四川臥龍國家級自然保護區雪豹地棲動物群落初探
2021-08-03 06:08:42周厚熊吳艷蓉王鵬彥施小剛李兆元
野生動物學報 2021年3期
周厚熊 姜 楠 李 君 楊 虎 吳艷蓉 王鵬彥 施小剛 李兆元*
(1.西南林業大學生物多樣性保護學院,昆明,650224;2.昆明逸境生態工程咨詢有限公司,昆明,650224;3.四川臥龍國家級自然保護區管理局,阿壩,623006)
雪豹(Pantherauncia)隸屬于食肉目(Carnivora),貓科(Felidae),豹屬[1],全球種群數量7 463—7 980 只[2],國外分布于阿富汗、不丹、印度、哈薩克斯坦、吉爾吉斯斯坦、蒙古、尼泊爾、巴基斯坦、俄羅斯、塔吉克斯坦和烏茲別克斯坦[3],國內分布于西藏、新疆、青海、甘肅、四川、云南[4],其中四川臥龍國家級自然保護區是雪豹全球分布的東南邊界[5]。作為山地生態系統的旗艦種和頂級捕食者,雪豹在維持生態系統的健康和穩定中起著極其重要的作用[6]。到目前為止,雪豹所生活的地棲動物群落構成及其在群落中的生態學作用還不清楚。
保護生物學理論認為,頂級捕食者的存在是判斷一個生態系統健康與否的重要標志[7]。然而,頂級捕食者如何影響生態系統健康需要從群落角度去認識。Odum[8]、Ricklefs[9]以及Putman[10]認為,群落是物種通過生態學關系(如:捕食、競爭、共生等)結成的整體。在這個整體中,生態學關系支撐物質和能量在物種間的傳遞,以實現生態系統的功能。因此,群落不是一群物種的簡單集合;它是物質和能量傳遞過程中涉及的物種的集合。生態學關系是理解群落結構的關鍵。然而,傳統的生態學研究難以采集物質和能量作為通貨,同時量化各個物種間的生態學關系[11]。由于所有種間生態學關系都需通過物種種群有規律地出現在特定地理空間中才得以實現,因此物種的生態學關系必然呈現在物種的空間關聯之中。為此,本研究利用四川臥龍國家級自然保護區紅外相機記錄,通過種間空間關聯構建雪豹所生活的地棲動物群落,從群落角度分析該物種的生態學作用,探討它對維持高山生態系統的意義,從而為保護管理工作提供科學建議。
1 研究區域概況
臥龍國家級自然保護區(30°45′—31°25′N,102°52′—103°25′E)位于四川省汶川縣,屬成都平原向青藏高原過渡的高山深谷地帶,面積約2 000 km2(圖1)。保護區地勢東南低、西北高,海拔1 150—6 250 m。保護區內有耿達河、皮條河、正河、西河和中河,均屬岷江水系。地表景觀隨海拔上升依次為常綠闊葉林、常綠落葉闊葉混交林、落葉闊葉林、針闊混交林、針葉林、亞高山灌叢和草甸,以及高山流石灘[12]。保護區內分布有大熊貓(Ailuropodamelanoleuca)、雪豹、小熊貓(Ailurusfulgens)、黑熊(Ursusthibetanus)、黃喉貂(Martesflavigula)、藏酋猴(Macacathibetana)、川金絲猴(Rhinopithecusroxellana)、羚牛(Budorcastaxicolor)、巖羊(Pseudoisnayaur)、林麝(Moschusberezovskii)、綠尾虹雉(Lophophoruslhuysii)、紅喉雉鶉(Tetraophasisobscurus)、血雉(Ithaginiscruentus)、白馬雞(Crossoptiloncrossoptilon)、藏雪雞(Tetraogallustibetanus)以及金雕(Aquilachrysaetos)等珍稀物種。
2 研究方法
2.1 數據采集
2017年4月—2018年4月,在保護區內皮條河東岸隨機選擇3個20 km2的樣區(圖1),所選樣區海拔1 749—4 430 m。樣區涵蓋所有生境類型,包括常綠闊葉林、落葉闊葉林、針闊混交林、針葉林、灌叢、高山草甸、流石灘。用ArcGIS 11軟件將樣區劃分成60個1 km×1 km的公里網格,每個網格為1個采樣單元。將網格內占地>50%的生境類型定義為采樣單元的生境類型,并在其中安裝1臺紅外相機(易安衛士L710)。相機固定在穩固的樹干或巖石上,距地面30—50 cm,并用枯枝落葉進行隱蔽。同時,記錄位點的經緯度、海拔、坡位、坡度、坡向、地物類型、森林演替狀態、植被高度和植被蓋度。采用Wei等[13]采集生境因子的方法。每臺相機配置16 GB內存卡,相機時間設定為北京時間,靈敏度設定為中,拍攝間隔時間1 s,觸發后連續拍攝3 次,隨后錄像20 s。電池和內存卡每隔3個月更換1次。物種鑒定參考《中國鳥類野外手冊》[14]和《四川獸類原色圖鑒》[15]。在安裝相機以及更換相機電池及內存卡時,數據采集者對相機位點周圍進行勘察,記錄所發現的動物實體及痕跡。痕跡鑒定參考《中國獸類蹤跡指南》[16]。所有照片數據導入電腦,經整理后錄入Microsoft Excel,以供分類、計算和匯總。
2.2 物種相關性分析
對各位點照片進行物種鑒定,獲取物種存在與否的二分型數據。針對這種二分型數據,采用phi系數對種間關聯度進行衡量[17]。
在進行Phi系數計算時,首先建立2×2列聯表(表1)。將全部相機位點分為4組:第1組為物種X和物種Y均出現的相機位點,記為A組;第2組是物種X不出現,但物種Y出現的位點,記為B組;第3組是物種X出現,物種Y不出現,記為C組;第4組是物種X和物種Y均不出現的位點,記為D組。計算各組位點數,填入列聯表。表1中,N=A+B+C+D,即:總相機位點數。

表1 Phi系數2×2列聯表
Phi系數(rφ)計算公式如下:
(1)
rφ取值0—1:接近0,表示種間空間關聯性弱;接近1,表示關聯性強。如果(AD-BC)>0,表明種間存在正關聯;如果(AD-BC)<0,則種間存在負關聯。在本研究中,2個物種共同出現于相同空間頻率越高,phi系數越大。在捕食者-獵物關系中,Phi系數越大,表明捕食者物種對獵物物種的捕食壓力越大以及獵物物種對捕食者物種的生存意義越大;在對相同資源競爭的物種中,則表明種間的競爭越激烈。
進一步用卡方檢驗求取上述phi系數值的顯著性,公式如下:
(2)
當χ2≥3.84時,P≤0.05,表明計算所獲rφ值有意義,是生態學關系導致的空間關聯,可用于研究分析。當χ2<3.84時,P>0.05,表明計算所獲rφ值無意義,關聯現象是隨機因素導致的,不能用于研究分析。
用lambda系數幫助判斷存在直接空間關聯的食肉目動物間的實際生態學關系,計算過程參照李兆元等[17]。
2.3 群落構建
依據上述種間關聯性計算,找出具有顯著正關聯的種對。通過種對關系,將物種構建成空間關聯網絡。最后,通過生態學文獻及野外工作中發現的證據,將種對空間關聯關系還原成生態學關系,從而獲得群落物種網絡。
2.4 種間關聯多樣性
群落的總體穩定性和抗干擾力與群落的整體復雜性緊密相關。本研究用香農-威納指數(Shannon-Wiener)來衡量群落的整體復雜性,計算如下:
(3)
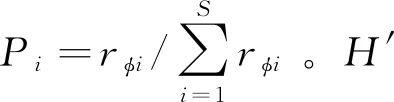
2.5 生態因子分析
采用卡方吻合度檢驗分析雪豹分布的生態制約因子。分析群落分布的生態制約因子涉及多個物種,各生態因子也涉及多種表達,如:生境類型。因此,采用克萊姆系數(C)進行檢驗,計算過程參照李兆元等[17]。
3 結果與分析
3.1 地棲動物區系
在60個公里網格中,紅外相機的總工作時間為16 966臺日,獲得清晰照片7 120張,鑒定出6目15科35種地棲動物。其中,獸類(Mammalia)有5目14科26種,分別為喜馬拉雅旱獺(Marmotahimalayana)、豪豬(Hystrixbrachyura)、藏鼠兔(Ochotonathibetana)、藏酋猴、川金絲猴、狼(Canislupus)、赤狐(Vulpesvulpes)、黑熊、大熊貓、小熊貓、黃喉貂、石貂(Martesfoina)、黃鼬(Mustelasibirica)、香鼬(M.altaica)、豬獾(Arctonyxcollaris)、花面貍(Pagumalarvata)、豹貓(Prionailurusbengalensis)、雪豹、野豬(Susscrofa)、林麝、毛冠鹿(Elaphoduscephalophus)、水鹿(Rusaunicolor)、羚牛、中華鬣羚(Capricornismilneedwardsii)、中華斑羚(Naemorhedusgriseus)和巖羊;鳥類(Aves)1目1科9種,分別為雪鶉(Lerwalerwa)、紅喉雉鶉、藏雪雞、血雉、紅腹角雉(Tragopantemminckii)、勺雞(Pucrasiamacrolopha)、綠尾虹雉、白馬雞和紅腹錦雞(Chrysolophuspictus)。此外,還記錄到山羊、黃牛、牦牛和村民。在35種野生動物中,食肉目物種豐度最高,含6科13種;其余依次為偶蹄目(Artiodactyla)4科8種,雞形目(Galliformes)1科9種,嚙齒目(Rodentia)2科2種,靈長目(Primates)1科2種,兔形目(Lagomorpha)1科1種。
雪豹在8個相機位點中出現,海拔范圍4 072—4 430 m,平均海拔4 252 m;屬于高山苔原生物相,生境景觀類型包括灌叢、流石灘和高山草甸。海拔分布重疊或部分重疊的物種有18個,包括喜馬拉雅旱獺(海拔范圍:3 718—4 430 m,平均4 125 m)、赤狐(3 292—4 430 m,3 980 m)、香鼬(4 303—4 338 m,4 320 m)、石貂(4 303—4 338 m,4 249 m)、豹貓(1 979—4 303 m,3 004 m)、豬獾(2 473—4 430 m,3 394 m)、巖羊(2 473—4 430 m,4 024 m)、藏鼠兔(2 473—4 430 m,4 151 m)、斑羚(1 749—4 430 m,2 924 m)、血雉(3 042—4 303 m,3 733 m;灌叢、流石灘、草甸)、雪鶉(3 186—4 430 m,4 048 m;灌叢、流石灘、草甸)、紅喉雉鶉(3 718—4 052 m,3 912 m;灌叢)、藏雪雞(4 152—4 300 m,4 226 m)、綠尾虹雉(3 093—4 338 m,3 841 m;灌叢、流石灘、草甸)。
卡方吻合度檢驗結果顯示,雪豹分布與海拔、坡向、地物類型和植物高度存在顯著相關性(表2),分布于海拔4 000 m以上、西向、灌叢和流石灘區域,回避低海拔高植被區域。植被蓋度、坡度和坡位對該物種的分布沒有顯著性影響。
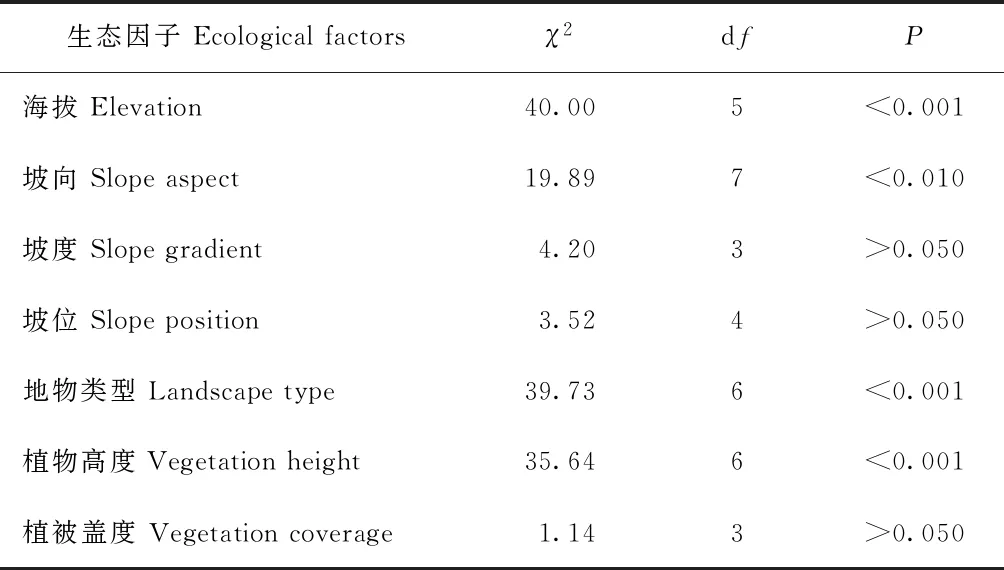
表2 雪豹分布與各生態因子相關性檢驗結果(卡方吻合度檢驗)
3.2 地棲動物空間關聯網絡
3.2.1 關聯網絡構成
Phi系數計算結果顯示,35種同域分布的地棲動物中,有13種與雪豹存在直接或間接空間正關聯,包括喜馬拉雅旱獺、赤狐、香鼬、石貂、巖羊、藏鼠兔、血雉、白馬雞、豬獾、雪鶉、紅喉雉鶉、綠尾虹雉和藏雪雞。這些物種與雪豹在海拔分布上重疊,共同構成空間關聯物種網絡(圖2)。網絡中共有14個物種,占區系物種數40%。這些物種結成38個種對,關聯多樣性(香農-威納指數)為7.30。在這個網絡中,赤狐與血雉的關聯度最強(0.67),其次是紅喉雉鶉和白馬雞(0.64)、雪豹和巖羊(0.63);與雪豹關聯度最高的是巖羊(0.63),其次是雪鶉(0.59)和石貂(0.59)。網絡結構大致分為2層:外層是藏雪雞、香鼬、藏鼠兔、白馬雞、紅喉雉鶉、豬獾和雪鶉,每個物種的種對最多4個,關聯系數平均為0.47;內層為綠尾虹雉、喜馬拉雅旱獺、血雉、赤狐、石貂、巖羊和雪豹,各物種的種對數至少7個,平均關聯系數是0.48。內、外層關聯系數無顯著性差異(F=0.75,df=55,P>0.05;t=0.98,df=55,P>0.05),但種對數存在顯著性差異(χ2=4.57,df=1,P<0.05),表明內層物種對維持網絡穩定有主要作用。其中,雪豹為大型食肉類,在本網絡中是最大的捕食者,可能是頂級捕食者;赤狐為中型捕食者。這2種捕食者沒有發生直接空間關聯,而是通過共同的關聯物種(包括綠尾虹雉、喜馬拉雅旱獺、石貂和巖羊)發生間接關聯,可能表明由于食性競爭而相互回避。雪豹與石貂、喜馬拉雅旱獺、巖羊、雪鶉空間關聯強(rφ>0.5),表明雪豹對這些物種可能依賴性強。赤狐與白馬雞、紅喉雉鶉、血雉、巖羊,以及喜馬拉雅旱獺也存在很強的空間關聯,可能對這些物種存在強的依賴性。值得注意的是,2種中大型食草動物巖羊、喜馬拉雅旱獺同時與雪豹、赤狐產生顯著空間正關聯,對支撐雪豹和赤狐的生存均表現出重要意義;小型食肉動物石貂也同時與雪豹、赤狐產生顯著空間關聯,表明食肉動物之間可能存在相互捕食或一方為另一方創造有利條件的關系。
3.2.2 種間非對稱關聯
上述物種網絡中,體形大小迥異但存在顯著空間正關聯的捕食物種種對有雪豹-石貂、雪豹-香鼬、赤狐-石貂、赤狐-豬獾、石貂-香鼬。Lambda系數檢驗結果顯示:石貂能預測雪豹出現(λB>0.01,P≤0.05),表明雪豹捕食石貂;香鼬能預測石貂出現(λB>0.03,P≤0.05),表明石貂捕食香鼬;豬獾能預測赤狐出現(λB>0.03,P≤0.05),表明赤狐捕食豬獾;雪豹與香鼬、赤狐與石貂不存在空間非對稱關聯(λB<0,P>0.05)。
3.2.3 物種網絡與生態因子的關系
網絡中的物種總體隨海拔升高出現頻次增加,其中,在3 531—4 430 m區段內出現的頻率占總頻率81.30%,為主要分布區段。克萊姆系數檢驗結果(表3)顯示,該網絡的分布與坡向、生境類型、森林演替狀態、植被高度和植被覆蓋度具有顯著相關性,坡度和坡位沒有顯著相關性;網絡中的物種在灌叢、草甸(或草地)、流石灘以及無林空間中出現的頻率最高,占78.05%;表明這些生境類型是網絡賴以存在的基礎;這些生境類型主要分布于海拔3 718—4 430 m區段中。
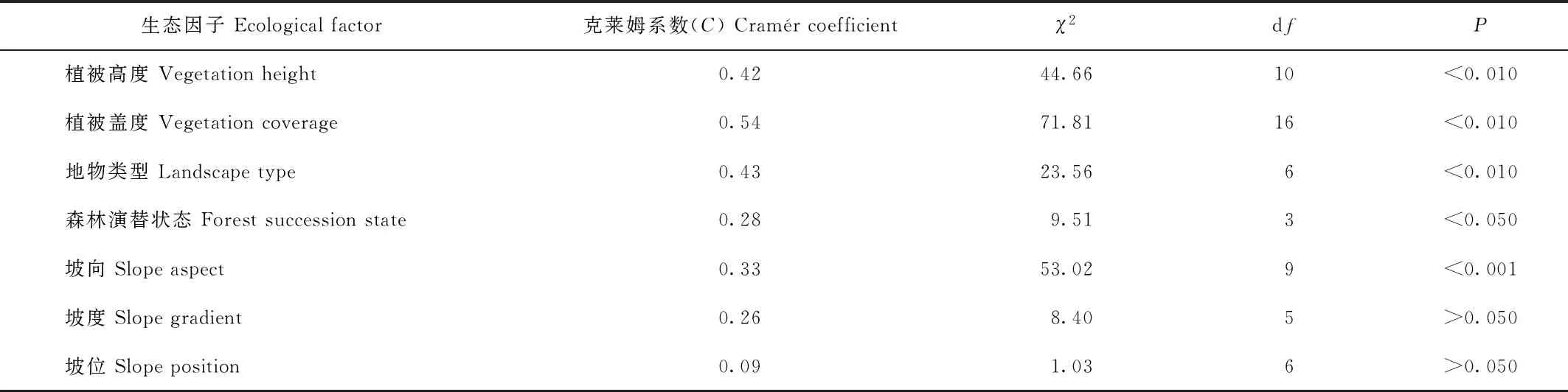
表3 物種分布網絡與生態因子的相關關系
綜上所述,該物種網絡分布于海拔3 718 m以上,植被稀疏的灌叢、草甸(或草地)和流石灘生境中。網絡由高海拔物種(旱獺、雪豹、香鼬、石貂、白馬雞、紅喉雉鶉、藏雪雞)以及海拔廣布種的高海拔種群(赤狐、巖羊、藏鼠兔、血雉、豬獾、雪鶉、綠尾虹雉)構成。
4 結論與討論
4.1 數據采集方法及質量評估
以往關于物種空間關聯的研究主要采用樣線法收集動物在沿線留下的痕跡(如毛發、糞便、取食痕跡等),從而獲取動物在特定空間中出現與否的數據[18-19]。這種方法存在以下問題:①考慮到數據可比性,所有地理空間中的數據需要同時采集。但是,樣線法無法使研究者同時進行大面積數據采集。②如果動物未留下痕跡,所獲數據可能非常不完整。③在確定不同地點發現的痕跡是否屬于分布于同一地理空間時,樣線法無法提供客觀的判斷依據。隨著紅外相機技術的出現,這些問題逐步得到有效解決。第一,紅外相機可以長時間連續工作,準確提供相機位點附近物種的出現情況[20-21],同時布設多臺相機,可以有效解決上述第1個問題。第二,紅外相機在無人操作的情況下可以自動觸發拍照,獲得動物活體照片,很大程度解決上述第2個問題。第三,相機位點可以作為客觀參照物,位點附近出現的動物痕跡以及相機拍攝到的動物可以視為相同地理空間中的存在物,這樣解決了上述第3個問題。
與其他研究方法一樣,紅外相機技術也存在特有問題。首先,紅外相機依靠溫血動物身體溫度與環境溫度的溫差來觸發。冷血動物(兩棲類Amphibia、爬行類Reptilia)身體溫度與環境溫度相似,無法觸發相機拍照,因此該技術無法獲取冷血動物的照片記錄。其次,李晟等[20]研究發現紅外相機對小型哺乳動物圖像捕獲率偏低。這種技術缺陷可能會影響研究結果。例如,魏萬紅等[22]研究表明香鼬主要獵物是藏鼠兔;但本研究結果顯示,這2個種沒有顯著空間關聯性。藏鼠兔是小型哺乳動物,可能存在紅外相機對其影像捕獲率偏低的問題,但有待驗證。本研究所獲數據中,小型哺乳動物似乎存在整體上的代表性偏低問題。空間關聯計算中,2個物種共同出現在相同地理空間中的頻率是建立種間關聯的基礎,受物種種群密度影響。因此,2項研究結果的差異可能由紅外相機的技術缺陷導致。當然,也可能是數據采集期間藏鼠兔實際種群密度偏低所致。然而,這個缺陷沒有影響中大型食肉動物雪豹的種間關系分析。
4.2 統計學方法選擇
地棲動物基本都有家域[23]。部分物種為獨居生活類型,其他為群居類型。如果相機布設在某一個體或群體的家域核心區,相同個體在相機探測區域反復活動,活動頻率與種群密度的換算關系因動物生活類型不同而異。現有紅外相機技術無法進行圖像識別,研究者憑肉眼對個體進行識別所獲結果不具備可靠性,尤其是對夜行性動物的識別(圖像清晰度低,黑白色)。因此,特定位點相機所獲“獨立有效照片數”[20]不能用作物種密度的指示,而只能代表物種出現與否。針對這種二分型數據,只能選擇非參數統計中的Phi系數來量化2個物種的空間關系[17]。
在一個群落中,食肉目物種常常不止一種。當體形差異較大的2種食肉目動物出現空間關聯時,關聯性可能是它們在相同地理空間中共同取食某種獵物(對共同食物的需求)所致,也可能是體形大的物種捕食體形小的物種(捕食關系)所致。此時,在沒有相關文獻支持下,難以判斷它們的種間生態學關系。然而,在捕食關系中,被捕食者傾向于在其生境內隨機活動,以擺脫被捕食;捕食者則依據被捕食者的活動來決定自己的活動空間,以便獲得捕食機會。因此,兩者的空間關聯性不對稱,可以用Lambda系數檢驗預測2個物種的空間出現[17]。
4.3 地棲群落構成
本研究所獲數據均為地棲動物空間分布數據。基于這些數據建立的物種網絡是個地棲物種網絡,通過生態學關系關聯起來形成一個地棲動物群落。雪豹是地棲動物,它的所有生態學關系發生在這個地棲動物群落中。
Schaller等[24]、劉楚光等[25]通過糞便(樣本量分別為206、644 份)研究發現,喜馬拉雅旱獺、巖羊是雪豹的重要食物來源,2種獵物的殘留物在糞便中出現頻次相當,說明喜馬拉雅旱獺和巖羊在雪豹食譜中的重要性相似。本研究結果顯示,雪豹與喜馬拉雅旱獺以及雪豹與巖羊的空間關聯系數相似,分別為0.57和0.63;表明雪豹對喜馬拉雅旱獺和巖羊的捕食壓力比較接近,與上述研究結果一致。Oli等[26]在尼泊爾的研究發現,雪豹捕食小型食肉類石貂、伶鼬(Mustelanivalis),在213份雪豹糞便中,石貂的殘留物有8份,伶鼬的殘留物有10份,說明雪豹作為機會主義捕食者也捕食中小型食肉類。本研究結果顯示,石貂能預測雪豹出現(λB>0.01,P≤0.05),表明雪豹捕食石貂,與上述結果一致。香鼬不能預測雪豹出現,表明雪豹不會規律性地捕食香鼬。從雪豹與喜馬拉雅旱獺的捕食關系和香鼬與喜馬拉雅旱獺較強的空間關聯性來推測,雪豹與香鼬之間可能是掠食關系,即雪豹捕食喜馬拉雅旱獺后,香鼬再利用剩余部分。Oli等[26]研究發現,雪豹捕食雞形目鳥類,但鑒于當年的技術條件無法鑒定是哪一種。根據研究地雞形目鳥類的分布,推測其可能是藏雪雞、暗腹雪雞(Tetraogallushimalayensis)或石雞(Alectorischukar)。本研究結果表明,雪豹與藏雪雞空間關聯性(0.47)強,捕食關系可能促使它們共同出現在相同空間的概率增加。雪豹與雪鶉、綠尾虹雉之間的空間關聯可能由類似的原因導致。基于以上分析,獲得雪豹與各直接關聯物種的種間生態學關系,如圖2所示。
4.4 群落抗干擾能力
圖2群落中,食草動物有巖羊、喜馬拉雅旱獺和藏鼠兔,雜食動物有綠尾虹雉、紅喉雉鶉、白馬雞、雪鶉、血雉、藏雪雞和豬獾,食肉動物有雪豹、赤狐、石貂和香鼬。每個營養級上都有2個以上的物種。當同一營養級中一個物種出現種群密度下降時,其他物種可以替補其生態學功能,由此為群落提供應對外來沖擊的韌性。如:當雪豹因環境動蕩出現種群密度下降時,同樣處于捕食者的赤狐可以替代雪豹控制食草動物數量;反之亦然。通過這種機制,食草動物種群數量可以受到持續調控,避免草場被過度利用,群落得以有效維持。
4.5 雪豹在群落中的角色
雪豹是群落的頂級捕食者,被保護生物學家視為高海拔旗艦物種[27]。本研究結果(圖2)表明,雪豹和赤狐是群落中重要捕食物種,像2只手控制著群落中的其他物種。雪豹捕食藏馬雞、香鼬、石貂、喜馬拉雅旱獺、綠尾虹雉、巖羊和雪鶉,共7種,占群落物種數50%,其中,石貂(0.59)、喜馬拉雅旱獺(0.57)、巖羊(0.63)和雪鶉(0.59)為主要食物物種。赤狐捕食白馬雞、紅喉雉鶉、豬獾、血雉、綠尾虹雉、巖羊、石貂和喜馬拉雅旱獺,共8種,占群落物種數57%;從相關系數看,白馬雞(0.53)、紅喉雉鶉(0.60)、血雉(0.67)、巖羊(0.55)和喜馬拉雅旱獺(0.54)為主要食物物種。二者共同的食物物種有:巖羊、喜馬拉雅旱獺、石貂、綠尾虹雉。從獵物物種總數來看,赤狐比雪豹多1種。但值得注意的是,從體型上看,赤狐體重5—7 kg不太可能捕食成年巖羊(60—75 kg)[15],在群落中僅有雪豹能對成體巖羊起到控制作用。因此,作為頂級捕食者雪豹在高海拔群落中具有不可替代的作用,應該加強保護與研究。
綜上所述,本研究認為臥龍國家級自然保護區雪豹地棲動物群落作為一個實體客觀存在,群落中各營養級都有2個以上的物種,同級物種的生態角色可以相互替換,表明臥龍雪豹群落是一個健康的群落,具備繼續演化的能力。
致謝:感謝四川省林業和草原局古曉東的大力支持;臥龍國家級自然保護區胡強、王茂麟、林紅強的傾力協助;劍橋大學李夢燁女士幫助修改英文摘要!
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:30
當代陜西(2021年17期)2021-11-06 03:21:36
當代陜西(2019年15期)2019-09-02 01:52:00
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
學苑創造·A版(2018年11期)2018-02-01 06:29:20
讀者(2017年5期)2017-02-15 18:04:18
