哺乳動物毛囊周期性生長調控機制的研究進展
2021-08-11 02:58:28劉羅蘭馮登偵李新海
生命科學研究 2021年3期
張 穩,劉羅蘭,馮登偵,李新海
(寧夏大學農學院,中國寧夏銀川750021)
毛發是哺乳動物皮膚的衍生物,由凹陷于皮膚真皮層的毛囊產生,具有冷熱隔離、物理保護、社交互作及感官感應等諸多生物學功能[1],是毛用動物的重要經濟性狀,其產量和質量直接影響著養殖戶的經濟效益。毛囊作為產生毛發的基本單位,其周期性生長對毛纖維的發生、生長、脫落及再生具有關鍵作用。現已發現毛囊的生長發育受內分泌激素褪黑素以及一系列基因和信號通路如Wnt/β-catenin、骨形態發生蛋白質(bone morphogenetic protein,BMP)、成纖維細胞生長因子5(fibroblast growth factor 5,FGF5)的調控。隨著高通量測序技術的應用普及,許多研究轉向非編碼RNA對毛囊周期性生長的調控,目前已有一些關于微RNA(microRNA,miRNA)及長鏈非編碼RNA(long non-coding RNA,lncRNA)在毛囊生長期、退行期、休止期差異表達及靶基因調控的研究。此外,近期研究顯示毛囊的生長發育及周期性活動還涉及糖酵解酶、膠原蛋白、干細胞及免疫T細胞。本文簡要描述了毛囊的周期性生長活動,論述了褪黑素在毛囊生長發育中的作用以及調控毛囊周期性生長的基因及信號通路、非編碼RNA和其他毛囊生長發育相關的蛋白質/細胞,以期為全面揭示毛囊周期性生長的調控機制、提高毛用經濟動物絨毛的產量和質量以及治療人類毛囊疾病等提供參考。
1 毛囊的周期性生長
毛囊是哺乳動物皮膚特殊的附屬器官,位于皮膚的真皮層。毛囊發生于哺乳動物胚胎期,主要依賴于真皮細胞和上皮細胞之間傳遞的一系列信號分子[2~3]。不同哺乳動物的毛囊形態千差萬別,但都具有毛球、毛干、內根鞘、外根鞘等基本結構(圖1)[4]。依照形成時間與結構功能特點,毛囊可分為初級毛囊(primary follicle,PF)和次級毛囊(secondary follicle,SF)。初級毛囊由上皮細胞和真皮層細胞分化發育形成,次級毛囊由發育中的初級毛囊分化而來[5~6]。
哺乳動物毛囊呈周期性生長,毛囊循環經歷著生長期、退行期、休止期,各個時期的形態變化存在很大差別[7](圖1),這一重要生理特征與毛囊干細胞(hair follicle stem cell,HFSC)周期性的生長、增殖、分化及真皮乳頭細胞與上皮細胞之間的信號互作密切相關[8~9]。以內蒙古絨山羊為例,其初級毛囊周期性變化不明顯,但次級毛囊具有明顯的季節性周期變化。在生長期,毛囊底端真皮乳頭細胞發出的增殖信號誘導毛母質細胞快速增殖分化并向真皮層延伸形成毛干和內根鞘[10~11],毛囊形態結構處于動態平衡。隨后,真皮區域的毛囊可再生部位的毛母質細胞增殖減少,毛球發生萎縮,毛干停止分化并形成杵狀毛發,進入退行期。接著,毛囊進入細胞活動程度最低的休止期[12]。在休止期末期,毛球開始再生,HFSC分化并進行增殖,毛囊再生部分進行重建,進入下一輪的生長期。此時杵狀毛干會因新毛干的產生被頂出皮膚表層,直至脫落。不同哺乳動物毛囊周期性生長的3個時期的持續時間長短不同,表1對幾種哺乳動物毛囊生長不同時期的持續時間進行了概括[13~16]。

表1 不同哺乳動物毛囊生長周期不同階段的持續時間Table 1 Duration of different stages of hair follicles in different mammals
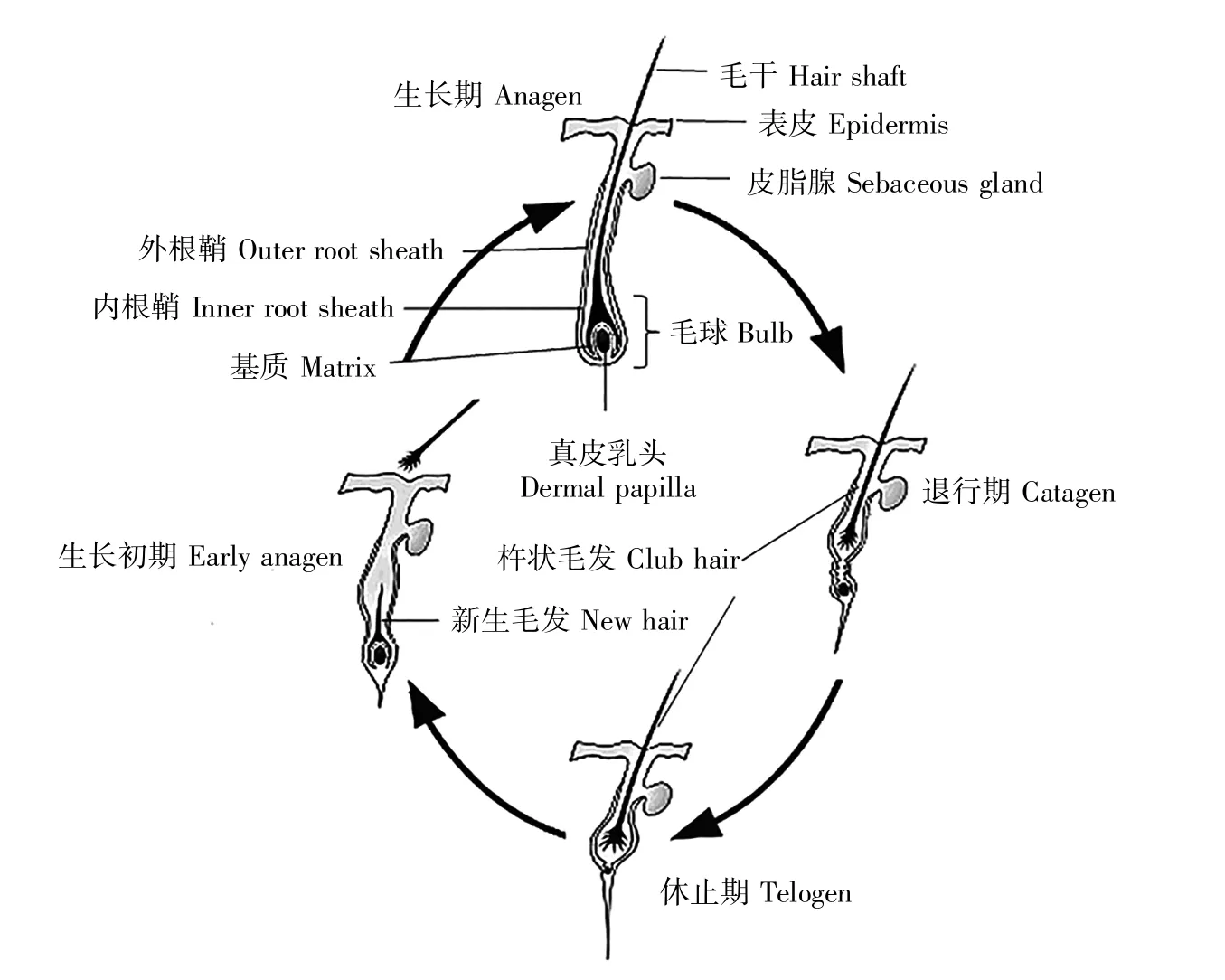
圖1 毛囊組織結構及生長周期[4,7]Fig.1 Structure and periodic growth of hair follicles[4,7]
2 毛囊周期性生長的調控機制
毛囊的周期性生長是在一個極其復雜的調控網絡下進行的,由一系列調控因子共同參與,包括褪黑素、毛囊生長發育相關的基因及信號通路、非編碼RNA和毛囊細胞活動相關的一些蛋白質/細胞等。
2.1 褪黑素與毛囊生長發育
褪黑素是主要由松果體和視網膜分泌的一種高度保守的吲哚類激素[17],在哺乳動物皮膚中也能合成和代謝[18~20],廣泛存在于多種動植物組織和器官內,通過與其受體MTR(melatonin receptor)結合發揮著重要的生物學作用。研究發現,褪黑素在調節不同哺乳動物皮毛生長中發揮重要作用(表2)[21~26]。早在1969年,Rust等[21]在使用外源褪黑素促使白鼬過早成熟、誘導蛻皮時就發現了褪黑素在動物季節性換毛中的重要作用。Allain等[27]在給成年雌貂埋植或者口服褪黑素時發現,其毛囊數目和毛產量大大增加,人們在獺兔[28]和內蒙古絨山羊[29~30]的研究中得到類似結果。Frank等[31]發現,經褪黑素治療的脫毛博美犬在數月后出現了毛發再生和增多,說明褪黑素能夠促進哺乳動物毛發生長、毛囊密度增加以及誘導毛發再生。針對濟寧青山羊的研究報道,MTR-1A在大猾皮中的表達水平極顯著高于其在胚胎期、小猾皮、成年羊背部皮膚中的表達水平,說明MTR-1A在促進初級毛囊毛纖維生長和次級毛囊發育起始階段起正向作用[32]。由此可見,褪黑素在多種哺乳動物毛皮生長中具有正向調控作用,將其科學應用到毛皮用經濟動物的養殖中,有可能帶來可觀的經濟收益。

表2 褪黑素在不同哺乳動物皮毛生長中的作用Table 2 The role of melatonin in the fur growth of different mammals
研究報道,埋植褪黑素能夠縮短內蒙古絨山羊次級毛囊的生長周期,促進絨毛生長,提高絨毛產量,并推遲脫絨時間,但對初級毛囊生長周期和粗毛長度無顯著影響[33],這與常子麗[34]在內蒙古絨山羊中的研究結果以及黃冬維等[35]在長毛兔上的研究結果具有一致性。此外,埋植褪黑素使Wnt10b在次級毛囊外根鞘和連接組織鞘中的表達提前1~2個月,進而使新絨毛的生長提前[33]。付紹印等[36]發現,在絨毛興盛前期(6月份)埋植褪黑素下調了 miR-203、miR-205、miR-96、miR-183、miR-199a的表達,并提前誘發了絨毛的二次生長,其中miR-96的下調能夠提高毛囊細胞活性,抑制毛囊細胞凋亡,推測褪黑素與轉錄后調控因子miRNAs相互聯系,共同作用于皮膚毛囊生長周期,影響絨毛生長。Notch信號通路是已知的能夠調節HFSC生長分化和增殖的通路,該通路受到Numb、Deltex等基因的調控。張寧寧[37]發現,不同濃度的褪黑素均能影響HFSC以及Notch信號通路相關基因Notch1、Hes-1、Numb和Deltex的表達趨勢,推測褪黑素可能與Notch通路相關基因相互作用影響HFSC的活動,從而調節毛囊生長周期。此外,Ge等[38]通過對褪黑素處理的陜北絨山羊背部皮膚次生毛囊干細胞(gsHFSC)進行lncRNA測序發現,褪黑素的介導影響了黏著斑和胞外基質受體通路相關的基因,促進了毛囊的生長;其中lncRNA TCONS_00255106和TCONS_00206163分別靶向影響了毛囊相關基因COL4A2和THBS2的表達趨勢,推測褪黑素能夠影響毛囊形態發生相關基因,并通過lncRNA調控靶基因表達,影響HFSC自我更新。Jin等[39]通過體外培養不同濃度、不同時間褪黑素處理的遼寧絨山羊成纖維細胞以及測序分析各組細胞中lncRNA和mRNA的差異表達發現,0.2 g/L褪黑素處理72 h對羊絨生長最有利,且褪黑素通過上調lncRNAMTC的表達促進成纖維細胞增殖,其過表達激活了具有相同促進作用的核因子κB(nuclear factor-κB,NF-κB)信號,推測褪黑素通過提高lncRNAMTC的表達激活NF-κB信號,從而控制毛囊生長發育和毛發生長。由此可知,褪黑素并不是單獨影響毛發生長的,它通過與相關調控因子互作,共同調節動物毛發生長,但其作用機制仍不明確,有待進一步研究。總的來講,多角度聯合探索褪黑素在動物皮毛中的作用機制有助于更好地理解影響動物毛皮品質的因素,為生產實踐提供理論基礎。
2.2 毛囊生長發育相關的信號通路
近年來,關于毛囊生長發育調控信號通路的研究越來越多,尤其是 Wnt/β-catenin、BMP和FGF5信號通路。Huelsken等[40]發現,表皮中βcatenin基因發生突變會導致小鼠毛囊基板形成受阻;即使在毛囊初次形成后敲除β-catenin基因,在一次生長周期后毛囊仍會出現生長缺失。與該結果類似,Tao等[41]發現敲除小鼠毛囊真皮乳頭中的β-catenin基因導致毛母質細胞凋亡,毛囊提前進入退行期,且激活真皮層的Wnt/βcatenin信號能夠調控HFSC的毛發誘導能力并誘發皮膚的纖維化,與下游的BMP及FGF等信號通路交互調控異位毛囊的發生。馬森[42]發現,Wnt/β-catenin通路的上游基因 Wnt3、β-catenin和Tef3等的表達在陜北絨山羊毛囊生長前期緩慢上調,在生長旺盛期達到最高,并在退行期至休止期逐漸下降。這些研究說明,Wnt/β-catenin在毛囊生長過程中起到正向調控作用,能夠促進毛囊生長初期基板形成、激活HFSC,使毛囊進入生長期(毛囊再生階段)。相關研究顯示,Wnt/β-catenin下游信號中表達于表皮層的BMP2和真皮層的BMP4、BMP7在毛囊形成初期抑制基板的形成[43],且當BPM2基因表達于小鼠皮下脂肪和真皮層時,其在休止期早期的表達水平較高,而末期沒有表達[44]。Song等[45]通過qRT-PCR和Western-blot實驗發現,BMP2基因和BMPR-IA(BMPⅠ型受體)基因在牦牛皮膚毛囊休止期的表達量水平高,而BMP的拮抗劑Noggin在生長期前期和中期的表達水平顯著高于休止期。Noggin通過抑制BMP信號促進毛囊生長,當位于毛囊真皮凝聚物中的Noggin過量表達時,真皮層中BMP抑制Wnt信號的抑制環境消失,從而促進HFSC的活化,使毛囊進入下一輪周期的生長期[45~46]。這些結果表明,BMP和Noggin在毛囊周期中的表達具有時空差異性,可作為毛囊休止期的標記信號。需要指出的是,相關研究對0~6周齡不同時期水貂皮膚組織內的BMP2表達水平進行了分析,發現在水貂不同周齡皮膚中BMP2表達較為穩定,推測BMP2對0~6周齡水貂毛囊發育無直接影響[47]。這說明BMP2在不同物種毛囊發育時期中的作用存在差異。同樣能夠抑制毛囊生長的FGF5蛋白是迄今已知的最強勁的促進毛囊從生長期向退行期轉化并進入休止期的調控因子。Higgins等[48]利用重組FGF5基因處理人皮膚毛囊,發現其在2~4 d后提前進入退行期,而對照組毛囊則在5~7 d進入退行期,提示FGF5基因能夠啟動毛囊退行期,抑制毛發生長。研究顯示,FGF5基因突變會引起貓和狗的毛發長度發生改變[49~50],這與梁東[51]發現的水貂絨毛長度與FGF5基因表達水平呈負相關的結果一致。FGF5基因過表達會增加BMP4 mRNA的表達,從而抑制Noggin表達,導致毛發生長受到抑制[52],這說明FGF5與BMP信號互作,共同抑制毛囊的生長期活動。有意思的是,趙德超等[53]研究發現埋植褪黑素會提高FGF5基因的表達,導致絨山羊毛囊生長期提前,促進絨山羊羊絨的生長,這與He[52]、Higgins[48]等的研究結果不一致,說明FGF5在不同物種和不同處理條件下對毛囊的生長調控作用存在差異,其具體作用機制需要進一步探究。
除此之外,TGF-β(transforming growth factorβ)、Notch、SHH(sonic hedgehog)等信號通路也都在毛囊的生長發育過程中起重要作用。各信號通路相互協調,共同參與毛囊的生長發育。這些相關研究可為人為調控毛用經濟動物的毛囊生長周期從而提高絨、毛產量和質量提供理論與實驗基礎。
2.2 非編碼RNA對毛囊生長發育的調控
2.2.1 miRNA調控毛囊生長發育
miRNA是一類長度為20~24 nt的內源性非編碼RNA分子,在多種真核生物中均有分布[54]。已知的miRNA生物學功能有調控胚胎發生、器官分化、個體發育[55]以及多個信號通路、代謝過程[56]等。近年來研究發現,miRNA參與哺乳動物毛囊的生長發育,而且關于miRNA參與小鼠、綿羊、山羊等哺乳動物皮膚毛囊發育和周期性生長的報道越來越多。
Yi等[57]首次描述了miRNA在皮膚中的特征,并證明其在胚胎發育過程中參與單層上皮多能干細胞分化形成表皮及其附屬物的生物學過程。研究報道,miRNA成熟所需的酶基因(Dicer基因)被敲除后,發育中的毛囊細胞發生非正常的凸起,出現毛囊形態的異常[57],說明miRNA參與構建毛囊細胞形態。Liu等[58]發現,miR-195可能通過靶向調控Wnt/β-catenin通路中的SMAD2(mothers against decapentaplegic homolog 2)和FZD6(Frizzled 6)基因而在毛囊發生初期促進毛囊生長。相關研究利用測序技術分析了內蒙古絨山羊皮膚毛囊3個生長時期的miRNA,得到生長期、退行期、休止期特異表達的miRNA分別為15個、8個和7個,說明差異表達的miRNA可能通過調節毛囊細胞活動影響毛囊周期,其在毛囊時期的轉換過程中可能發揮重要作用[59]。這有利于篩選鑒定毛囊生長階段的miRNA分子標記。同時,通過人為干預相關miRNA調控靶基因的表達調節毛囊生長進程,可在一定程度上避免季節因素對毛囊生長周期的影響,從而得到高產高質的絨、毛。
2.2.2 lncRNA調控毛囊生長發育
lncRNA是一類長度超過200 nt且本身不具有編碼能力的非編碼RNA[60]。研究發現,lncRNA在表觀遺傳水平(如基因組印記、染色質修飾、劑量補償效應)、轉錄及轉錄后水平、核內運輸等重要過程發揮調控作用[61~63]。隨著高通量測序技術的發展,lncRNA在毛囊生長發育中的研究也已獲得一定成果,其中關于絨山羊皮膚毛囊的研究頗多。
研究證明,相關lncRNA在絨山羊毛囊周期性生長過程的表達具有時間特異性。針對陜北絨山羊皮膚毛囊周期性生長的lncRNA的研究發現,198個lncRNA在生長期和休止期差異表達。其中,lnc_00092、lnc_000183、lnc_000406 和 lnc_000559為休止期特異表達的lncRNA;lnc_000173為生長期特異表達的lncRNA,可作為生長時期鑒定的標記lncRNA[64]。Bai等[65]通過研究遼寧絨山羊次級毛囊生長期和休止期的轉錄模式發現,6種已知的lncRNA(lncRNA-599618,-599556,-599554,-599547,-599531和-599509)在生長初期的表達顯著高于休止期,且均在毛囊生長期的Wnt信號通路中高表達,而4種已知的lncRNA(lncRNA-599528,-599518,-599511和-599497)在休止期的表達顯著高于生長期,這說明生長期高表達的6種lncRNA可能與Wnt信號協同作用,促進毛囊基板形成及HFSC的激活、分化和增殖,從而促進絨毛纖維生長,而休止期高表達的lncRNA可能參與抑制HFSC的分化,從而使毛囊處于休眠狀態。這些結果為進一步揭示lncRNA在其他哺乳動物皮膚毛囊中的周期轉換機制提供了借鑒。
研究發現,lncRNA可通過調節下游靶基因的表達調控毛囊的生長發育。Zhou等[66]利用多組學方法,通過順式和反式調節研究了lncRNA的靶基因,發現角蛋白基因和角蛋白相關基因KRT38、KRT4、KRTAP15-1、KRTAP13-1及 KRTAP3-1在毛囊生長期的表達高于退行期,而KRT2、KRTDAP、KRT77和KRT80則相反,推測lncRNA通過靶向調控毛囊相關角蛋白影響毛囊生長周期進程。Si等[67]通過細胞轉染實驗發現,PlncRNA-1轉染會顯著促進HFSC的細胞周期、增殖和分化,且引起 TGF-β1、Wnt3和 β-catenin基因表達的上調。此外,TGF-β1的抑制劑會導致HFSC中Wnt和β-catenin的表達水平降低,并抑制HF-SC的細胞周期,降低由PlncRNA-1轉染誘導的細胞周期的加速。這一結果說明PlncRNA-1可通過TGF-β1基因介導的Wnt/β-catenin信號通路調節HFSC的細胞周期,影響其增殖和分化。Cai等[68]發現,lncRNA5322轉染后通過上調PI3K/Akt信號促進miR-21表達,并誘導HFSC的增殖和分化,其中miR-21是lncRNA5322在HFSC分化時的直接靶標,提示miRNA與lncRNA協同作用促進毛囊生長。
此外,上文也提到lncRNA能通過調控基因的靶向表達,或通過褪黑素介導,調節毛囊生長周期的活動。而且,有研究證實lncRNA在毛囊真皮乳頭細胞的分化、真皮凝聚物和毛囊基板的形成及HFSC的增殖與分化中有重要的作用,是次級毛囊發生時的必需因素[69]。由此可見,lncRNA在毛囊生長發育中發揮重要作用,并通過參與調控毛囊生長相關基因的表達、與多種信號通路互作以及褪黑素的介導發揮生物學功能。因此,通過高通量測序技術、生物信息學手段及分子實驗技術分析lncRNA在毛囊生長發育過程中的作用機制并構建其與其他相關調控的互作網絡,對了解毛囊生長發育的分子機理尤為重要。
2.3 毛囊生長發育相關的其他蛋白質/細胞
2.3.1 ⅩⅦ型膠原蛋白
ⅩⅦ型膠原蛋白是半橋粒的主要成分之一,由COL17A1基因編碼,因此也被稱為COL17A1蛋白。已有研究證實,COL17A1蛋白參與維持HFSC的自我更新[70]。Matsumura等[71]以小鼠為研究對象發現,過表達COL17A1蛋白的年老小鼠僅有較少毛囊發生萎縮,且脫毛現象減輕,HFSC活性高,而正常表達COL17A1蛋白的年老小鼠的毛囊大面積萎縮且脫毛嚴重,HFSC活性低,同時皮膚彈性變差。這說明在COL17A1蛋白的介導下毛囊中HFSC的老化情況得到抑制,從而有助于減少脫發和皮膚衰老。這一發現對治療人類脫發和延緩皮膚衰老具有重要意義。Liu等[72]發現COL17A1蛋白表達水平的高低會影響上皮干細胞的生存能力,高表達COL17A1的表皮干細胞競爭力強,并可加快毛囊的生長發育及生長周期的更替。這些研究說明COL17A1蛋白的表達會影響毛囊細胞的生存能力,從而影響毛囊周期進程。這為人工參與并調控人以及皮毛用動物(如水貂、長毛兔、絨山羊)的毛囊生長周期提供了新的實驗方向,可能有助于解決人類脫發問題以及獲得高產高質的皮毛商品。
2.3.2 乳酸脫氫酶
乳酸脫氫酶(lactate dehydrogenase,LDH)是一種能夠催化NADH依賴性丙酮酸還原成乳酸和NAD+依賴性乳酸氧化成丙酮酸的糖酵解酶,其能夠促進HFSC活化,而HFSC的活化和增殖是促進毛囊周期更新的先決條件[73~74]。Flores等[74]以小鼠為研究對象,通過酶比色法發現,LDH在HFSC中的活性明顯比在其他皮膚細胞中高,且在HFSC大量富集。當LDH缺乏時,HFSC無法被激活;相反,當促進乳酸產生時,HFSC被快速激活,毛囊進入新的循環周期,推測乳酸的產生是HFSC激活的關鍵因素,影響毛囊周期的更替。該結果有助于進一步探究HFSC活動對毛囊周期性生長的調控機制。
2.3.3 調節性T細胞
調節性T細胞(regulatory T cells,Tregs)是一類與機體免疫抑制相關的T細胞亞群,在維持機體免疫穩態、腫瘤免疫、移植耐受等方面具有重要的調節功能[75]。小鼠和人類皮膚組織中都含有大量的Tregs[76~77],且皮膚中的Tregs在毛囊附近富集[76,78],能夠促進毛囊休止期向生長期的轉換。Ali等[79]通過RNA-seq發現,缺乏Tregs的小鼠HFSC中與細胞增殖相關的基因比Tregs豐富的對照組明顯減少,這說明Tregs可通過促進細胞增殖基因的表達加快HFSC的激活和分化,使得毛囊生長期提前,從而提高絨、毛的產量。以上結果從免疫T細胞的角度揭示了HFSC的活動規律,但其與相關基因的互作機制仍需進一步探究。
上述結果說明毛囊的周期性生長過程涉及到許多蛋白質/細胞,它們通過直接或間接的相互作用影響毛囊細胞的生長活動,尤其是HFSC,從而調控毛囊的生長周期。雖然一些關于毛囊周期性生長的新研究在糖酵解酶和免疫細胞方面已經有所突破,但它們大多集中在對HFSC活動的影響上,具體的調控機制有待進一步探究。另外,目前相關研究多是在小鼠中進行的,考慮到物種間毛囊生長發育的差異性,在人、絨山羊、兔子、水貂等哺乳動物中的研究仍需要繼續推進。
3 結語與展望
毛發作為哺乳動物皮膚的重要衍生物,是羊、兔、水貂等哺乳動物的重要經濟性狀,具有重要的研究意義。毛囊作為產生毛發的微小器官,其周期性生長主要受到褪黑素的節律調節。此外,miRNA、lncRNA、膠原蛋白、糖酵解酶及T細胞等眾多因素也參與其調控,各因子之間通過促進、抑制或間接靶向等作用調控毛囊生長發育。近年來,褪黑素、相關基因及信號通路在哺乳動物毛囊周期性生長上的研究成果頗豐,但從非編碼RNA靶向基因調控、非編碼RNA-mRNA調控網絡構建、代謝途徑和免疫反應角度研究毛囊生長發育的報道較少,其具體調控機制仍需進一步挖掘。由于HFSC是毛囊生長周期更替的關鍵因素,對毛囊再生意義非凡,因此深入研究HFSC的激活機制有重要意義。其中,從基因互作、乳酸代謝途徑及免疫反應等角度全方位揭示哺乳動物HFSC的活化機制亟待進行。
高通量測序技術的飛速發展為進一步解析哺乳動物毛囊生長發育的分子調控機制提供了有利工具。利用轉錄組技術進一步深入探究毛囊周期性生長轉錄水平上的分子調控機制;結合多組學技術分析并構建轉錄后水平調控模式;通過細胞水平和活體驗證來全方位解析毛囊生長發育規律,并將其應用到生產實踐中,可為進一步改善兔、絨山羊、水貂等毛用哺乳動物的毛皮品質提供科學的理論指導和研究方向。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
小讀者(2021年2期)2021-03-29 05:03:48
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
中國生殖健康(2019年3期)2019-02-01 06:12:26
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
