基于BSA-SSR 技術的高丹草低氫氰酸性狀目的片段的篩選與鑒定
2021-07-19 07:08:54吳國芳于肖夏于卓楊東升盧倩倩
草業學報 2021年7期
吳國芳,于肖夏,于卓,楊東升,盧倩倩
(內蒙古農業大學農學院,內蒙古呼和浩特010019)
高丹草(Sorghum bicolor×Sorghum sudanense)為高粱(Sorghum bicolor)與蘇丹草(Sorghum sudanense)種間雜交育成的品種(2n=2x=20)[1?2]。其具有生長速度快、分蘗數多、再生性強、抗寒、抗旱、耐倒伏、產草量高、品質好等優點[3?5],可多次青刈飼喂家畜,亦可青貯及調制優質青干草,是我國農區及半農半牧區養殖業發展的重要一年生優良飼草[6]。但幼嫩的高丹草含有一定量的氫氰酸,家畜采食過量后,可引起中毒現象[7?9]。因此,急需培育低氫氰酸含量、飼喂家畜安全的高丹草新品種。
簡單重復序列(simple sequence repeats,SSR),亦稱微衛星DNA 標記,其均勻分布在植物基因組中[10]。SSR標記呈共顯性遺傳,可鑒別純合子和雜合子、多態性好、對DNA 質量要求不高且用量少,目前已成功應用于作物品種鑒定、遺傳圖譜構建、重要性狀QTL 定位及標記輔助育種等研究[11?14]。群體分離分析法(bulked segregation analysis,BSA)即DNA 混池法,由Michelmore 等[15]1991 年首次提出。該方法是利用具有一定表型差異的親本雜交產生后代分離群體單株,建立具有目標性狀的兩個極端DNA 基因池,進行相關性狀的分子標記篩選,若在2 個極端DNA 基因池之間存在一定的多態性分子標記差異,則表明該標記與目標性狀基因呈連鎖遺傳[16]。目前,國內外學者將BSA 法與SSR 分子標記相結合,已成功應用于小麥(Triticum aestivum)根部線蟲抗性基因片段[17]及水稻(Oryza sativa)葉枯病[18]、甘蔗(Saccharum officinarum)褐銹病[19]、高粱粒重[20]等性狀的基因片段篩選,而在高丹草低氫氰酸含量性狀相關基因片段的篩選尚未見有研究報道。
本課題組前期以氫氰酸含量較高的散穗高粱(scattered ear sorghum)作母本,氫氰酸含量較低的紅殼蘇丹草(red hull sudangrass)作父本,通過人工去雄雜交成功獲得高丹草F1代植株,進而將F1代套袋自交獲得F2代分離群體[21?22]。本試驗以高丹草F2分離單株群體及其親本為材料,通過測定氫氰酸含量篩選出高氫氰酸含量植株和低氫氰酸含量植株,構建DNA 基因池,利用BSA?SSR 法篩選與高丹草低氫氰酸含量性狀相關的SSR?DNA 目的片段,并對其進行回收、純化和測序,以期鑒定出高丹草低氫氰酸含量性狀的相關基因片段,為下一步低氫氰酸性狀主效QTL 精細定位、基因圖位克隆及其功能解析奠定基礎。
1 材料與方法
1.1 供試材料及種植
材料為高丹草F2分離群體及其親本散穗高粱和紅殼蘇丹草。2020 年5 月種植在內蒙古農業大學農場,行距為50 cm,株距為25 cm。生長期內根據生長狀況及時施肥、灌水,保證植株對養分和水分的需求,并注意防除田間雜草。
1.2 研究方法
1.2.1 氫氰酸含量測定 在高丹草拔節期,株高約為100 cm 時,田間剪取F2代500 個分離單株及親本的新鮮莖葉,按單株用微型粉碎器粉碎混勻,每個樣品分別稱取1.0 g,放入裝有8 滴甲基橙、10 mL 硝酸鋅和270 mL 蒸餾水的500 mL 蒸餾瓶中加熱蒸餾。用裝有10 mL 1.0% NaOH 吸收液的100 mL 容量瓶收集餾出液,當容量瓶中的餾出液為100 mL 時,蒸餾結束。吸取10 mL 餾出液放入25 mL 容量瓶中,加入5 mL 磷酸鹽緩沖液和4 滴1.0%的氯胺T,充分搖勻3 min,然后加入5 mL 異煙酸?吡唑啉酮比色酸(5∶1 混合物)進行顯色處理[23],用日本島津公司產紫外分光光度計UV-1800 在636.5 nm 處比色測定樣液的吸光度值y,每個樣品重復測定3 次。以CN?濃度(μg·mL?1)為橫坐標,吸光值為縱坐標繪制標準曲線,得到回歸方程y=0.1078x?0.0019(R2=0.9999)。方程中y為吸光度,x為 CN?的質量濃度,計量單位換算成 mg·kg?1FW[7]。
1.2.2 DNA 提取與DNA?BSA 混池建立 本試驗用植物基因組DNA 提取試劑盒提取F2低氫氰酸含量(<15 mg·kg?1FW,簡稱低氰)和高氫氰酸含量(>300 mg·kg?1FW,簡稱高氰)的極端單株各10 株及母本散穗高粱和父本紅殼蘇丹草的DNA。從極端株中各吸取5 μL 的DNA 進行等量混合,形成低氰基因池和高氰基因池。
1.2.3 SSR 引物設計與PCR 擴增 試驗所用70 對SSR 引物是根據已公布的高粱基因組EST 序列,借助SSR Hunter 軟件查找SSR 位點,利用Primer 5 軟件設計出SSR 引物序列,委托南京金瑞斯生物科技有限公司合成。
PCR 反應體系總體積為 20 μL。其中含 Mg2+的 10×buffer 2.0 μL,2.5 mmol·L?1的 dNTP 1.5 μL,5 U·μL?1的Taq-poLymerase 0.2 μL,10 ng·μL?1上下游引物各 1 μL,模板 DNA 2 μL,ddH2O 補足 20 μL。
PCR 擴增程序:循環前 94 ℃預變性 5 min;94 ℃變性 30 s,56 ℃退火 30 s,72 ℃延伸 1 min,38 個循環。循環后 72 ℃延伸 10 min,4 ℃保存。PCR 反應結束后,加入 5.0 μL 變性劑,95 ℃變性 5 min,迅速取出在冰上冷卻,置于?20 ℃條件下保存備用。
1.2.4 電泳及膠板銀染 將上述PCR 變性后的樣品取5 μL,用6%聚丙烯酰胺凝膠電泳檢測PCR 產物的質量。電泳緩沖液為1×TBE,恒定功率70 W,電泳時長1.2 h。電泳后對膠板進行銀染,參照李建武等[24]的染色方法。
1.2.5 低氰DNA 目的片段的回收與純化 以母本散穗高粱SSR?DNA 片段為高氰參比,以紅殼蘇丹草SSR?DNA 片段為低氰參比,用100 bp 的LadderDNA 作Marker,在電泳膠板上找出低氫氰酸含量的DNA 片段進行回收。在聚丙烯酰胺凝膠電泳膠板上分別用無菌手術刀將不同SSR 引物擴增出的單一低氰目的片段條帶切下,置1.5 mL 離心管中,加入0.2 mL ddH2O,用槍頭將膠塊充分碾碎,加蓋后將離心管置入100 ℃沸水中加熱10 min,釋放凝膠內的DNA。冷卻2 min 后,4 ℃下12000 r·min?1離心5 min,吸取上清液轉入新的離心管中。
1)低氰目的片段二次PCR 擴增及電泳檢測。將以上得到的上清液用原PCR 體系和程序進行再次擴增,產物用1.2%瓊脂糖凝膠電泳檢測(電壓120 V,電泳25 min)。若電泳結果顯示清晰的單一條帶,且分子量大小與聚丙烯酰胺凝膠中的低氰條帶相對應,則表明回收質量較好;若瓊脂糖凝膠電泳檢測結果顯示的條帶在2 條以上,則表明目的片段回收質量較差,需進一步純化。
2)目的片段純化。用索萊寶公司生產的PCR 產物純化試劑盒,依據說明書對2 條以上低氰目的片段的DNA進行純化。
3)目的片段命名。采用目的片段(target fragment)的英文縮寫“TF”+“片段數目”,如TF1。
1.2.6 純化DNA 檢測 吸取純化得到的DNA 片段溶液5 μL,用1.2%瓊脂糖凝膠電泳檢測,若低氰DNA 目的片段呈清晰單一條帶,則達到測序要求。
1.2.7 低氰DNA 目的片段的測序及序列比對 委托上海生物工程股份有限公司對篩選出的高丹草低氰DNA 目的片段進行測序。將測序獲得的低氰目的片段序列,與GenBank(http//:www. ncbi. nLm. nih. gov/BLAST/)和高粱基因組數據庫V3.1.1(http://www.gramene.org/Sorghum_bicolor/Location)進行同源性比對分析。
2 結果與分析
2.1 基因組DNA 的純度檢測
將提取后的各供試材料基因組DNA 用紫外分光光度計在不同吸光度(OD)下檢測結果顯示,OD260/OD280均在1.80~2.00 范圍內,說明所提取的DNA 濃度適宜。進而用1.2%瓊脂糖凝膠電泳檢測結果顯示,各樣品電泳條帶清晰穩定,無拖尾降解現象(圖1),表明DNA 純度高,完全能滿足SSR 分子標記試驗的要求。

圖1 高丹草F2部分單株基因組DNA 電泳檢測Fig.1 Sorghum-sudangrass hybrids of F2 plants genomic DNA electrophoresis results
2.2 高丹草F2 BSA 混池及其親本DNA 擴增
用70 對SSR 引物對F2的高氰和低氰植株基因池及親本DNA 進行PCR 擴增及電泳檢測結果表明,70 對引物中有9 對引物(表1)可以區分高氰、低氰性狀(在高氰中無SSR 條帶,在低氰中有明顯的SSR 條帶)。9 對引物中,篩選到與高丹草低氰性狀有關的特異性SSR 條帶共26 個,其片段大小在100~750 bp(表2,圖2)。從圖2 還可以看出,個別條帶帶型較寬,例如TF3-240、TF8-100 等,這可能是相似的序列遷移造成的,需要進行瓊脂糖凝膠電泳檢測后進一步純化。

圖2 高丹草BSA 混池及親本部分SSR 低氰目的片段的篩選Fig.2 Screening of low cyanide target fragments of the BSA gene pools of sorghum-sudangrass hybrid and parents

表1 9 對SSR 適宜引物的堿基序列Table 1 The nucleotide sequences of 9 pairs of SSR primers combinations

表2 高丹草SSR 低氰目的片段篩選Table 2 Screening results of low cyanide target fragments in sorghum-sudangrass hybrid
2.3 二次PCR 擴增檢測與純化
將26 個低氰SSR 目的片段切膠回收后作為模板,按原PCR 程序再次擴增,擴增產物用1.2%瓊脂糖凝膠電泳 檢 測 發 現 ,共 有 14 個 低 氰 DNA 片 段 目 的 片 段(TF2、TF4~TF6、TF8、TF11、TF12、TF14、TF15、TF17~TF20 和TF26)均呈現出一條清晰的DNA 條帶(圖3),表明這14 個片段不需要純化;而有12 個低氰片段(TF1、TF3、TF7、TF9、TF10、TF13、TF16、TF21~TF25)出現了 2 條以上條帶,說明這 12 個目的片段回收質量較差,需從SDS?PAGE 中進行純化。
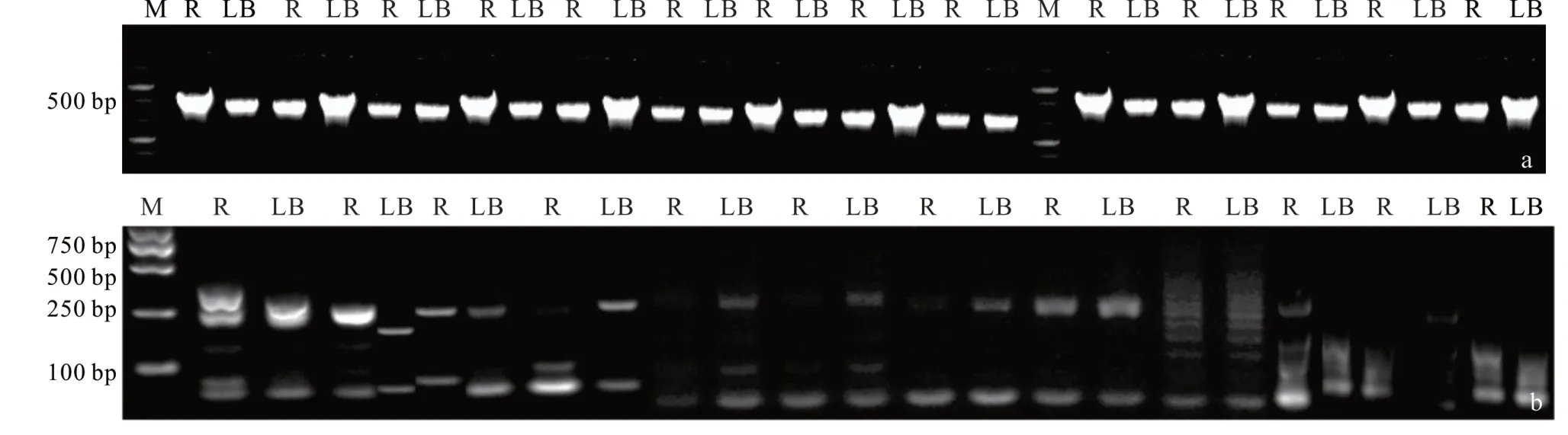
圖3 26 個低氰目的片段二次擴增檢測Fig.3 Secondary amplification detection of 26 low cyanide target fragments
經純化后的12 個低氰DNA 目的片段均呈現單一清晰的條帶,其分子量大小在250 bp 左右,與前面聚丙烯酰胺凝膠中的條帶相吻合(150~300 bp),表明DNA 片段純度高,純化效果理想,符合測序要求(圖4)。
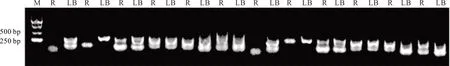
圖4 12 個低氰目的片段純化檢測Fig.4 Purification and detection of 12 low cyanide target fragments
2.4 目的片段測序分析
從表3、圖5 和圖6 可知,26 個低氰目的片段測序長度在56~672 bp 范圍內,這與Marker 估計片段大小100~750 bp 有一定差異,如原片段 TF1、TF2、TF8 和 TF16 的大小分別為 300、280、100 和 550 bp,而實際測序結果分別為282、271、101 和586 bp,表明目的片段對應的Marker 遷移的位置與堿基序列的大小存在一定差異。

圖6 高丹草部分低氰目的片段DNA 堿基序列Fig.6 DNA base sequence of some low cyanide target fragments in sorghum-sudangrass hybrid

表3 26 個低氰目的片段DNA 測序Table 3 DNA sequencing results of 26 low cyanide target fragments
低氰片段TF1、TF2、TF8 和TF16 的序列峰圖中,用綠色、紅色、黑色和藍色4 種顏色的峰表示4 種堿基(A、T、G、C)的信號強度;圖中波峰與波谷清晰,峰與峰之間的距離均勻;未出現重疊峰與堿基缺失,在底部沒有雜峰干擾,表明測序結果是理想的(圖5)。
2.5 同源性分析
根據DNA 測序結果,利用BLAST(http//:www.ncbi.nLm.nih.gov/BLAST/)網站序列數據庫和高粱基因組數據庫(http://www.gramene.org/Sorghum_bicolor/Location)對篩選出的26 個低氰目的片段進行同源性比對發現,14 個序列具有同源性,目的片段 TF1、TF4、TF5、TF12 和 TF26 與過氧化物酶受體 XM_002758639.5、K0J259、AF310249.1、A0A2K6B7D4 和 A0A2K6RRD2 的同源性在 88.2%~100.0% 范圍內;片段 TF17 與干腐菌未表征蛋白F8QJV4 的同源性為96.4%,TF18 與未培養的真菌克隆基因MT703519.1 的同源性為91.6%;片段TF21 與鼠傷寒沙門氏菌氨肽酶Q8ZQ76 的同源性為84.6%;片段TF19 和TF20 與黃孢原毛單胞菌18S RNA基因的同源性為83.2%;片段TF23 與磷脂酰絲氨酸脫羧酶原酶2 基因A0A072PQP8 的同源性為81.3%;片段TF25 與甘藍型油菜蛋白A0A078FW90 的同源性為81.0%;而與高粱基因組序列相關的只有TF8 和TF16 2 個片段,其中TF8 與葉綠體磷酸核糖激酶XM_021458168.1 的同源性高達95.4%,片段TF16 與高粱克隆BAC 88M4 基因AY661656.1 的同源性高達到97.0%,這表明這2 個基因與高丹草的低氰性狀相關,推測其是調控低氫氰酸含量性狀的基因。其余的 TF2、TF3、TF6、TF7、TF9~TF11、TF13~TF15、TF22 和 TF24 這 12 個片段的堿基序列沒有同源性(表4),說明前人尚未研究與這些DNA 目的片段相關的基因。

表4 高丹草低氰目的片段同源性比對Table 4 Identity of comparison of low cyanide target fragments in sorghum-sudangrass hybrid
2.6 低氰目的片段TF8 和TF16 的堿基序列比對
用DNAMAN 8.0 軟件,將低氰父本紅殼蘇丹草及低氰F2共有目的片段TF8 和TF16 與同源性高的高粱基因AY661656.1 和XM_021458168.1 進行了序列比對和反向翻譯,發現低氰父本紅殼蘇丹草及F2的低氰片段堿基序列完全一致,而與高粱基因的堿基比對有一定的變異。進而將低氰序列代入BLASTn,得到片段TF8 的登錄號為lcl|Query_51852,片段TF16 的登錄號為lcl|Query_10405。前者與高粱葉綠體磷酸核糖激酶XM_021458168.1 基因相比,發生了3 次堿基替換(C-A、A-T 和C-G)和1 個C 堿基插入,相應的丙氨酸和甘氨酸替換成半胱氨酸,蘇氨酸替換成丙氨酸,而缺失了甘氨酸;后者與高粱克隆BAC 88M4 基因AY661656.1 相比,僅有一個A 堿基缺失,導致丙氨酸缺失(圖7 和表5)。
表5 低氰片段TF8 和TF16 氨基酸差異Table 5 The difference amino acids of low cyanide fragments TF8 and TF16

圖 5 低氰目的片段 TF1、TF2、TF8、TF16 的序列峰Fig.5 Sequence peaks of some low hydrocyanic acid target fragments TF1,TF2,TF8,TF16

圖7 低氰目的片段TF8 和TF16 堿基序列特征Fig.7 Base sequence characteristics of TF8 and TF16 of low cyanide target fragments
綜上,本研究首次找到了兩個(XM_021458168.1 和AY661656.1)與高粱相關的低氰性狀片段基因,但這兩個基因的功能注釋中并沒有關于氫氰酸性狀的描述,本試驗補充了這兩個基因具有調控低氰性狀的作用,從而為下一步高丹草低氰性狀主效QTL 精細定位、圖位克隆及標記輔助等研究奠定了基礎。
3 討論
本試驗將獲得的26 個低氰SSR?PCR 產物片段進行回收、純化、測序后,與BLASTn 比對發現,其在GenBank 數據庫中有14 個目的片段與過氧化物酶、真菌、氨肽酶、葉綠體磷酸核糖激酶和BAC 88M4 基因的同源性在81.0%~100.0%范圍內,其中葉綠體磷酸核糖激酶和BAC 88M4 分別是高粱基因XM_021458168.1 和AY661656.1 的功能注釋,它們正好與低氰片段TF8 和TF16 相對應,其同源性分別高達95.4%和97.0%。進一步通過比對分析堿基序列發現,片段TF8 在XM_021458168.1 基因的218~276 bp 之間的丙氨酸和甘氨酸替換為半胱氨酸,蘇氨酸替換為丙氨酸,甘氨酸缺失;片段TF16 在AY661656.1 基因的127691~127632 bp 之間僅有1個丙氨酸缺失。據此推測TF8 和TF16 這2 個低氰DNA 片段由于半胱氨酸、丙氨酸和甘氨酸發生替換和缺失,產生了調控低氫氰酸含量性狀的功能。研究表明,XM_021458168.1 編碼高粱葉綠體磷酸核糖激酶,而磷酸核糖激酶(phosphoribulokinase,PRK)是(calvin-benson-bassham)CBB 循環的必需酶[25?28],表明 XM_021458168.1 可能調控氫氰酸的合成,其調控機制有待深入研究。其余12 個DNA 目的片段在現有GenBank 數據庫中尚未找到同源性數據,這與劉改艷等[29]在研究烏鱧性別時發現8 個可以區分烏鱧性別的DNA 目的片段,卻在GenBank 中沒有找到相似的同源性數據,表明現有的GenBank 數據庫還需要研究者們不斷完善。
BSA 法在植物重要性狀目的基因片段篩選中,已有一些成功應用的研究報道[30]。本試驗采用BSA 與SSR結合的方法篩選出26 個與低氰相關的DNA 目的片段,從中首次鑒定出TF8 和TF16 這2 個DNA 目的片段基因具有調控低氫氰酸含量性狀的作用,再次證明BSA 法是篩選和鑒定植物目標性狀基因片段的一種有效方法,它不僅可以克服沒有創建近等基因系的限制[31],而且還具有省時和減少工作量的優點。
本試驗在對高丹草低氰SSR 目的片段切膠回收過程中發現,按原引物及PCR 體系、程序進行二次PCR 擴增及產物用瓊脂糖凝膠電泳檢測時,出現了目的片段條帶2 條以上的現象(圖3),且主要出現在原目的條帶相鄰位置。通過與原樣品聚丙烯酰胺凝膠的條帶逐一對比發現,并未產生的新的SSR 條帶,表明在瓊脂糖凝膠電泳中出現了DNA 條帶共遷移現象[32?33]。為了消除DNA 測序帶來的誤差,對出現共遷移現象的低氰DNA 片段條帶進行了純化,保證了DNA 片段測序的準確性(圖4)。該經驗可為今后類似試驗研究提供借鑒。
4 結論
利用BSA?SSR 技術篩選出26 個高丹草低氰SSR 目的片段,經測序分析明確了這26 個低氰DNA 片段的堿基序列。序列比對發現,TF8 和TF16 這2 個低氰目的片段分別與高粱葉綠體磷酸核糖激酶XM_021458168.1 和高粱克隆BAC 88M4 AY661656.1 基因同源性高達95.4%和97.0%,片段大小分別為101 和586 bp。但在DNA堿基上有一定變異,TF8 片段發生了3 次堿基替換和1 次插入(C-A、A-T、C-G、C 堿基插入),導致丙氨酸和甘氨酸替換成半胱氨酸,蘇氨酸替換成丙氨酸,甘氨酸缺失;TF16 片段有1 個A 堿基缺失,導致丙氨酸缺失。據此推斷TF8 和TF16 低氰片段有調控低氫氰酸含量性狀的功能。
