青龍湖浮游植物群落與環(huán)境因子相關(guān)性分析
2021-08-11 15:22:44劉川閆金霞劉家樂(lè)李鼎馬玉露王榮欣成慶利陸建紅
人民黃河 2021年7期
劉川 閆金霞 劉家樂(lè) 李鼎 馬玉露 王榮欣 成慶利 陸建紅

摘 要:污染物匯入和閘壩攔截等人為干擾影響湖泊水量和水質(zhì),進(jìn)而影響湖泊生物群落的結(jié)構(gòu)和組成。為了解青龍湖浮游植物群落狀況及其環(huán)境驅(qū)動(dòng)因子,于2018年10月、12月,2019年4月、7月對(duì)青龍湖的水質(zhì)及浮游植物進(jìn)行采樣調(diào)查,利用藻類(lèi)生物學(xué)指標(biāo)評(píng)價(jià)水體營(yíng)養(yǎng)狀況,并對(duì)浮游植物和環(huán)境因子進(jìn)行冗余分析(RDA),結(jié)果表明:調(diào)查期間共鑒定出浮游植物8門(mén)53屬,浮游植物的密度為134.20萬(wàn)~283.16萬(wàn)個(gè)/L,生物量為2.48~8.09 mg/L;春季綠藻-硅藻為優(yōu)勢(shì)種,夏季轉(zhuǎn)以綠藻為主,秋季以藍(lán)藻為主,冬季以硅藻為主;水體大部分時(shí)期為中-富營(yíng)養(yǎng)型;水溫、pH值、總磷、氨氮、CODMn和電導(dǎo)率是影響浮游植物生物量的主要環(huán)境因子。
關(guān)鍵詞:浮游植物;環(huán)境因子;冗余分析;青龍湖
中圖分類(lèi)號(hào):TV211.1+1;X824 文獻(xiàn)標(biāo)志碼:A
doi:10.3969/j.issn.1000-1379.2021.07.020
引用格式:劉川,閆金霞,劉家樂(lè),等.青龍湖浮游植物群落與環(huán)境因子相關(guān)性分析[J].人民黃河,2021,43(7):104-109.
Abstract:Pollution inflows and dam interception affect the lake water quality and then influence the structure and composition of aquatic organisms. In order to explore the phytoplankton community and its environmental driving factors in Qinglong Lake, the composition of phytoplankton community and environmental factors had been investigated in October and December 2018 and April and July 2019. The algal biological indices were used to evaluate the water quality of Qinglong Lake. Meanwhile, redundancy analysis (RDA) was performed between phytoplankton and environmental factors. The results show that 53 genera belonging to 8 phyla of phytoplankton are identified. The density of phytoplankton is from 1.342 0~2.831 6 million cells/L and the biomass of phytoplankton is from 2.48 to 8.09 mg/L respectively. In spring, the phytoplankton communities are dominated by green algae and diatoms, then it is dominated by green algae in summer and followed by cyanobacteria and diatoms in autumn and winter. Water quality assessment results indicate that the water body is in a medium-eutrophication condition. RDA makes clear that water temperature, pH, total phosphorus, ammonia nitrogen, CODMn and conductivity are the environmental factors affecting phytoplankton community significantly.
Key words: phytoplankton; environmental factors; RDA; Qinglong Lake
浮游植物是水生態(tài)系統(tǒng)中的初級(jí)生產(chǎn)者,在物質(zhì)循環(huán)和能量流動(dòng)中起著重要作用[1]。浮游植物對(duì)于環(huán)境因素(如溫度、pH值、營(yíng)養(yǎng)鹽等)的變化十分敏感[2],能夠準(zhǔn)確反映水質(zhì)的變化,同時(shí)其生物群落結(jié)構(gòu)的變化也會(huì)反作用于水質(zhì)[3]。目前,浮游植物的密度、生物量及多樣性指數(shù)等生物學(xué)指標(biāo)已廣泛應(yīng)用于各類(lèi)水體的水質(zhì)評(píng)價(jià)[4-6]。
青龍湖位于河南省新鄉(xiāng)市封丘縣和長(zhǎng)垣縣境內(nèi),為200多a前黃河決堤沖刷而成的沼澤型濕地,水域面積約59.4 hm2,水深較淺,最大水深約3 m,是新鄉(xiāng)黃河濕地鳥(niǎo)類(lèi)國(guó)家級(jí)自然保護(hù)區(qū)內(nèi)少數(shù)常年集水區(qū)之一。該濕地動(dòng)植物資源豐富,是許多珍稀禽鳥(niǎo)的繁殖棲息地,也是候鳥(niǎo)的越冬北界[8]。近年來(lái)隨著經(jīng)濟(jì)的快速發(fā)展,人類(lèi)活動(dòng)對(duì)環(huán)境的影響加劇,青龍湖水域面積減小、水質(zhì)狀況及生態(tài)功能惡化,保護(hù)并恢復(fù)其生態(tài)功能迫在眉睫。因此,筆者調(diào)查青龍湖不同時(shí)空浮游植物的群落組成及水質(zhì)狀況,通過(guò)藻類(lèi)生物學(xué)指標(biāo)及其冗余分析,探討浮游植物與水質(zhì)因子間的關(guān)系,旨在為青龍湖水體富營(yíng)養(yǎng)化防治和生態(tài)保護(hù)提供依據(jù)。
1 材料與方法
1.1 采樣點(diǎn)分布及采樣時(shí)間
根據(jù)生境特點(diǎn)的不同將青龍湖分為A區(qū)北部淺水區(qū)(挺水植物占優(yōu)勢(shì))、B區(qū)中心敞水區(qū)(浮游植物占優(yōu)勢(shì))[7]。考慮人類(lèi)活動(dòng)影響的大小、距排污口的遠(yuǎn)近及水生生物特征等因素在A(yíng)、B兩區(qū)分別設(shè)置4個(gè)采樣點(diǎn),于2018年10月(秋季)、12月(冬季),2019年4月(春季)、7月(夏季)每月下旬進(jìn)行采樣,每次采樣安排在天氣晴朗的上午。
1.2 浮游植物的采集及鑒定
使用2.5 L有機(jī)玻璃采水器于水下0.5 m處采集水樣,裝于1 L高密度聚乙烯瓶中。使用冰袋冷藏送回實(shí)驗(yàn)室后,利用FlowCAM流式細(xì)胞成像儀對(duì)藻類(lèi)的形態(tài)進(jìn)行成像,同時(shí)測(cè)定浮游植物密度及等效球體直徑。若不能立即檢測(cè),則加入4%甲醛溶液固定后于4 ℃下保存,以待檢測(cè)。浮游植物的鑒定依據(jù)水生生物圖譜、《中國(guó)淡水藻類(lèi)——系統(tǒng)、分類(lèi)及生態(tài)》[8]等進(jìn)行。
1.3 水體理化因子的測(cè)定
共分析透明度(SD)、水溫(WT)、總?cè)芙庑怨腆w(TDS)、電導(dǎo)率(COND)、氧化還原電位(ORP)、pH值、溶解氧(DO)、高錳酸鹽指數(shù)(CODMn)、總磷(TP)、氨氮(NH+4-N)10項(xiàng)水質(zhì)指標(biāo),其中:透明度采用塞氏盤(pán)測(cè)定;WT、COND、TDS、ORP、pH值等采用UltrameterⅡTM6PFCE多功能水質(zhì)儀現(xiàn)場(chǎng)測(cè)定;DO采用雷磁JPBJ-608型便攜式溶解氧分析儀現(xiàn)場(chǎng)測(cè)定;使用2.5 L有機(jī)玻璃采水器另取水樣貯存于1 L高密度聚乙烯瓶中,利用冰袋冷藏后送回實(shí)驗(yàn)室,根據(jù)《水和廢水監(jiān)測(cè)分析方法》[9]測(cè)定TP、NH+4-N、CODMn等指標(biāo)。
1.4 數(shù)據(jù)處理與分析
根據(jù)浮游植物密度、Shannon-Wiener多樣性指數(shù)(H′)、Pielou均勻度指數(shù)(J)和優(yōu)勢(shì)度(Y)對(duì)青龍湖浮游植物群落結(jié)構(gòu)特征進(jìn)行計(jì)算分析,各指標(biāo)計(jì)算公式:
式中:Ni、N分別為第i個(gè)屬浮游植物密度、浮游植物總密度;S為樣品中浮游植物總屬數(shù);Pi=Ni/N,為第i個(gè)屬浮游植物密度占總密度的比例,可通過(guò)流式細(xì)胞成像儀直接讀取;fi為第i個(gè)屬出現(xiàn)的頻率;Y為優(yōu)勢(shì)度,Y≥0.02即為優(yōu)勢(shì)屬。
浮游植物生物量根據(jù)《內(nèi)陸水域漁業(yè)自然資源調(diào)查手冊(cè)》[10]相關(guān)方法計(jì)算。水體營(yíng)養(yǎng)狀況的藻類(lèi)生物學(xué)評(píng)價(jià)標(biāo)準(zhǔn)[11-12]見(jiàn)表1。
利用Origin 2017制圖,使用SPSS 24.0對(duì)多樣性指數(shù)和環(huán)境因子進(jìn)行Pearson相關(guān)性分析,使用Canoco for Windows 4.5分析浮游植物生物量和環(huán)境因子之間的關(guān)系。分析前對(duì)環(huán)境因子數(shù)據(jù)和浮游植物群落數(shù)據(jù)進(jìn)行l(wèi)n(x+1)轉(zhuǎn)換(x為原始數(shù)據(jù)),使其趨于正態(tài)分布;并且先對(duì)浮游植物物種信息進(jìn)行去趨勢(shì)對(duì)應(yīng)分析(DCA),若最大梯度軸長(zhǎng)度小于3,則進(jìn)行冗余分析(RDA)。通過(guò)軟件自動(dòng)篩選和蒙特卡洛檢驗(yàn)剔除膨脹因子大于20的環(huán)境因子,避免環(huán)境因子間存在較強(qiáng)的多重共線(xiàn)性,影響排序結(jié)果。利用CanoDraw繪制物種與環(huán)境因子的雙序圖。
2 結(jié)果與分析
2.1 浮游植物種類(lèi)組成與優(yōu)勢(shì)屬
調(diào)查期間,青龍湖共檢出8門(mén)53屬浮游植物。其中:綠藻門(mén)(Chlorophyta)23屬,隱藻門(mén)(Cryptophyta)3屬,硅藻門(mén)(Bacillariophyta)14屬,裸藻門(mén)(Euglenophyta)4屬,黃藻門(mén)(Xanthophyta)1屬,金藻門(mén)(Chrysophyta)2屬,甲藻門(mén)(Pyrrophyta)2屬,藍(lán)藻門(mén)(Cyanophyta)4屬。綠藻全年平均密度約占總密度的35.9%,硅藻、藍(lán)藻次之,分別約占24.5%、23.5%。浮游植物季節(jié)分布情況見(jiàn)表2,從不同季節(jié)分布來(lái)看,春季檢出的浮游植物種類(lèi)最多,冬季最少;夏季、冬季浮游植物群落分別以綠藻、硅藻為主,春、秋兩季綠藻和硅藻種類(lèi)均較多。不同季節(jié),青龍湖浮游植物優(yōu)勢(shì)屬統(tǒng)計(jì)結(jié)果見(jiàn)表3(+表示為優(yōu)勢(shì)種),共檢出6門(mén)16屬優(yōu)勢(shì)浮游植物。春季有5門(mén)10屬,夏季4門(mén)7屬,秋季4門(mén)8屬,冬季4門(mén)5屬;衣藻在全年均有優(yōu)勢(shì),小環(huán)藻和桿藻等硅藻在溫度較低時(shí)占有優(yōu)勢(shì),尖頭藻和念珠藻在夏、秋季形成優(yōu)勢(shì)。
2.2 浮游植物密度及生物量
調(diào)查期間,各季節(jié)浮游植物的密度存在差異,為134.20萬(wàn)~283.16萬(wàn)個(gè)/L(見(jiàn)圖1)。秋季浮游植物密度最大,此時(shí)藍(lán)藻最多,占總密度的60.5%,尖頭藻和念珠藻為當(dāng)時(shí)優(yōu)勢(shì)類(lèi)群;冬季浮游植物密度最小,硅藻和綠藻為主要類(lèi)群,分別占總密度的43.1%、37.2%;春季主要浮游植物群落仍是硅藻和綠藻;夏季綠藻密度最大,占64.5%。浮游植物生物量為2.48~8.09 mg/L(見(jiàn)圖2),冬季最低,夏季最高,秋季次之,與其密度的變化情況基本一致。
結(jié)合多樣性指數(shù)和藻類(lèi)生物學(xué)評(píng)價(jià)標(biāo)準(zhǔn)評(píng)價(jià)青龍湖水質(zhì),結(jié)果見(jiàn)表4。H′值顯示青龍湖四季均處于中營(yíng)養(yǎng)狀態(tài),由于此次調(diào)查中浮游植物僅鑒定到屬,因此H′可能相對(duì)偏低。J值顯示不同季節(jié)青龍湖均處于貧營(yíng)養(yǎng)狀態(tài)。H′值與J值的評(píng)價(jià)結(jié)果與馬劍敏等[13]對(duì)該濕地的研究結(jié)果一致。生物量顯示春、冬兩季水體處于貧中營(yíng)養(yǎng)狀態(tài),夏、秋兩季處于富營(yíng)養(yǎng)狀態(tài)。浮游植物密度評(píng)價(jià)結(jié)果顯示,湖泊全年均為中營(yíng)養(yǎng)。綜合各評(píng)價(jià)指標(biāo),青龍湖春季、冬季處于貧中營(yíng)養(yǎng)-中營(yíng)養(yǎng)狀態(tài),夏季、秋季為中營(yíng)養(yǎng)-富營(yíng)養(yǎng)狀態(tài)。
2.3 環(huán)境因子
該濕地部分水質(zhì)因子調(diào)查結(jié)果見(jiàn)表5。水體pH值四季變化較小,基本保持穩(wěn)定。春、冬季湖水透明度高于夏、秋季的,原因是除季節(jié)性降水外,黃河水是青龍湖主要補(bǔ)給水源,補(bǔ)水時(shí)間、補(bǔ)給水量的不確定性導(dǎo)致水體透明度波動(dòng)較大。DO為8.81~10.53 mg/L,均符合Ⅰ類(lèi)地表水質(zhì)量標(biāo)準(zhǔn)。除秋季外,TP質(zhì)量濃度均滿(mǎn)足Ⅲ類(lèi)水標(biāo)準(zhǔn),秋季為Ⅳ類(lèi)水。NH+4-N、CODMn質(zhì)量濃度均符合Ⅱ類(lèi)水標(biāo)準(zhǔn),其中CODMn變化情況與透明度類(lèi)似,可能也受黃河水補(bǔ)給影響。相較于新鄉(xiāng)黃河濕地鳥(niǎo)類(lèi)國(guó)家級(jí)自然保護(hù)區(qū),毗鄰該研究區(qū)的另一常年集水區(qū)陳橋東湖[13],青龍湖的TP、NH+4-N質(zhì)量濃度均較低。此外,與1992年黎道豐等[14]對(duì)青龍湖的調(diào)查結(jié)果相比較,TP質(zhì)量濃度下降了約50%,NH+4-N質(zhì)量濃度增加了約76%。這種變化產(chǎn)生的原因可能是,20世紀(jì)90年代初濕地保護(hù)政策相對(duì)缺乏,百姓日常生活(如洗碗、洗衣等)產(chǎn)生的廢水排入湖體,導(dǎo)致TP質(zhì)量濃度較高;隨著青龍湖漁業(yè)養(yǎng)殖的發(fā)展,管理部門(mén)逐漸限制或禁止生活廢水入湖,以保護(hù)魚(yú)類(lèi)生長(zhǎng)及水生態(tài)環(huán)境,但這也造成了水生動(dòng)物排泄物增加,使得水體氨氮質(zhì)量濃度升高。
2.4 環(huán)境因子與多樣性指數(shù)的Pearson相關(guān)性分析
青龍湖浮游植物的Shannon-Wiener指數(shù)H′和Pielou均勻度指數(shù)J變化趨勢(shì)見(jiàn)圖3。Shannon-Wiener指數(shù)為2.490~2.896,呈現(xiàn)出從春到冬逐漸降低的趨勢(shì);Pielou均勻度指數(shù)為0.643~0.717,表現(xiàn)為春>冬>夏>秋。環(huán)境因子與物種多樣性的Pearson相關(guān)性分析結(jié)果見(jiàn)表6。結(jié)果顯示,Pielou均勻度指數(shù)與COND、TDS及Shannon-Wiener指數(shù)均呈顯著正相關(guān)關(guān)系,而Shannon-Wiener指數(shù)僅與均勻度指數(shù)顯著正相關(guān),說(shuō)明浮游植物的密度與COND、TDS等密切相關(guān)。水質(zhì)指標(biāo)間,高錳酸鹽指數(shù)、氨氮均與透明度顯著負(fù)相關(guān),從側(cè)面反映了黃河水補(bǔ)給是導(dǎo)致夏、秋季湖水CODMn和NH+4-N質(zhì)量濃度偏高的重要原因。
2.5 浮游植物與水質(zhì)因子的RDA分析
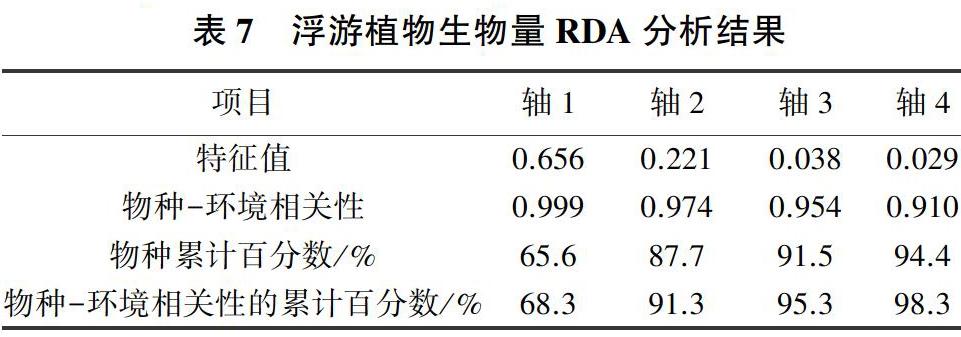
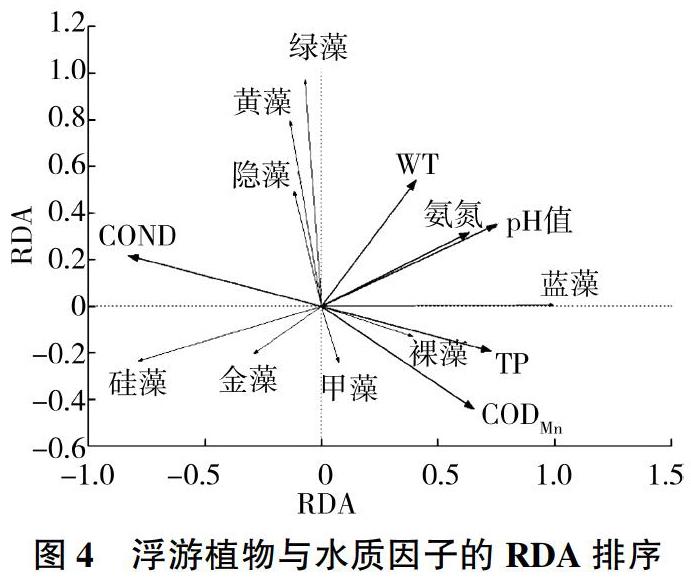
對(duì)青龍湖浮游植物生物量進(jìn)行去趨勢(shì)對(duì)應(yīng)分析(DCA),結(jié)果顯示梯度長(zhǎng)度最大為2.008,故應(yīng)對(duì)浮游植物和環(huán)境因子進(jìn)行冗余分析。對(duì)環(huán)境變量進(jìn)行自動(dòng)篩選和蒙特卡洛檢驗(yàn),共選出COND、WT、pH值、TP、NH+4-N和CODMn等6項(xiàng)膨脹因子均小于20的變量進(jìn)行RDA分析,結(jié)果見(jiàn)表7(所有典范特征值之和為0.960)。RDA排序前兩個(gè)物種軸之間的相關(guān)系數(shù)為-0.009,幾乎互相垂直。前兩個(gè)環(huán)境軸間的相關(guān)系數(shù)為0.000,相互垂直。環(huán)境因子軸與物種軸1、物種軸2的相關(guān)系數(shù)分別為0.999、0.974。第1、2排序軸的特征值分別為0.656、0.221,共解釋了87.7%的物種信息與91.3%的物種-環(huán)境關(guān)系信息,表明排序軸能較好地反映物種與水質(zhì)指標(biāo)間的關(guān)系。從圖4可以看出,硅藻、金藻與pH值、NH+4-N高度負(fù)相關(guān);藍(lán)藻、裸藻與TP正相關(guān),與COND負(fù)相關(guān);綠藻、隱藻、黃藻均與水溫正相關(guān),與CODMn負(fù)相關(guān);此外CODMn、TP與甲藻正相關(guān)。
3 討 論
浮游植物群落結(jié)構(gòu)的變化是其與外界環(huán)境相適應(yīng)的結(jié)果,受溫度、營(yíng)養(yǎng)鹽等環(huán)境因子的驅(qū)動(dòng),浮游植物的密度和生物量處于動(dòng)態(tài)變化之中[15]。浮游植物群落存在明顯的季節(jié)演替現(xiàn)象[16-18]。研究發(fā)現(xiàn),春季青龍湖浮游植物群落結(jié)構(gòu)為硅藻-綠藻型,夏季轉(zhuǎn)以綠藻為主,秋季以藍(lán)藻為主,冬季則以硅藻、綠藻為主。浮游植物的密度和生物量大體表現(xiàn)為夏、秋>春、冬。一般認(rèn)為水體中總氮含量>0.2 mg/L、總磷含量>0.02 mg/L時(shí),水華爆發(fā)的概率較大[19]。夏、秋時(shí)溫度、光強(qiáng)較高,營(yíng)養(yǎng)鹽充足,滿(mǎn)足綠藻、藍(lán)藻的適宜生長(zhǎng)條件,故此時(shí)念珠藻、尖頭藻等藻類(lèi)大量繁殖,浮游植物的總密度和生物量較大。硅藻和綠藻有很多普適種和冷水種,對(duì)環(huán)境適應(yīng)性較好,容易形成優(yōu)勢(shì)種;藍(lán)藻在水溫較低時(shí)會(huì)下沉到底部休眠[20]。研究表明,青龍湖中浮游植物的演替規(guī)律與Sommer等[21]提出的浮游植物生態(tài)群落(PEG)演替模式基本一致。
物種多樣性指數(shù)反映了群落特有的物種組成和密度特征,可以用來(lái)表征群落結(jié)構(gòu)的穩(wěn)定性以及種群分布的均勻程度[22-24]。H′值和J值分別大于3.0、0.8時(shí),表明浮游植物群落結(jié)構(gòu)較為穩(wěn)定。研究區(qū)Shannon-Wiener指數(shù)和Pielou均勻度指數(shù)均小于上述值,且呈現(xiàn)基本一致的變化趨勢(shì)。夏、秋季浮游植物密度較高時(shí),Shannon-Wiener指數(shù)和Pielou指數(shù)較小,說(shuō)明此時(shí)某些優(yōu)勢(shì)藻類(lèi)的密度較大,導(dǎo)致生物多樣性指數(shù)減小。物種多樣性指數(shù)和環(huán)境因子的Pearson相關(guān)性分析表明,Shannon-Wiener指數(shù)、COND、TDS與均勻度指數(shù)高度正相關(guān);同時(shí),RDA分析結(jié)果表明,COND與藍(lán)藻等富營(yíng)養(yǎng)狀態(tài)指示物種負(fù)相關(guān)。TDS一般用來(lái)衡量水體中所有離子的總含量;COND反映了水的純度,水體中離子態(tài)的物質(zhì)越多,COND越大。水華爆發(fā)期間需要消耗水體中大量氮、磷等營(yíng)養(yǎng)鹽,水體物質(zhì)含量的減少使得電導(dǎo)率下降[25]。這與本文RDA分析結(jié)果一致,與Pearson相關(guān)性分析結(jié)果相左。其原因可能是,Pielou指數(shù)是根據(jù)各藻類(lèi)優(yōu)勢(shì)屬密度計(jì)算得出的,反映了種群分布的均勻程度,包括但不限于藍(lán)藻等易發(fā)生水華的物種。
藻類(lèi)生物學(xué)評(píng)價(jià)結(jié)果表明,冬季青龍湖處于貧中-中營(yíng)養(yǎng)狀態(tài),夏、秋季為中-富營(yíng)養(yǎng)狀態(tài)。結(jié)合水體理化性質(zhì)來(lái)看,夏季水質(zhì)全年最差。其原因可能是,夏季溫度較高,湖水蒸發(fā)量大且水位低,外界的擾動(dòng)易造成底泥的再懸浮,使水體污染物增多;夏季降雨較多,該濕地周邊道路全年施工不斷且毗鄰農(nóng)田,地表徑流夾雜著污染物輸入水體。
浮游植物能快速響應(yīng)環(huán)境因素的變化,其群落特征是多種環(huán)境因素在時(shí)間和空間上綜合作用的結(jié)果[26]。不同水體,影響浮游植物的主要環(huán)境因子不盡相同[27-29]。RDA分析表明,青龍湖中浮游植物群落主要與水溫、營(yíng)養(yǎng)鹽、pH值、CODMn和電導(dǎo)率等因素緊密相關(guān)。溫度是各種生物生存的必要條件,水溫主要通過(guò)影響藻類(lèi)細(xì)胞酶的活性以及水體的對(duì)流運(yùn)動(dòng)影響浮游植物的生命活動(dòng)[30]。不同種類(lèi)的浮游植物都有其適宜生長(zhǎng)的溫度,如綠藻、藍(lán)藻等藻類(lèi)嗜高溫,硅藻、金藻等好低溫[31]。金藻作為清潔水質(zhì)的指示種,在冬季浮游植物密度和生物量比其他季節(jié)的高,表明冬季青龍湖水質(zhì)較好。本研究結(jié)果表明,溫度是青龍湖浮游植物群落季節(jié)性演替的重要因素。
氮、磷等營(yíng)養(yǎng)物質(zhì)是浮游植物生長(zhǎng)繁殖的物質(zhì)基礎(chǔ),國(guó)際上一般認(rèn)為水體中總氮含量大于0.2 mg/L,總磷含量大于0.02 mg/L時(shí),水體富營(yíng)養(yǎng)化。淡水湖泊中磷通常被認(rèn)為是藍(lán)藻水華的主要限制元素[32]。Abell等[33]認(rèn)為,氮磷含量比小于7時(shí),氮為藻類(lèi)生長(zhǎng)的主要限制性營(yíng)養(yǎng)元素。青龍湖NH+4-N與TP的含量分別大于0.2、0.02 mg/L,且兩者含量的比值秋季時(shí)約為6,其他季節(jié)均大于7。考慮未檢測(cè)總氮含量,實(shí)際氮磷比值更大。因此,磷元素可能是青龍湖浮游植物生長(zhǎng)的主要限制因子。此外,Downing等[34]研究發(fā)現(xiàn),磷濃度小于0.03 mg/L時(shí),藍(lán)藻形成優(yōu)勢(shì)的風(fēng)險(xiǎn)小于10%;當(dāng)磷濃度為0.03~0.07 mg/L時(shí),風(fēng)險(xiǎn)增至40%;當(dāng)總磷含量接近0.1 mg/L時(shí),風(fēng)險(xiǎn)高達(dá)80%。青龍湖不同季節(jié)總磷含量均大于0.03 mg/L,因此藍(lán)藻可能有大量繁殖的風(fēng)險(xiǎn)。
除溫度和營(yíng)養(yǎng)鹽外,pH值也是影響浮游植物生長(zhǎng)和分布的重要因素,pH值的變化對(duì)細(xì)胞酶活性有一定影響[35]。和溫度相似,不同種類(lèi)的浮游植物都有適宜其生命活動(dòng)的pH值范圍。相關(guān)研究表明,堿性水環(huán)境較適宜浮游植物進(jìn)行光合作用[36]。本研究中,pH值與第一、第二物種軸均正相關(guān),說(shuō)明pH值與藻類(lèi)生產(chǎn)力密切相關(guān)。
CODMn反映了水體有機(jī)物的含量,青龍湖中污染物主要源于漁業(yè)養(yǎng)殖、生活污水和部分農(nóng)田廢水排放。本研究表明,CODMn也是影響浮游植物生物量的重要因素,這與對(duì)漢豐湖[29]等的調(diào)查結(jié)果一致。
4 結(jié) 論
對(duì)青龍湖共鑒定出8門(mén)53屬浮游植物,綠藻最多有23屬,其次為硅藻14屬;浮游植物密度為134.20萬(wàn)~283.16萬(wàn)個(gè)/L,生物量為2.48~8.09 mg/L;季節(jié)變化方面,藻類(lèi)的生物量夏、秋高于春、冬,浮游植物的密度則是秋季最高,春、夏次之;浮游植物的演替模式與PEG模式相符。基于藻類(lèi)生物學(xué)評(píng)價(jià)標(biāo)準(zhǔn)得出春季、冬季青龍湖處于貧中-中營(yíng)養(yǎng)狀態(tài),夏、秋兩季為中-富營(yíng)養(yǎng)狀態(tài)。冗余分析(RDA)結(jié)果表明,水溫、pH值、總磷、氨氮、CODMn和電導(dǎo)率是影響青龍湖浮游植物群落的主要環(huán)境因素。
參考文獻(xiàn):
[1] FIELD C B, BEHRENFELD M J, RANDERSON J T, et al. Primary Production of the Biosphere: Integrating Terrestrial and Oceanic Components[J]. Science, 1998, 281: 237-240.
[2] FEIO M J, ALMEIDA S F P, CRAVEIRO S C, et al. A Comparison Between Biotic Indicesand Predictive Models in Stream Water Quality Assessment Based on Benthic Diatom Communities[J]. Ecological Indicators, 2008, 9(3): 497-507.
[3] PADISAK J, BORICS G, GRIGORSZKY I, et al. Use of Phytoplankton Assemblages for Monitoring Ecological Status of Lakes Within the Water Framework Directive: the Assemblage Index[J]. Hydrobiologia, 2006, 553(1): 1-14.
[4] 吉正元,劉紹俊.撫仙湖浮游植物群落結(jié)構(gòu)、影響因子及水質(zhì)評(píng)價(jià)[J].中國(guó)環(huán)境監(jiān)測(cè),2019,35(4):67-77.
[5] 姜慶宏,王佳寧,李衛(wèi)平,等.南海湖春季浮游植物群落結(jié)構(gòu)及其與水質(zhì)因子的關(guān)系[J].干旱區(qū)資源與環(huán)境,2018,32(8): 166-171.
[6] 劉輝,鄒繼穎,邊紅楓,等.吉林市小型景觀(guān)水體夏季浮游植物群落特征及水質(zhì)評(píng)價(jià)[J].海洋湖沼通報(bào),2018(4): 91-99.
[7] 蔡慶華,伍焯田,黎道豐,等.河南曹崗湖浮游動(dòng)物野外現(xiàn)場(chǎng)試驗(yàn)初報(bào)[J].水生生物學(xué)報(bào),1995,19(2):190-192.
[8] 胡鴻鈞,魏印心.中國(guó)淡水藻類(lèi):系統(tǒng)、分類(lèi)及生態(tài)[M].北京:科學(xué)出版社,2006:57-333.
[9] 國(guó)家環(huán)境保護(hù)總局,水和廢水監(jiān)測(cè)分析方法編委會(huì).水和廢水監(jiān)測(cè)分析方法[M].4版.北京:中國(guó)環(huán)境科學(xué)出版社,2002:223-279.
[10] 張覺(jué)民,何志輝.內(nèi)陸水域漁業(yè)自然資源調(diào)查手冊(cè)[M].北京:農(nóng)業(yè)出版社,1991:20-23.
[11] 況琪軍,馬沛明,胡征宇,等.湖泊富營(yíng)養(yǎng)化的藻類(lèi)生物學(xué)評(píng)價(jià)與治理研究進(jìn)展[J].安全與環(huán)境學(xué)報(bào),2005,5(2):87-91.
[12] 孫軍,劉東艷.多樣性指數(shù)在海洋浮游植物研究中的應(yīng)用[J].海洋學(xué)報(bào)(中文版),2004,26(1):62-75.
[13] 馬劍敏,褚一凡,趙閃閃,等.陳橋東湖浮游植物群落結(jié)構(gòu)與環(huán)境因子的關(guān)系[J].河南師范大學(xué)學(xué)報(bào)(自然科學(xué)版),2019,47(2):83-91.
[14] 黎道豐,劉瑞秋,倪樂(lè)意,等.黃淮海平原封丘試區(qū)水體理化性狀的多元分析[J].水生生物學(xué)報(bào),1992,16(3):251-259.
[15] XIAO L J,WANG T. Grazing of Daphnia Galeata and Phyllodiaptomus Tunguidus on Phytoplankton in Liuxihe Reservoir, South China: Insitu Bottle Experiments[J]. Ecological Science, 2008, 27(5): 362-367.
[16] LU J, YANG Z, ZHANG Y. Algae Functional Group Characteristics Inreservoirs and Lakes with Different Trophic Levels in Northwesternsemi-Humid and Semi-Arid Regions in China[J]. Journal of Environmental Sciences, 2018, 64(2): 166-173.
[17] 賀玉曉,劉天慧,任玉芬,等.北運(yùn)河秋冬季浮游植物群落結(jié)構(gòu)特征及影響因子分析[J].環(huán)境科學(xué)學(xué)報(bào),2020,40(5): 1710-1721.
[18] 汪琪,黃蔚,陳開(kāi)寧,等.大溪水庫(kù)浮游植物群落結(jié)構(gòu)特征及營(yíng)養(yǎng)狀態(tài)評(píng)價(jià)[J].環(huán)境科學(xué)學(xué)報(bào),2020,40(4):1286-1297.
[19] 劉光釗.水體富營(yíng)養(yǎng)及其藻害[M].北京:中國(guó)環(huán)境科學(xué)出版社,2005:216-272.
[20] 姜慶宏,潘彤,李衛(wèi)平,等.南海湖冬季浮游植物特征及與水質(zhì)因子關(guān)系[J].東北林業(yè)大學(xué)學(xué)報(bào),2019,47(4):71-75.
[21] SOMMER U, GLIWICZ Z M, LAMPERT W, et al. The PEG-Model of Seasonal Succession of Planktonic Events in Fresh Waters[J]. Arch. Hydrobiol, 1986, 106(4): 433-471.
[22] 沈會(huì)濤,劉存歧.白洋淀浮游植物群落及其與環(huán)境因子的典范對(duì)應(yīng)分析[J].湖泊科學(xué),2008,20(6):773-779.
[23] 孟順龍,瞿建宏,裘麗萍,等. 富營(yíng)養(yǎng)化水體降磷對(duì)浮游植物群落結(jié)構(gòu)特征的影響[J]. 生態(tài)環(huán)境學(xué)報(bào),2013,22(9):1578-1582.
[24] 陳琳荔,鄒華,莊嚴(yán),等.小溪港浮游植物群落特征及水質(zhì)評(píng)價(jià)[J].環(huán)境科學(xué)研究,2014,27(9):1016-1023.
[25] 張慶文,宋林旭,紀(jì)道斌,等.香溪河庫(kù)灣水質(zhì)特征與非回水區(qū)水華響應(yīng)關(guān)系[J].中國(guó)環(huán)境科學(xué),2019,39(7):3018-3026.
[26] LAMPE R H, WANG S, CASSAR N, et al. Strategies Among Phytoplankton in Response to Alleviation of Nutrient Stress in a Subtropical Gyre[J]. ISME Journal,2019,13(12):2984-2997.
[27] 徐宗學(xué),劉麟菲.基于著生藻類(lèi)的渭河流域水生態(tài)系統(tǒng)健康評(píng)價(jià)[J].人民黃河,2020,42(9):123-129.
[28] 劉黎,賀新宇,付君珂,等.三峽水庫(kù)干流底棲硅藻群落組成及其與環(huán)境因子的關(guān)系[J].環(huán)境科學(xué),2019,40(8):3577-3587.
[29] 胡芯,印江平,唐洪玉,等.烏楊調(diào)節(jié)壩運(yùn)行初期漢豐湖浮游植物群落結(jié)構(gòu)及其與環(huán)境因子的相關(guān)性分析[J].淡水漁業(yè),2019,49(6):48-55.
[30] 周彥鋒,宋江騰,劉凱,等.懷洪新河浮游植物群落結(jié)構(gòu)與水環(huán)境因子的關(guān)系研究[J].生態(tài)科學(xué),2017,36(1): 35-42.
[31] 李博韜,劉凌,朱燕,等.利用浮游植物生物完整性對(duì)河流健康進(jìn)行評(píng)價(jià)[J].人民黃河,2020,42(6):73-78.
[32] LIN Y J, HE Z L, YANG Y G, et al. Nitrogen Versus Phosphorus Limitation of Phytoplankton Growth in Ten Mile Creek, Florida, USA[J]. Hydrobiologia, 2008, 605(1): 247-258.
[33] ABELL J, ZKUNDAKCI D, HAMILTON D. Nitrogen and Phosphorus Limitationof Phytoplankton Growth in New Zealand Lakes: Implications for Eutrophication Control[J]. Ecosystems, 2010, 13(7): 966-977.
[34] DOWNING J A, WATSON S B, MCCAULEY E. Predicting Cyanobacteria Dominance in Lakes[J]. Canadian Journal of Fisheries and Aquatic Sciences, 2001, 58(10): 1905-1908.
[35] 王茂偉,周利,高靜思,等.水體pH對(duì)偽魚(yú)腥藻生長(zhǎng)及葉綠素?zé)晒鈪?shù)的影響[J].生態(tài)科學(xué),2019,38(3):159-165.
[36] JAKOBSEN H H, BLANDA E, STAEHR P A, et al. Development of Phytoplankton Communities: Implications of Nutrient Injections on Phytoplankton Composition, pH and Ecosystem Production[J]. Journal of Experimental Marine Biology and Ecology, 2015, 473: 81-89.
【責(zé)任編輯 呂艷梅】
