恒溫培養箱中培養24 h以上。向培養后的接合平板上加1 mL新鮮培養基,用涂布棒刮下菌體于培養基中,取20 μL刮下的菌液稀釋10倍后涂布于氯霉素和氨芐青霉素雙抗平板上,于30 ℃恒溫培養箱中過夜培養,挑取單菌落驗證接合。挑選成功接合的單菌落接種于添加有氨芐青霉素的LB培養基中,30 ℃恒溫培養過夜使其充分發生重組。取過夜培養的菌液依次稀釋10、20、30倍,取200 μL各稀釋梯度的菌液涂布于8%蔗糖和氨芐青霉素平板上,30 ℃恒溫倒置過夜培養。挑選生長出的單菌落用F0/R0引物進行P CR驗證敲除菌株。
1.5 生長曲線測定挑取野生型和敲除flrB基因的單菌落接種過夜活化,以2×106CFU·mL-1起始菌量轉接至添加有氨芐青霉素的200 mL新鮮LB培養基中,150 r·min-1,30 ℃恒溫培養24 h,每1 h取菌檢測OD值。每組樣品設置3個生物學重復,統計測量數據并繪制隨時間變化的生長曲線,觀察其生長情況。
1.6 生物膜形成測定采用96孔板微量法檢測生物膜形成,挑取野生型和敲除flrB基因的單菌落接種于加氨芐青霉素的新鮮LB培養基中過夜活化,以2×106CFU·mL-1為起始菌量轉接至添加有氨芐青霉素的LB培養基中,吸取200 μL菌液于無菌96孔板中置于30 ℃靜置培養36 h。吸出菌液并用PBS緩沖液清洗至澄清,常溫晾干96孔板,加入200 μL 1%結晶紫燃料在常溫下染色10 min,用無菌水清洗板孔至澄清狀態,55 ℃烘干96孔板,加入200 μL 33%冰乙酸于37 ℃下靜置30 min充分溶解附著的結晶紫染 料,酶標儀檢測595 nm處吸光度值。每組樣品設5個生物學重復并統計數據計算差異性。
1.7 數據統計分析所有數據均采用Graphpad 6.02統計軟件進行分析,(ANOVA)顯著性水平設定為P<0.05[11]。
2 結果與分析
2.1 pRE112-ΔflrB載體構建使用維氏氣單胞菌C4基因組DNA為模板擴增上下游同源片段,使用1%瓊脂糖凝膠電泳檢測擴增DNA大小與純度。由圖1-A可知,第1、2泳道分別為flrB基因上游同源臂片段、flrB基因下游同源臂片段,條帶單一明亮且大小正確,適合作為模板進行Overlap PCR連接該上下游片段。將提取的pRE112質粒和Overlap PCR連接的flrB基因上下游同源臂片段進行雙酶切,純化回收酶切片段并連接過夜,通過電轉至大腸桿菌WM3064中并涂布于氯霉素和二氨基庚二酸平板上進行篩選,使用pRE112載體引物進行驗證,以空載pRE112作為陰性對照。從圖1-B可知,泳道1~6條帶大小與陰性對照一致,說明未成功構建,泳道7、8條帶單一且大小位于1 500~2 000 bp之間,大于陰性對照條帶,并且該條帶符合插入目的片段后應具有的大小,說明7、8為成功連接載體,提取質粒測序進一步 驗證。測序結果表明,本實驗成功構建了pRE112-ΔflrB載體。
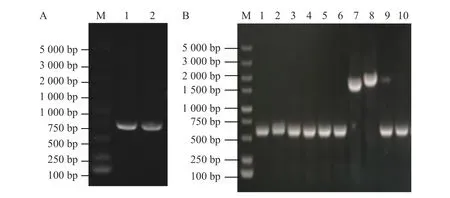
圖1 pRE112-ΔflrB載體構建M:DL5000DNA Maker。A. flrB基因上下游同源臂擴增結果(1:flrB基因上游同源臂擴增;2:flrB基因下游同源臂擴增);B. pRE112-ΔflrB重組載體驗證(1~9:使用載體引物驗證pRE112-ΔflrB的擴增片段;10:以空載體為模板的陰性對照)。Fig. 1 pRE112-ΔflrB vector constructionM: DL5000DNA Marker. A. flrB gene upstream and downstream homology arms amplification results (1: flrB gene upstream homology arm amplification; 2: flrB gene downstream homology arm amplification); B. pRE112-ΔflrB recombinant vector verification (1-9: Vector primers used to verify the amplified fragment of pRE112-ΔflrB; 10: Negative control with empty vector as template pRE112-ΔflrB).
2.2 雙親接合及篩選敲除菌株使用接合的方法將大腸桿菌WM3064中的重組pRE112-ΔflrB載體傳遞到維氏氣單胞菌中,通過抗性平板篩選并進行PCR驗證成功接合菌落。從圖2-A可知,泳道1、2、4、5條帶大小為2 000 bp,與預期大小一致,表明pRE112-ΔflrB載體成功導入野生型維氏氣單胞菌中。為篩選發生同源雙交換的基因敲除菌株,將成功接合的菌落接種培養并涂布于8%蔗糖平板上篩選,使用PCR驗證敲除菌株。從圖2-B可知,泳道3為陰性對照,條帶大小為2 100 bp,泳道1、2 的PCR條帶大小為1 500 bp,減少的片段大小為預期敲除的部分,挑選到2個成功敲除菌株,為驗證敲除的準確性,使用 F0/R0引物擴增并送生工測序,測序結果表明,這2個菌株為成功敲除菌株。
2.3 檢測flrB基因敲除菌株的生長情況通過測定在LB培養基中野生型和flrB基因敲除菌株的生長曲線來研究flrB基因是否會影響維氏氣單胞菌C4的生長。由圖3可知,野生型菌株和敲除flrB菌株生長速率一致,說明敲除flrB后對維氏氣單胞菌的生 長無影響。
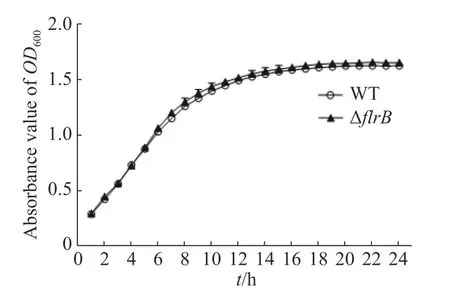
圖3 野生型維氏氣單胞菌和flrB敲除株生長曲線的測定Fig. 3 Determination of the growth curve of wild-type Aeromonas veronii and flrB knockout strain
2.4 檢測flrB基因敲除菌株的生物膜形成定量接種野生型維氏氣單胞菌和flrB敲除菌株并在30 ℃培養箱內培養36 h,吸出菌液使用PBS緩沖液清洗干凈,加入結晶紫染色,洗去多余的染料,用33%乙酸進行溶解脫色后,使用酶標儀檢測OD595的吸光度值。結晶紫染料可以特異性地染色生物膜,由圖4-A可知,敲除flrB后附著在孔壁上的生物膜變少,染色后顏色較淺,而野生型菌株生物膜形成較多,染色較深;使用冰乙酸溶解結晶紫進行定量分析,由圖4-B可知,敲除flrB后生物膜形成顯著降低。說明敲除flrB后生物膜的形成顯著降低,在維氏氣單胞菌C4中flrB基因能促進生物膜的形成。
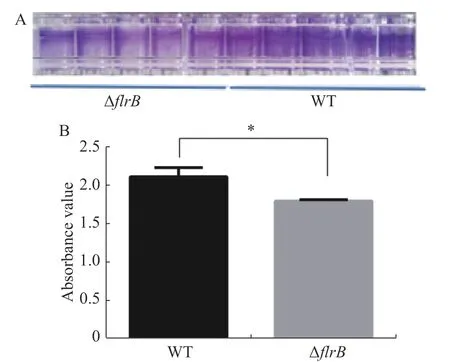
圖4 野生型維氏氣單胞菌和flrB敲除菌株生物膜形成測定A. 96孔板生物膜形成染色結果;B. 使用酶標儀進行測定生物膜形成定量結果。Fig. 4 Determination of the biofilm formation of wild-type Aeromonas veroniis C4 and flrB knockout strainA. 96-well plate biofilm formation staining results;B. Quantitation of biofilm formation on microplate reader.
3 討 論
維氏氣單胞菌是一種廣泛存在的人魚共患致病菌,已對魚類養殖和人類健康造成了巨大的威脅[15],但目前大多數相關研究僅局限于病原菌的分離,未研究其致病機理,這不利于對病原菌的防治。研究維氏氣單胞菌的致病機制將有利于防治病原菌,減少維氏氣單胞菌在養殖業上造成的損失。基因敲除是探究生物表型與基因功能的重要方法,通過敲除候選基因觀察病原菌的致病性有利于研究發掘潛在致病機制。使用同源雙交換進行基因敲除已在革蘭氏陰性菌和革蘭氏陽性菌中廣泛應用,在納塔葡糖酸蠟桿菌、環己氨降解菌、金黃色葡萄球菌中都已通過同源雙交換成功進行基因操作[16-18]。筆者在本研究中采用同源雙交換在維氏氣單胞菌中進行分子操作,研究靶標基因發揮的生物學功能,進而解析維氏氣單胞菌的致病機理。
筆者以維氏氣單胞菌中鑒定出的FlrB/FlrC雙組分系統為靶標,重點研究flrB基因發揮的功能。FlrB是雙組分系統FlrB/FlrC中的傳感器激酶,主要功能為感受信號并自磷酸化將磷酸基團傳遞至同源反應調節器上,從而啟動下游一系列基因的表達,進而影響生物學功能。FlrB/FlrC負責調控細菌鞭毛合成的3級基因的轉錄[19]。flrB的缺失會導致其同源效應調節器不能磷酸化而失去激活下游基因表達的能力,進而影響鞭毛的合成進而影響鞭毛相關的致病表現,如運動性、細胞侵襲、生物膜形成。因此研究flrB基因在維氏氣單胞菌的功能對于解析維氏氣單胞菌的致病機制具有重要的意義。
本研究還鑒定了維氏氣單胞菌中flrB基因對生長和生物膜形成的影響。研究結果表明,敲除flrB基因后并不影響維氏氣單胞菌的生長,flrB基因主要調控細菌鞭毛合成,主要影響細菌的運動以及粘附,因此敲除flrB基因不影響生長,該結果與空腸彎曲菌中鞭毛合成系統缺失但不影響生長特性的研究結果一致[20]。敲除flrB基因后生物膜形成顯著降低,說明在維氏氣單胞菌中flrB具有促進生物膜形成的功能,該結果與溶藻弧菌中flrB基因敲除表型的研究結果一致[13]。本研究可為解析維氏氣單胞菌的致病機制提供新的研究思路與作用靶標。
