小麥Brock抗白粉菌侵染早期防御應答基因分析
2021-07-28 04:51:46張露露卞云迪劉曉穎范寶莉王振英
天津師范大學學報(自然科學版) 2021年3期
張露露,卞云迪,張 馳,劉曉穎,范寶莉,王振英
(1.天津師范大學生命科學學院,天津300387;2.天津師范大學 天津市動植物抗性重點實驗室,天津300387)
由禾本科布氏白粉菌(Blumeria graminisf.sp.tritici(Bgt))引起的小麥白粉病常發生于多雨、潮濕地區,病菌多侵染小麥的葉片、葉鞘等部位,影響小麥生長發育,嚴重時可導致減產13%~34%[1].小麥受到病原體侵害后,會立即啟動抗病基因進行防御反應.在流行病害初期,植物抗病基因誘導產生的抗性可以有效減緩病害的發展進程,隨著病原菌最適生長溫度、濕度等環境條件的改變,病原菌增殖受到抑制,最終避免病害發生.因此,分析小麥在白粉菌侵染早期的基因表達模式對于探究小麥抗病機理具有重要意義.
Brock是一個強抗白粉病小麥品種,李根橋等[2]研究發現,該品種攜帶Pm2基因.Pm2是位于小麥5DS染色體上的一個編碼NBS-LRR結構的抗病蛋白基因[3].本課題組前期分別在抗病親本Brock、感病親本京411及其抗病近等基因系BJ-1中擴增Pm2基因的cDNA序列,最終只在Brock中擴增到該基因cDNA序列,而京411和BJ-1中沒有擴增到該基因,因此推測抗病近等基因系BJ-1中的抗性不是來自Pm2,Brock作為BJ-1的抗源供體,至少還含有一個Pm2以外的抗病基因[4].Carver等[5]利用白粉菌孢子侵染大麥時發現,侵染20 s內就可檢測到白粉菌孢子所釋放的細胞外基質(ECM).Klement等[6]研究發現,假單胞菌能夠在短時間內誘導煙草細胞發生超敏反應.Both等[7]發現,小麥白粉菌病原體侵染30 min到5 d內小麥中會出現一些與初級代謝產物相關的酶.這些結果均表明,病原菌與宿主細胞的互作發生在接種后的1 h內.本課題組前期的研究工作中,比較了抗病品種Brock、感病品種京411以及它們的抗病近等基因系BJ-1在白粉菌脅迫下的早期應答模式,發現抗病品種Brock和抗病近等基因系BJ-1均在2 hpi(hours post inoculation)對白粉菌做出應答反應[8-9],說明小麥在白粉菌侵染早期就已開始啟動防御反應.在小麥與病原菌互作應答的研究中,雖已鑒定出許多差異表達基因(differentially expressed gene,DEG),但對于早期防御應答中的差異表達基因以及由這些基因構建的不同抗病信號途徑還需要進一步探究.
本研究利用高通量RNA-seq技術鑒定了白粉菌脅迫下抗病品種小麥Brock中2~8 hpi的差異表達基因,并分析了抗病相關基因的表達模式.這將為深入探究白粉菌侵染早期的小麥抗白粉病模式提供實驗基礎.
1 材料和方法
1.1 材料
抗白粉病小麥品種Brock,英國引進.白粉菌生理小種E09,由中國農業科學院植物保護研究所提供.
實驗所用引物如表1所示,由蘇州金唯智生物科技有限公司合成.
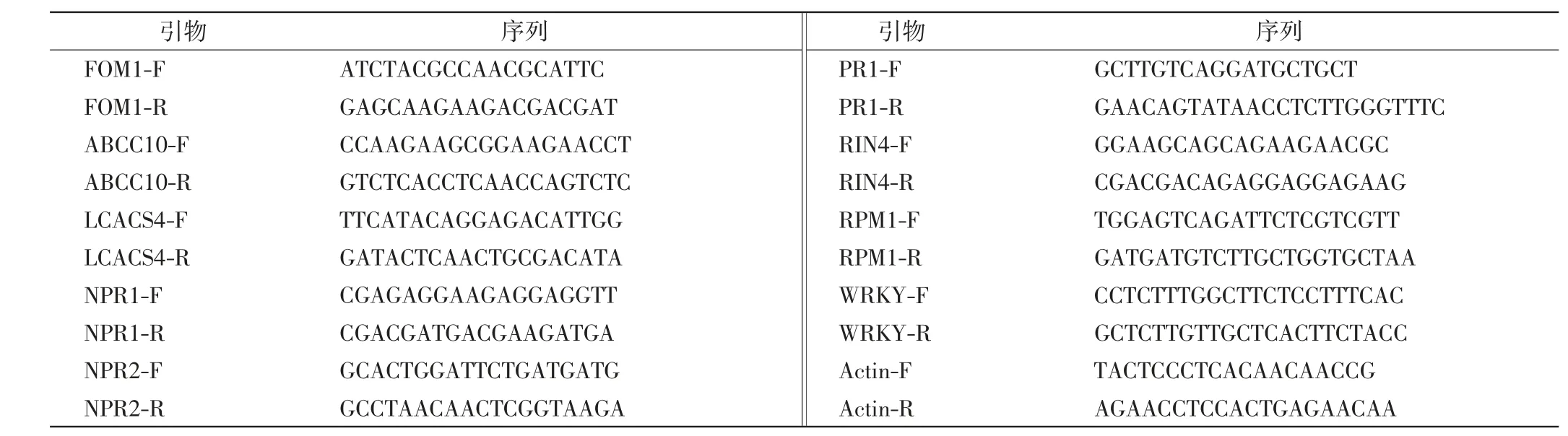
表1 引物序列Tab.1 Primer sequences used in this study
1.2 方法
1.2.1 轉錄組測序樣品的制備及文庫構建
待小麥生長至一心一葉期接種白粉菌E09.分別在0、2、4和8 hpi,剪下相同部位的小麥葉片,分別提取葉片總RNA.提取步驟按照植物RNAprep Pure Plant Kit(天根生化科技(北京)有限公司)說明書進行.將2、4和8 hpi的小麥葉片總RNA樣品等量混合作為實驗組文庫(BMI),以0 hpi的葉片總RNA作為對照組文庫(BCI),2組樣品處理各設置2個重復,共4個文庫.RNA-seq文庫的構建和轉錄組分析工作由諾禾致源生物技術有限公司完成.所有RNA-seq數據已上傳至NIH Short Read Archive數據庫(http://www.ncbi.nlm.nih.gov/sra),登錄號為PRJNA480377.
1.2.2 數據處理和差異表達分析
從Ensembl(ftp://ftp.ensemblgenomes.org/pub/plants/release-35/fasta/triticum_aestivum/dna/)下載小麥參考基因組信息.使用DESeq R軟件(1.18.0)對2個處理(每個處理設置2個生物學重復)進行差異表達分析,并設置經校正后的P值<0.05時為差異表達基因.
1.2.3 差異表達基因的GO和KEGG富集分析
利用GOseq R package(1.18.0)對差異表達基因進行GO(gene ontology)分類,并對基因長度偏差進行校正.利用KOBAS software在KEGG(Kyoto encyclopedia of genes and genomes)數據庫(http://www.genome.jp/kegg/)中對差異表達基因進行KEGG通路富集分析.GO和KEGG富集分析中,經校正后的P值<0.05時被認為顯著富集.
1.2.4 qRT-PCR驗證
Brock幼苗長至一葉一心期時,按照1.2.1方法接種白粉菌并分別對0、2、4和8 hpi的葉片進行取材.按照1.2.1中的方法制備小麥RNA,并利用M-MLV Reverse Transcriptase(美國Promega公司)反轉錄為cDNA,同時根據cDNA非保守區域序列設計引物,如表1所示.根據Fast Start Universal SYBR Green Master mix(美國Roche公司)說明配制PCR反應體系:SYBR Green Mix 10μL,cDNA 1μL,F 1μL,R 1μL,ddH2O 7μL.利用ABI 7500型定量PCR儀(美國ABI公司)進行擴增,擴增體系為:50℃2 min;95℃10 min,95℃15 s,58℃30 s,40個循環.反應完成后進行溶解曲線分析,以確定引物的特異性.每個qRT-PCR測定設置3個重復.使用小麥Actin基因作為內參基因,并通過2-ΔΔCt法計算基因相對表達水平.
2 結果與分析
2.1 白粉菌脅迫下Brock小麥轉錄組測序分析
轉錄組測序結果顯示,小麥BMI組的平均clean reads數據量為61.61 M,BCI組的平均clean reads數據量為57.50 M.Q20(%)均大于96%,且只存在<1%的錯誤,說明測序結果良好,如表2所示.將獲得的轉錄組數據與小麥參考基因組序列(Ensembl Genomes release-35)進行對比,結果顯示每個文庫中約有91%的reads都能與參考基因組比對,且與參考基因組有唯一比對位置的reads大約為86%.以上結果證明測序數據可信,可用于后續分析.統計FPKM>1的轉錄本數據,構建維恩圖和火山圖分析兩組之間的差異基因,結果如圖1所示.BCI組樣品中,FPKM>1的轉錄本數量為45 195,BMI組為47 963.此外,從BCI和BMI樣品中檢測到unigenes分別為3 064條和5 832條.以P<0.05、|log2(fold change)|>|1|為條件進行DEGs的篩選分析,BMI vs.BCI共有4 625個差異表達基因,其中1 511個被上調,3 114個被下調,如圖1(b)所示.
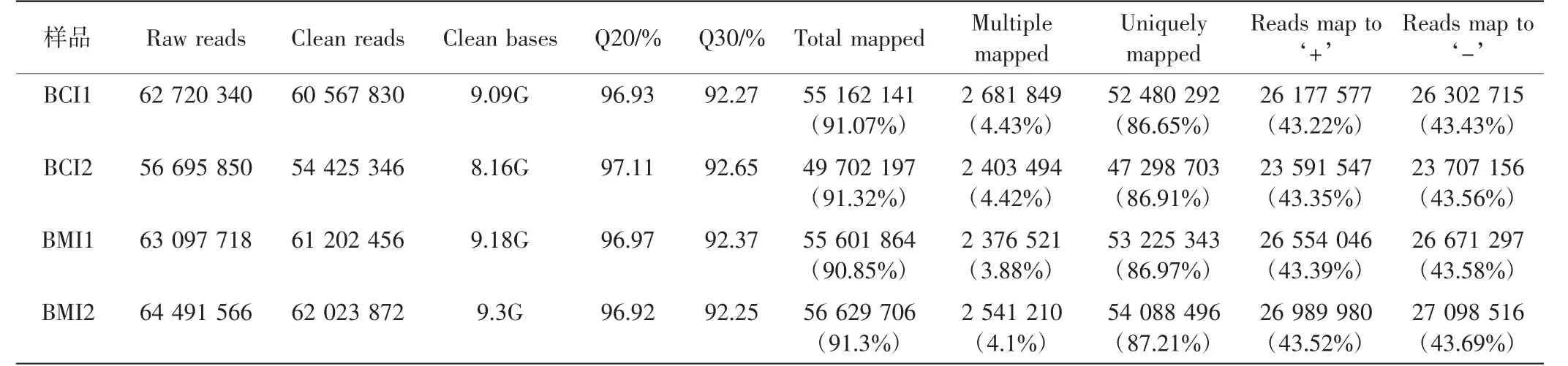
表2 轉錄組測序數據統計Tab.2 Summary for the transcriptome sequencing and assembly
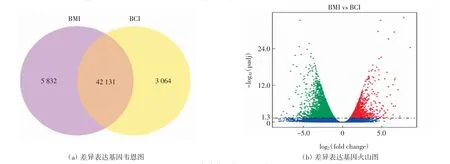
圖1 差異表達基因韋恩圖和火山圖Fig.1 Venn diagram and volcano map of differentially expressed genes
2.2 差異表達基因的功能注釋
為了探究白粉菌脅迫下小麥中的抗病基因表達模式,對差異表達基因進行了KEGG通路分析,富集結果如表3和圖2中所示.在BMI與BCI的比較中,上調轉錄本主要富集在亞麻油酸代謝、α-亞麻酸代謝和玉米素生物合成途徑,而下調轉錄本中顯著富集苯丙氨酸代謝、脂肪酸鏈伸長和植物-病原體互作通路.另外,在一組熱圖分析中顯示了白粉菌脅迫下主要參與基因的表達水平,例如植物-病原體互作通路和植物激素信號途徑等,如圖2所示.
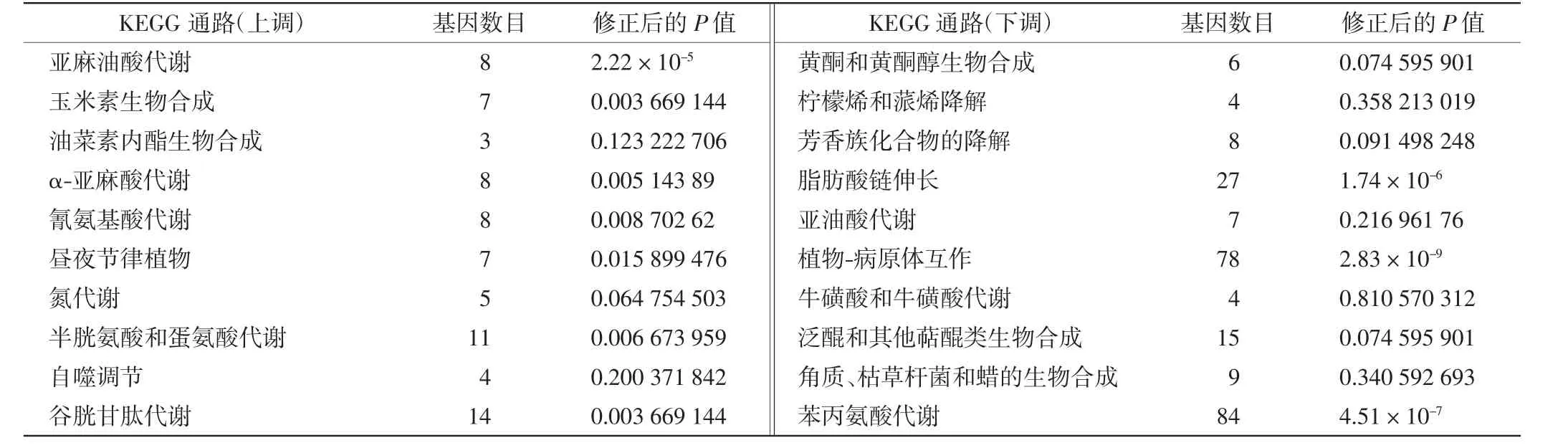
表3 KEGG富集分析Tab.3 Statistical enrichment analysis for KEGG pathways
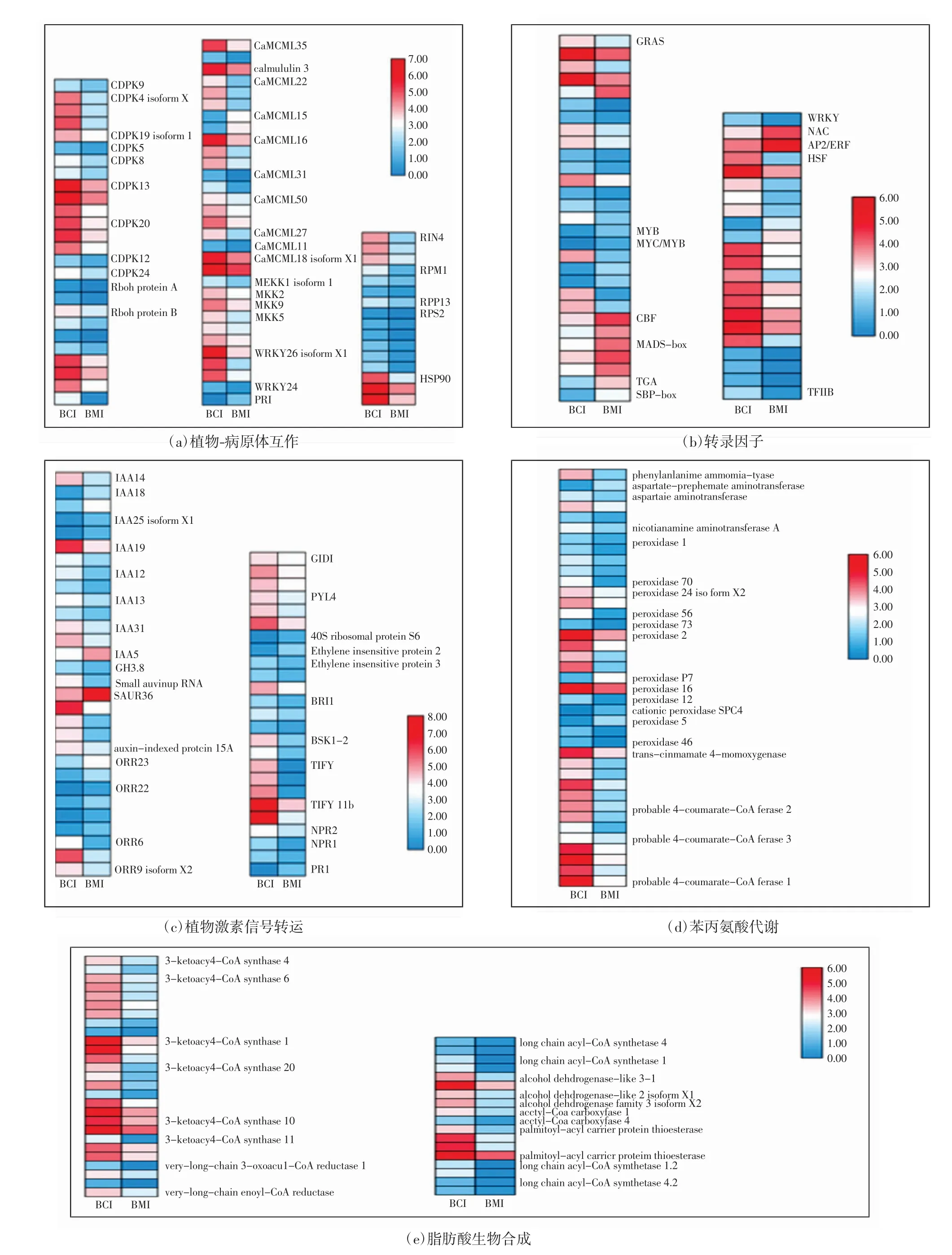
圖2 差異表達基因熱圖分析Fig.2 Heatmaps analysis of a group of differentially expressed genes
根據KEGG和熱圖分析結果,利用qRT-PCR檢測不同通路中的9個差異表達基因的表達模式:來自植物-病原體互作途徑中的4個DEGs,分別為PR1(pathogenesis-related proteins 1)、WRKY33(WRKYtranscription factor33)、RPM1(resistance to pseudomonas syringae pv.maculicola 1)和RIN4(RPM1 interacting protein 4);1個參與黃酮和黃酮醇的生物合成途徑的基因FOM1(flavone O-methyltransferase l);1個參與脂肪酸的生物合成和伸長基因LCACS4(long chain acyl CoA synthetase 4);2個參與植物激素信號的傳導基因NPR1和NPR2(nonexpressor of pathogenesis-related gene 1 and 2);1個編碼轉運蛋白基因ABCC10(ABC transporter family C10).接種白粉菌后,WRKY33、RPM1、RIN4、FOM1、LCACS4、NPR1和NPR2共7種DEGs的表達水平出現不同程度的降低,只有PR1和ABCC10的表達水平被上調,如圖3所示.
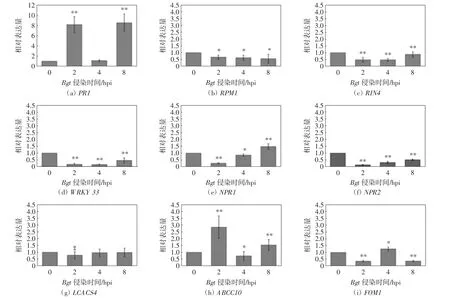
圖3 差異表達基因的表達模式分析Fig.3 Analysis of expression patterns of differentially expressed genes
2.3 RPM1和RIN4的表達模式分析
本研究中,病程相關蛋白PR1在2 hpi上調表達,為本底表達的8倍,說明由病原微生物觸發的免疫反應已經開始;而抗病基因RPM1在初期2~8 hpi內表達下調,且與之互作的RIN4表達在2~4 hpi下調,到8 hpi恢復到本底表達水平.為了進一步分析RPM1和RIN4在小麥抗白粉病應答反應初期的關聯,對抗病小麥Brock中RPM1和RIN4在0~72 hpi的表達趨勢進行檢測,發現RPM1的相對表達量在2 hpi降低,在12 hpi增加,在24 hpi達到最大,如圖4(a)所示,即為本底表達的2.3倍,隨后略有下降,到72 hpi時仍高于本底表達,約為1.8倍.而RIN4表達量雖然在24 hpi時略有升高,在2 hpi也略有降低,但總體趨勢變化不明顯,均徘徊在本底水平,如圖4(b)所示.
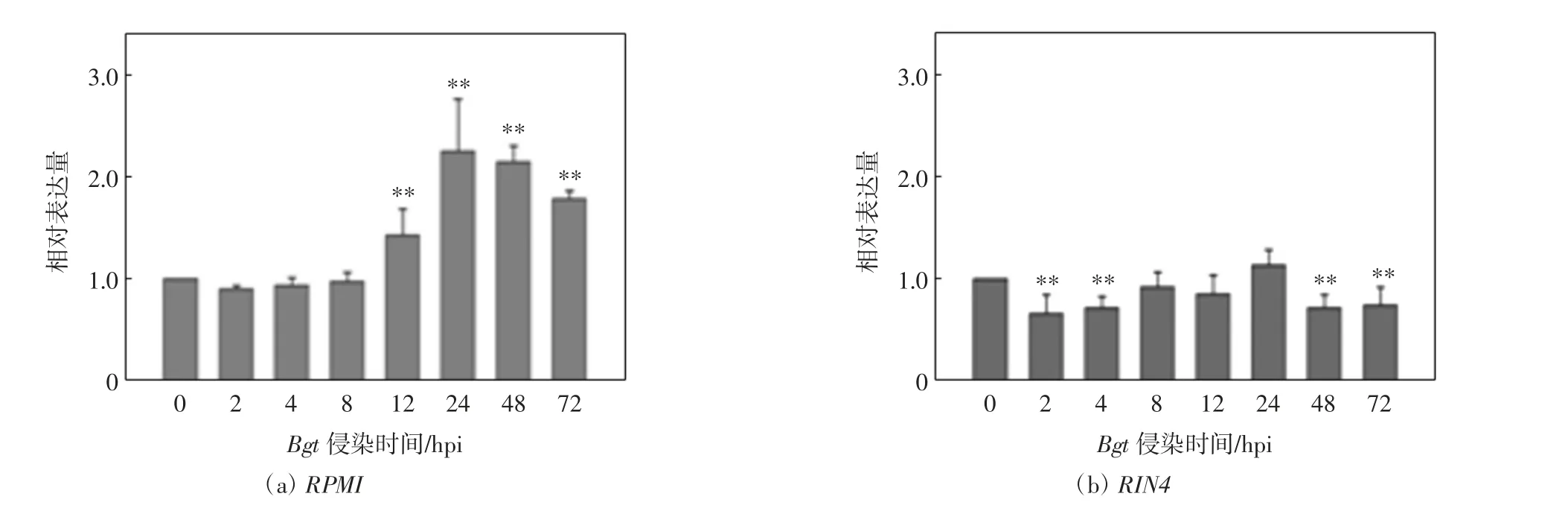
圖4 白粉菌侵染0~72 h RPM1和RIN4的表達模式分析Fig.4 Expression of RPM1 and RIN4 under powdery mildew stress at 0-72 hours post inoculation
3 討論與結論
3.1 早期抗病防御應答在植物抵御病原菌侵染中發揮重要作用
小麥白粉病是由布氏白粉菌侵染引起的真菌病害.白粉菌孢子接觸到介質后,20 s內產生細胞外基質(ECM),有助于白粉菌孢子附著到介質表面,激發植物表面防衛反應信號系統工作[5,7].ECM發揮作用的時間為12 hpi左右,之后ECM便降解.在8 hpi后,小麥葉片表面會有附著胞形成,15 hpi后,附著胞下開始形成侵入釘,在表皮細胞形成初生吸器[7].因此,8~15 hpi應該是白粉菌成功侵染小麥的關鍵階段.在本課題組早期研究中克隆得到了一個早期應答基因TaCOI1,并發現在2 hpi時該基因即在抗性小麥品種Brock中開始表達,且在抗病小麥中的基因應答速率明顯快于感病材料[8-9].由此說明小麥在病原菌侵染早期就已開始應答,故而探究植物與病原菌互作的早期應答網絡對了解小麥抗病機理具有重要意義.本研究對抗白粉病小麥品種Brock的早期應答基因進行了轉錄組測序,共鑒定出4 625個差異表達基因,分屬小麥與白粉菌互作應答的20條主要途徑.在已經公布的中國春小麥基因組序列中,Pm2基因的同源序列編號為TraesCS5D01G044600.1(TRIAE_CS42_5DS_TGACv1_457363_AA1485540.1).對本研究鑒定出的差異表達基因進行分析,結果發現,白粉菌侵染早期,TraesCS5-D01G044600.1的表達量沒有發生顯著變化.因此認為在白粉菌侵染早期(8 hpi以內),Brock中Pm2基因對白粉菌侵染應答不明顯.Brock可能通過其他的抗病基因或抗病反應通路發揮作用.
通過qRT-PCR技術檢測了來自不同途徑的9種DEGs的轉錄水平,發現在8 hpi之前,大部分基因表達下調,只有編碼病原菌與小麥互作蛋白的PR1和ABCC10基因的轉錄水平上調.說明來自不同途徑的多個基因均參與小麥與病原菌的早期互作反應,但病原菌侵染早期大多數基因首先被抑制表達.
3.2 早期抗病應答中的PTI和ETI響應時間
植物在遭受病原菌侵害時,為了保證自身的正常生長發育,逐漸進化出一套有序的分子調控機制[10].第一類是類受體激酶或類受體蛋白識別受體(RPPs)識別微生物/病原菌相關的分子模式(MAMPs/PAMPs)觸發的免疫反應(MTI/PTI)[11-12].PR1基因是PTI反應過程的一個標志性基因,PR1表達上調,意味著PTI被激活[13].為了進一步抵御親和性病原菌,植物逐漸進化出第二層防御屏障,稱為效應子觸發的免疫反應(ETI)[14].植物ETI過程主要依賴于抗病(R)基因產物,如RPM1[15]、MLA[16]等NBS-LRR類[17]基因.有報道RIN4作為植物基礎防御的調節劑,既可以調控PR1表達,也可以通過AvrRpm1和AvrB激活RPM1介導的植物抗性[18].
本研究發現在接種白粉菌2 hpi后,PR1的表達量平均增加了8倍,說明由病原微生物觸發的免疫反應在2 hpi就做出應答.抗病基因RPM1在接種白粉菌初期表達下調,12 hpi才開始增加,此后一直維持高水平表達.Wu等[19]在研究由Lr47介導的小麥抗葉銹病基因表達網絡中發現,TaPR1(GenBank NO.FJ815169.1)和TaRPM1(RPM1-AA1836940)基因的表達高峰分別發生在48 hpi和72 hpi.雖然總體趨勢相似,但本研究發現PR1和RPM1基因對白粉菌侵染的響應時間更早,分別為2 hpi和12 hpi,而RPM1的互作基因RIN4的表達量在0~72 hpi雖有變化,但不顯著,進一步說明RIN4作為“衛兵分子”,并不直接參與抗病反應,而是通過病原菌分泌蛋白,調節RPM1產生抗性[18,20].綜上,在小麥抗白粉病防御反應初期,PTI早在2 hpi就被觸發,ETI在12 hpi也開始響應,植物PTI和ETI協同作用抵抗白粉菌早期侵染.本研究結果為揭示小麥-白粉菌早期互作機制提供了新的實驗依據.
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
電子制作(2018年18期)2018-11-14 01:48:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
