不同耐藥鮑曼不動桿菌毒力效應檢測與分析*
2021-07-29 07:40:10林雪霏蔣月婷賴斯華鄧穎珊吳愛武
檢驗醫學與臨床 2021年14期
林雪霏,蔣月婷,賴斯華,鄧穎珊,吳愛武△
1.廣州醫科大學金域檢驗學院,廣東廣州 510182;2.中山大學熱帶病防治研究教育部重點實驗室,廣東廣州 510080;3.廣州醫科大學附屬第一醫院檢驗科,廣東廣州 510120
鮑曼不動桿菌是一種醫院內感染病原體,可在人類中引起多種感染,包括心內膜炎、腦膜炎、肺炎(在機械通氣患者中)、敗血癥和尿路感染[1]。由于鮑曼不動桿菌可以在醫院環境和設備上持續存在并以多藥耐藥病原體的形式出現[2],因此該菌也成為臨床的研究熱點,但以往臨床研究更多關注的是其耐藥性和耐藥機制,對該菌的生理機制及毒力研究關注度相對較低。細菌毒力的物質基礎主要包括侵襲力和毒素,其中侵襲力主要包括黏附素、外膜蛋白A、莢膜、侵襲性酶類和生物被膜。目前鮑曼不動桿菌中發現的毒力因子主要包括外膜蛋白A、脂多糖、外膜囊泡、鐵載體、生物被膜等[3]。在細菌耐藥性日益增強的大背景下,如果細菌在獲得耐藥性的同時,其毒力也相應增加,將嚴重影響臨床救治。本研究通過蹭行運動測定、明膠酶活性試驗、生物被膜形成試驗、紅細胞凝集及抑制試驗、常見毒力基因的檢測和小鼠體內毒力效應試驗對不同耐藥性鮑曼不動桿菌的毒力因子和毒力效應進行檢測,了解不同耐藥鮑曼不動桿菌在獲得耐藥性的同時其毒力改變情況,為臨床相應菌株的抗感染治療和毒力效應研究提供實驗室依據。
1 資料與方法
1.1菌株來源和鑒定 在廣州醫科大學附屬第一醫院微生物室菌種庫收集2016-2018年臨床分離的鮑曼不動桿菌共110株。隨后再用PCR方法檢測鮑曼不動桿菌特有的分子標記基因blaOXA-51-like,進一步確認110株菌株均為鮑曼不動桿菌。110株鮑曼不動桿菌中多重耐藥鮑曼不動桿菌(MDRAB) 42株,泛耐藥鮑曼不動桿菌(XDRAB)43株,相對敏感鮑曼不動桿菌(RDSAB) 25株。各耐藥菌株的定義如下[4],(1)MDRAB:對下述5類抗菌藥物中3類或者3類以上藥物不敏感的鮑曼不動桿菌,5類抗菌藥物包括氨基糖苷類、碳青酶烯類、頭孢菌素類、氟喹諾酮類和含有β-內酰胺酶抑制劑的復合制劑(包括哌拉西林/他唑巴坦、頭孢哌酮/舒巴坦、氨芐西彬/舒巴坦);(2)XDRAB:僅對1~2種抗菌藥物[主要指替加環素和(或)多黏菌素]敏感的鮑曼不動桿菌;(3)其余菌株都歸類為RDSAB。質控菌株:大腸埃希菌ATCC25922、銅綠假單胞菌ATCC27853由廣州醫科大學附屬第一醫院檢驗科微生物室饋贈。
1.2儀器與試劑 LB肉湯、營養瓊脂、明膠酶、D-甘露糖、結晶紫、甲醇、無水乙醇、1 mL注射器、細菌一次性培養皿、96孔聚苯乙烯板、96孔U型聚苯乙烯板均購自廣州捷倍斯生物科技有限公司,有效期內使用。6周齡的Balb/c雌性小鼠、鼠糧和墊料購自廣州晴樂生命科學有限公司;O型、AB型人紅細胞由廣州醫科大學附屬第二醫院檢驗科提供。PCR試驗中所有的引物合成和基因測序均由生工生物工程(上海)股份有限公司完成。
1.3方法
1.3.1生長曲線的測定[5]將從-80 ℃冰箱中復蘇的菌液用新鮮的LB肉湯通過比濁儀調成0.5麥氏濁度;吸取50 μL上述菌液用LB肉湯按1∶20稀釋成1 mL;在96孔聚苯乙烯板中加入200 μL稀釋的菌液,每株均設置3個復孔,其中空白對照只加LB肉湯;在搖床上以100 r/min進行培養,保持溫度在37 ℃,每隔2 h取出用酶標儀讀取吸光度值(A600)并記錄,監測至12 h結束。
1.3.2蹭行運動的測定[6]將在-80 ℃冰箱中保存的不同耐藥性的菌株在營養瓊脂平板上復蘇后,用無菌槍頭取單個菌落穿刺LB固體培養基接種于培養皿的接觸面;在37 ℃培養箱中孵育24 h后,測量其菌暈的長徑。若菌暈長徑>10 mm,則為蹭行運動陽性。本實驗需獨立重復3次,結果為3次試驗的平均值。以銅綠假單胞菌ATCC27853作為陽性對照。
1.3.3明膠酶活性檢測[7]將從-80 ℃復蘇的菌液用新鮮的LB肉湯通過比濁儀調成0.5麥氏濁度,取50 μL上述菌液接種于含30 g/L的明膠酶固體培養基上,放入37 ℃培養箱中培養24 h;取出后放置4 ℃冰箱4 h后觀察結果。若菌落周圍出現渾濁環,則為陽性,以銅綠假單胞菌ATCC27853作為陽性對照。
1.3.4結晶紫染色測定生物被膜的形成能力[8]將從-80 ℃冰箱中復蘇的菌液用新鮮的LB肉湯通過比濁儀調成0.5麥氏濁度;在96孔聚苯乙烯板中加入200 μL稀釋的菌液,每株菌株設置3個復孔,空白對照只加LB肉湯,在37 ℃細菌培養箱中靜置培養48 h;培養后吸出孔中上清液,用200 μL磷酸鹽緩沖液(PBS)清洗以去除浮游菌;加入200 μL 10%甲醇固定30 min,用200 μL PBS清洗隨后棄去;加入200 μL 1%結晶紫染液染色20 min,用200 μL PBS清洗隨后棄去;待孔中液體干燥后,每孔加入200 μL 95%乙醇,待孔中與生物被膜結合的結晶紫被乙醇完全溶解后,用酶標儀讀取各孔A570。3個復孔取均值A,未加菌液的空白LB肉湯均值為AC,將均值A與AC比較。細菌生物被膜形成能力分為4類:A≤AC為陰性;AC
1.3.5微量法進行D-甘露糖抵抗紅細胞凝集試驗[9]將復蘇的菌液用新鮮的LB肉湯通過比濁儀調至4麥氏濁度約為1.2×109CFU/mL,再稀釋成濃度為1.0×109CFU/mL的菌液,以4 000 r/min離心15 min,去除上清液;取離心沉淀的菌液用無菌PBS重懸制備新鮮的菌懸液(起始濃度為1×109CFU/mL)及用無菌PBS配制1%新鮮的AB型和O型的紅細胞懸液(其中加含或不含1%的D-甘露糖,各1份);在96孔U型聚苯乙烯板中加入制備好的菌懸液50 μL進行倍比稀釋;每個系列的孔中加入各50 μL含或不含1%甘露糖的AB型或O型的1%紅細胞懸液,于振蕩器上振蕩混勻;放入4 ℃冰箱靜置孵育2 h,觀察孔中是否有紅細胞凝集現象。若紅細胞在孔底聚集成小圓點,則為陰性;若紅細胞遍布整個孔底,則為陽性。以50 μL PBS中加入50 μL紅細胞懸液作為陰性對照,以大腸埃希菌ATCC25922作為陽性對照。
1.3.6毒力基因的檢測 用煮沸法提取細菌DNA。選取毒力基因BasD、BauA、Omp33-36、traT。25 μL反應體系:預混液12.5 μL,模板2 μL,10 μmoL/L的上下游引物各1 μL,滅菌雙蒸水補足體積。引物序列合成及擴增條件參考文獻[10],引物序列見表1。每種基因陽性條帶挑取1株菌株,將PCR擴增產物進行純化并送生工生物工程(上海)股份有限公司測序,測序結果提交到GenBank在線數據庫應用比對工具進行比對。

表1 毒力基因PCR擴增引物
1.3.7小鼠體內毒力實驗[6]隨機選取不同耐藥鮑曼不動桿菌各一株進行復蘇,將復蘇的菌液用新鮮的LB肉湯通過比濁儀調至4麥氏濁度約為1.2×109CFU/mL,再稀釋成濃度為1.0×109CFU/mL的菌液;稀釋后的菌液以1.0×109CFU/mL作為初始濃度進行10倍遞增的稀釋至菌液濃度為1.0×104CFU/mL,其中包括1.0×109、1.0×108、1.0×107、1.0×106、1.0×105、1.0×104CFU/mL 6個濃度;將57只小鼠隨機分成19組,每組3只,其中1~18組為感染組,19組為對照組;感染組每只小鼠注射200 μL菌液,對照組注射等量的生理鹽水;1~6組分別接種菌液濃度為1.0×109、1.0×108、1.0×107、1.0×106、1.0×105、1.0×104CFU/mL的RDSAB,7~12組接種上述濃度的MDRAB,13~18組接種上述濃度的XDRAB。接種后每天觀察小鼠的死亡情況,連續觀察10 d。
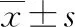
2 結 果
2.1生長曲線的測定 隨機選取不同耐藥性的菌株各10株檢測其生長能力。在培養的12 h里,隨著時間的增加其測定的A值也增加。RDSAB比MDRAB或者XDRAB生長得快,但差異無統計學意義(P>0.05)。見圖1。

圖1 不同耐藥性的鮑曼不動桿菌生長曲線
2.2蹭行運動的測定 在110株鮑曼不動桿菌中,除了2株RDSAB蹭行運動陽性外,其余菌株均為陰性。MDRAB和XDRAB均未觀察到蹭行運動,蹭行運動能力在組間分布差異無統計學意義(P>0.05)。
2.3明膠酶活性的檢測 在110株鮑曼不動桿菌中,全部菌株明膠酶活性均為陰性,不同耐藥表型的鮑曼不動桿菌產生以明膠酶為代表的侵襲性酶類的能力無差異。
2.4不同耐藥性鮑曼不動桿菌生物被膜形成能力測定 110株試驗株均能形成生物被膜。RDSAB的生物被膜形成能力比MDRAB或XDRAB的生物被膜形成能力強,差異有統計學意義(P<0.05)。25株RDSAB生物被膜形成能力以強陽性(56.00%,14/25)為主,其次為陽性(36.00%,9/25)。而42株MDRAB和43株XDRAB表現為以陽性為主,分別占59.52%(25/42)和69.77%(30/43);其次為弱陽性,分別占35.71%(15/42)和18.60%(8/43)。見表2。

表2 不同耐藥性的鮑曼不動桿菌生物被膜形成能力的比較[n(%)]
2.5紅細胞凝集試驗結果 在110株鮑曼不動桿菌中,只有1株RDSAB在有或者無D-甘露糖存在的條件下,均能與AB型和O型紅細胞發生凝集,但組間比較差異均無統計學意義(P>0.05)。
2.6毒力基因的檢測 常見毒力基因BasD、BauA、Omp33-36和traT中,除了基因traT無陽性擴增外,其他基因均可檢出,部分陽性擴增結果見圖2。

注:M為DNA marker 2000;1為BasD基因(868 bp);2為BauA基因(224 bp);3為Omp33-36基因(726 bp);N為陰性對照。
在110株鮑曼不動桿菌中,毒力基因BasD具有較高的檢出率,為59.1%(65/110);毒力基因BauA和Omp33-36檢出率較低,分別為6.4%(7/110)和3.6%(4/110);毒力基因traT無檢出。見表3。

表3 不同耐藥性鮑曼不動桿菌常見毒力基因的檢出結果[n(%)]
對比不同耐藥性的鮑曼不動桿菌毒力基因的檢出情況,MDRAB的BasD毒力基因檢出率比RDSAB高,差異有統計學意義(P<0.05);而RDSAB的毒力基因Omp33-36檢出率比MDRAB和XDRAB高,差異均有統計學意義(P<0.05);MDRAB的BauA和BasD毒力基因檢出率比XDRAB高,差異有統計學意義(P<0.05)。見表4、5、6。

表4 RDSAB和MDRAB檢出的毒力基因結果比較(n)

表5 RDSAB和XDRAB檢出的毒力基因結果比較(n)
2.7小鼠體內毒力實驗 對3種不同耐藥性的鮑曼不動桿菌進行小鼠體內毒力實驗,經過10 d的觀察,實驗組和對照組無1例小鼠發生死亡。

表6 MDRAB和XDRAB的毒力基因的比較(n)
3 討 論
近些年來對于鮑曼不動桿菌的研究主要集中在流行特征和耐藥機制方面,而對鮑曼不動桿菌的毒力機制研究比較少。目前鮑曼不動桿菌中發現的毒力因子主要包括外膜蛋白A、脂多糖、磷脂酶D、外膜囊泡、生物被膜、鐵載體、運動能力等[3]。
鮑曼不動桿菌是機會致病菌,廣泛存在于醫院環境中并長期存活,可通過醫療器械或者醫務人員進行克隆性傳播[2],體外存活能力是病原菌重要的致病條件。本研究中對不同耐藥性的鮑曼不動桿菌進行生長能力的測定,結果表明在體外不同耐藥性的鮑曼不動桿菌生長能力比較,差異無統計學意義(P>0.05),表明耐藥性的獲得并未導致菌株體外生存能力減弱。
細菌的運動性與其引起疾病的能力密切相關。鮑曼不動桿菌無鞭毛不能主動運動,但其具備Ⅳ型菌毛可以介導細菌產生不依賴鞭毛的獨特運動,此運動叫做蹭行運動[11]。在許多革蘭陰性菌中,Ⅳ型菌毛通過介導蹭行運動,參與細菌在宿主體內黏附和定植并引起免疫應答,同時還在生物被膜形成早期發揮作用[12]。本研究中,MDRAB和XDRAB均未檢測到蹭行運動,而RDSAB雖然有2株菌株蹭行運動陽性,但與MDRAB、XDRAB比較,差異無統計學意義(P>0.05),說明不同耐藥表型鮑曼不動桿菌在蹭行運動能力方面無明顯差異,提示感染時這些菌株的黏附能力可能也無差異。
明膠酶是細菌產生的一種侵襲性酶類,是具有水解明膠、血紅蛋白、膠原蛋白及其他生物活性肽功能的金屬蛋白酶,可增強細菌感染性炎癥發生時的致病力[7]。本研究中,110株菌株均未檢測出明膠酶活性,說明不同耐藥表型的鮑曼不動桿菌產生以明膠酶為代表的侵襲性酶類的能力無明顯差異。
鮑曼不動桿菌生物被膜的形成與其耐藥性和致病力之間的相關性報道較多,但研究結果差異較大。BARDBARI等[13]研究顯示MDRAB生物被膜形成能力比非MDRAB生物被膜形成能力強。而QI等[14]研究認為隨著生物膜形成能力的增強其耐藥性相應降低。本研究結果顯示,RDSAB的生物被膜形成能力比MDRAB或XDRAB的生物被膜形成能力強,差異有統計學意義(P<0.05)。RDSAB生物被膜形成能力以強陽性(56.00%)為主,其次為陽性(36.00%);而MDRAB和XDRAB以陽性為主,分別占59.52%和69.77%,其次為弱陽性,分別占35.71%和18.60%。耐藥菌株生物被膜形成能力較相對敏感菌株弱,表明菌株耐藥性的獲得使其生物被膜的形成能力相應減弱,導致其在致病能力方面可能也會相應減弱,這可能是菌株為了在人體更好地生存下來而表現出的一種對人體的適應性改變,具體機制有待進一步探究。
細菌的菌毛除了可以使細菌黏附于宿主細胞,對紅細胞也有凝集能力。細菌的致病力與細菌的黏附能力、血凝能力相關,血凝能力強的細菌毒力也強。在本研究菌株的紅細胞凝集試驗中,僅1株RDSAB在有或者無D-甘露糖存在的條件下,均能與AB型和O型紅細胞發生凝集,具有紅細胞凝集能力,其余菌株紅細胞凝集試驗均為陰性。試驗結果提示鮑曼不動桿菌整體凝集紅細胞的能力弱,不同耐藥表型的鮑曼不動桿菌對紅細胞的凝集能力無明顯差異。
鮑曼不動桿菌常見的毒力基因有介導與宿主細胞結合的黏附和侵襲相關毒力基因Omp33-36,鐵載體的合成和運輸相關毒力的基因(BasD和BauA)及編碼R6-5質粒特異的外膜蛋白的毒力基因traT[10]。本研究結果顯示,在鮑曼不動桿菌中,除了毒力基因BasD具有較高的檢出率(59.1%)外,毒力基因BauA和Omp33-36檢出率較低,未檢測到毒力基因traT,總體上,研究結果顯示鮑曼不動桿菌毒力基因檢出率不高,說明鮑曼不動桿菌是一種相對低毒力的細菌,與該菌在臨床上主要引起免疫功能低下或感染的現狀相吻合。對比不同耐藥表型的鮑曼不動桿菌毒力基因的檢出情況,MDRAB的BasD毒力基因檢出率比RDSAB高,MDRAB的BauA和BasD毒力基因檢出率比XDRAB高,而RDSAB的毒力基因Omp33-36檢出率比MDRAB和XDRAB高。這些結果提示,在不同耐藥表型鮑曼不動桿菌中均可檢出各占優勢的毒力基因,說明這些不同耐藥表型的鮑曼不動桿菌毒力特征可能類似,差異不明顯。
小鼠體內細菌感染毒力實驗結果顯示,將不同濃度不同耐藥表型的鮑曼不動桿菌菌株在小鼠體內進行注射感染,感染10 d后所有小鼠均未出現死亡。實驗結果提示不同耐藥表型的鮑曼不動桿菌均為低毒力的菌株,對動物致病能力不強。
綜上所述,不同耐藥表型的鮑曼不動桿菌除了在生物被膜形成能力方面敏感菌株比耐藥菌株強以外,在其他的細菌毒力檢測試驗中,不同耐藥表型鮑曼不動桿菌之間無明顯差異,也就是說菌株在獲得耐藥性的同時其毒力并沒有相應增加。鮑曼不動桿菌作為重要的院內感染菌,其致病性、耐藥性逐漸增強的同時伴隨的是治療難度的增加,因此,臨床和實驗室應注意監測其毒力變化趨勢,為臨床治療提供依據。
猜你喜歡
發明與創新(2022年30期)2022-10-03 08:40:56
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
動漫星空(興趣百科)(2020年12期)2020-12-12 05:31:40
科學大眾(2020年12期)2020-08-13 03:22:22
人大建設(2018年6期)2018-08-16 07:23:10
文理導航·科普童話(2017年5期)2018-02-10 19:42:14
無人機(2017年10期)2017-07-06 03:04:36
小星星·閱讀100分(低年級)(2015年10期)2015-10-22 08:30:04
